Targeting the NLRP3 Inflammasome-Related Pathways via Tianeptine Treatment-Suppressed Microglia Polarization to the M1 Phenotype in Lipopolysaccharide-Stimulated Cultures

 , , and
, , and
Abstract
:1. Introduction
2. Results
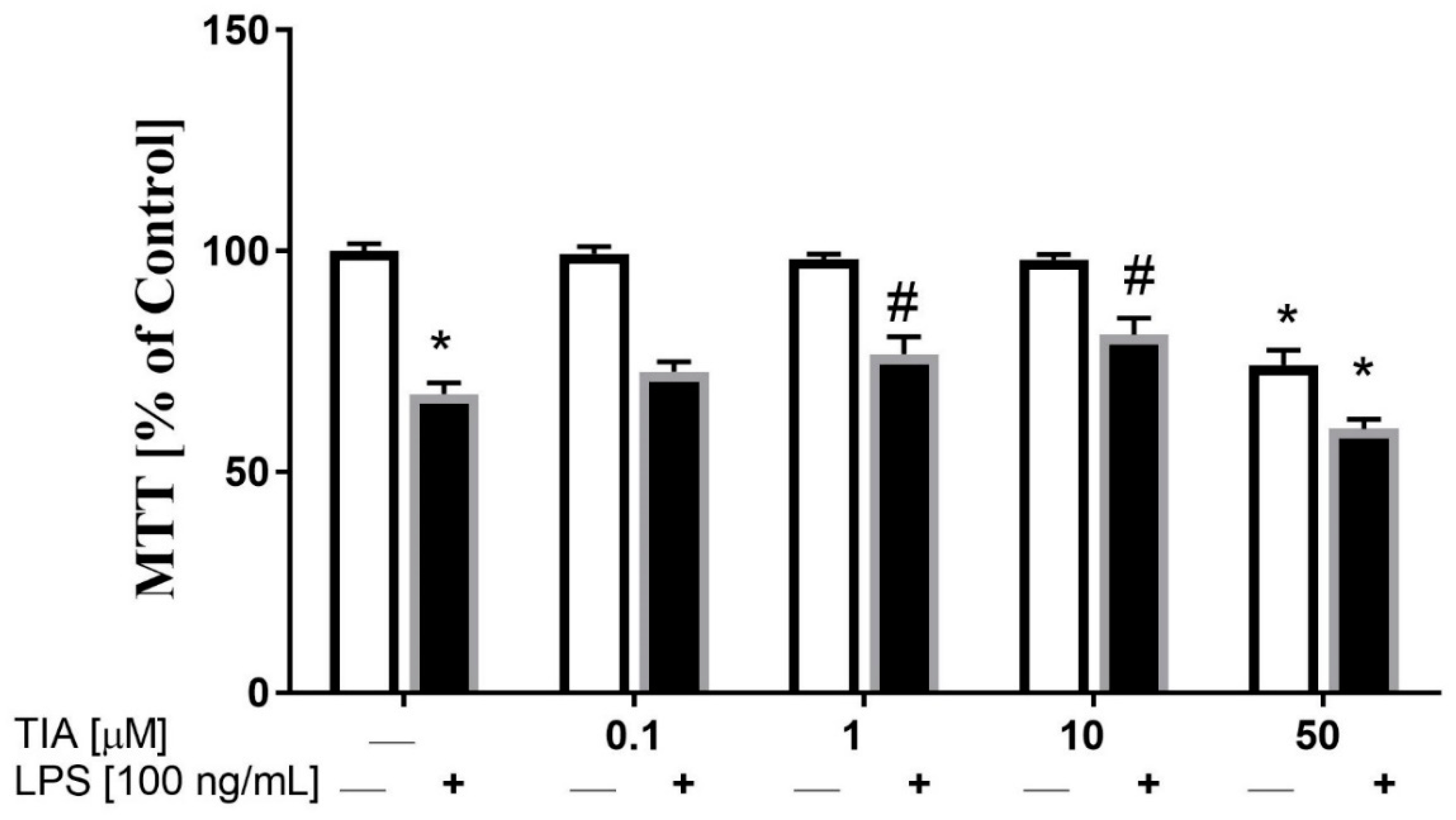
2.1. Tianeptine Modulates Changes in the Viability of LPS-Stimulated Microglial Cells
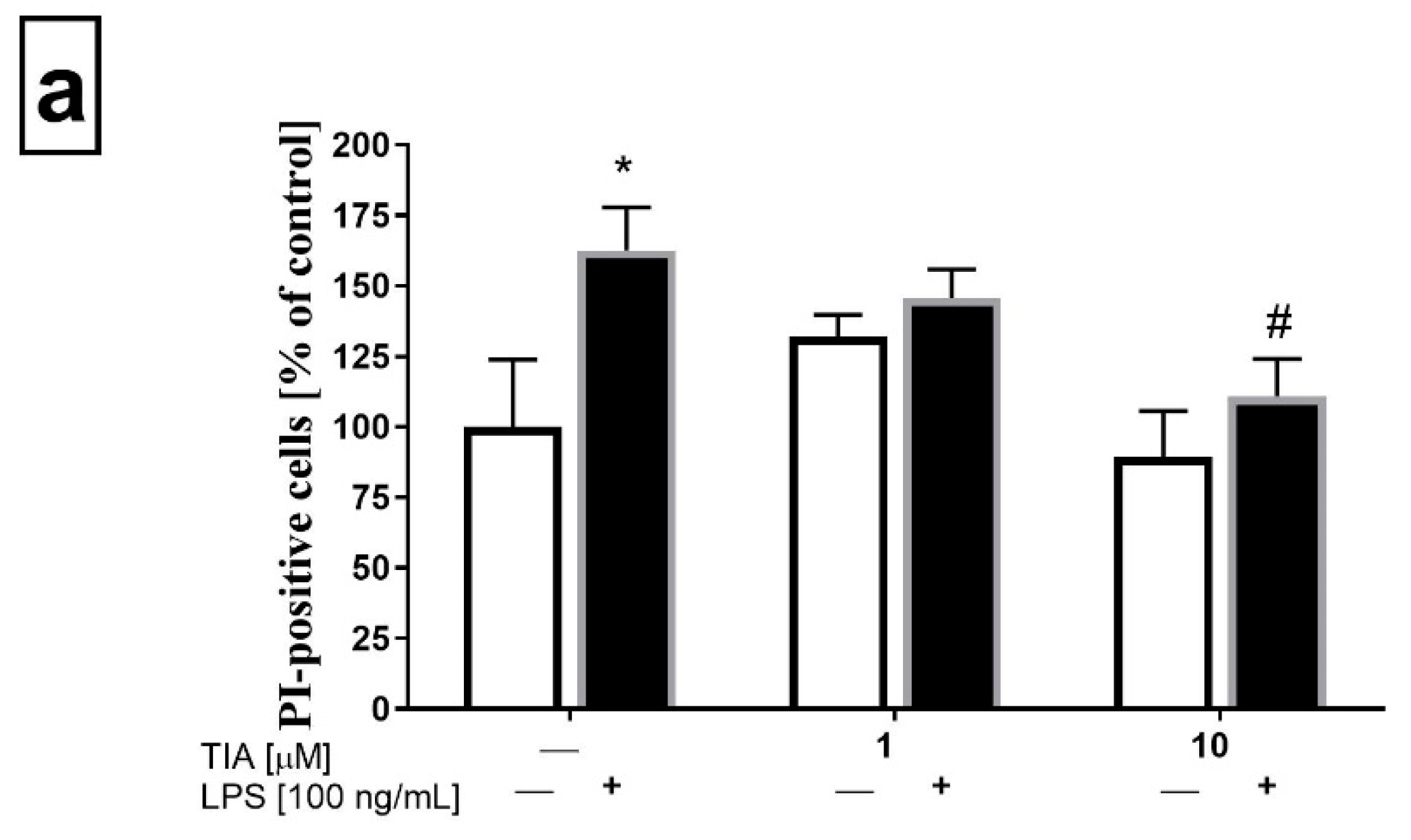
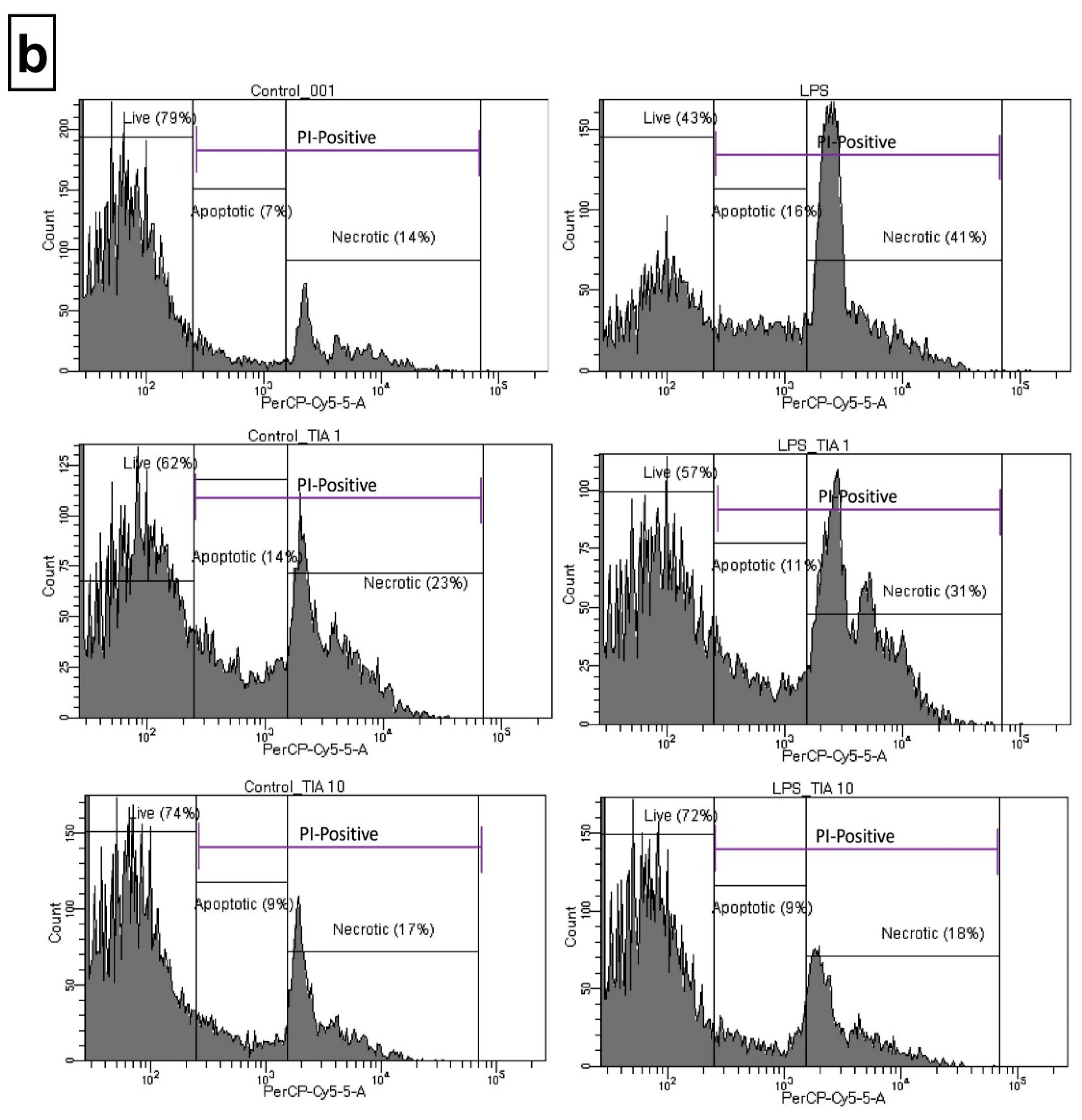
2.2. Tianeptine Modulates Changes in the Number of Propidium Iodide (PI)-Positive LPS-Stimulated Microglia Cells
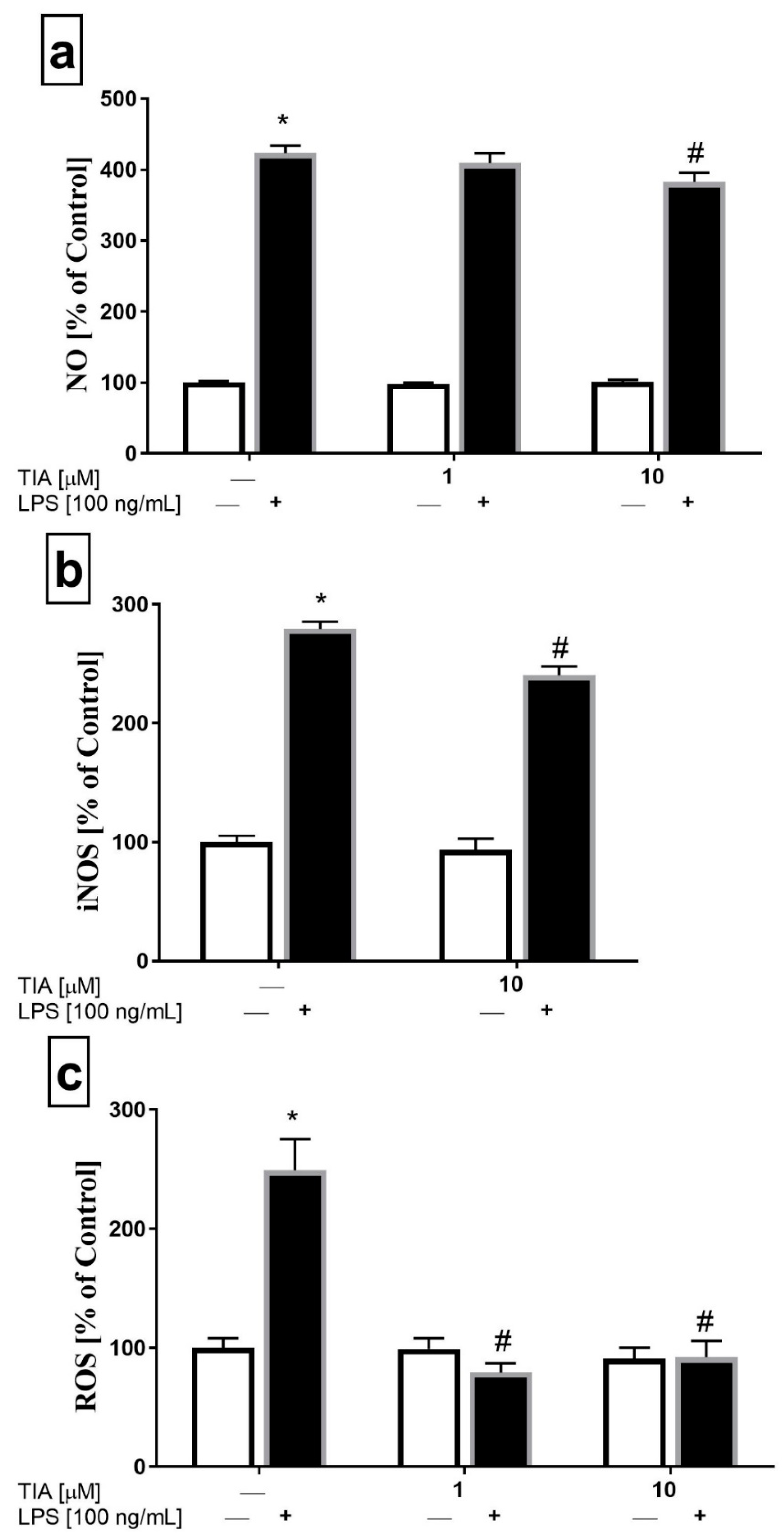
2.3. Tianeptine Diminishes NO Production, As Well As iNOS and ROS Levels in LPS-Stimulated Microglial Cells
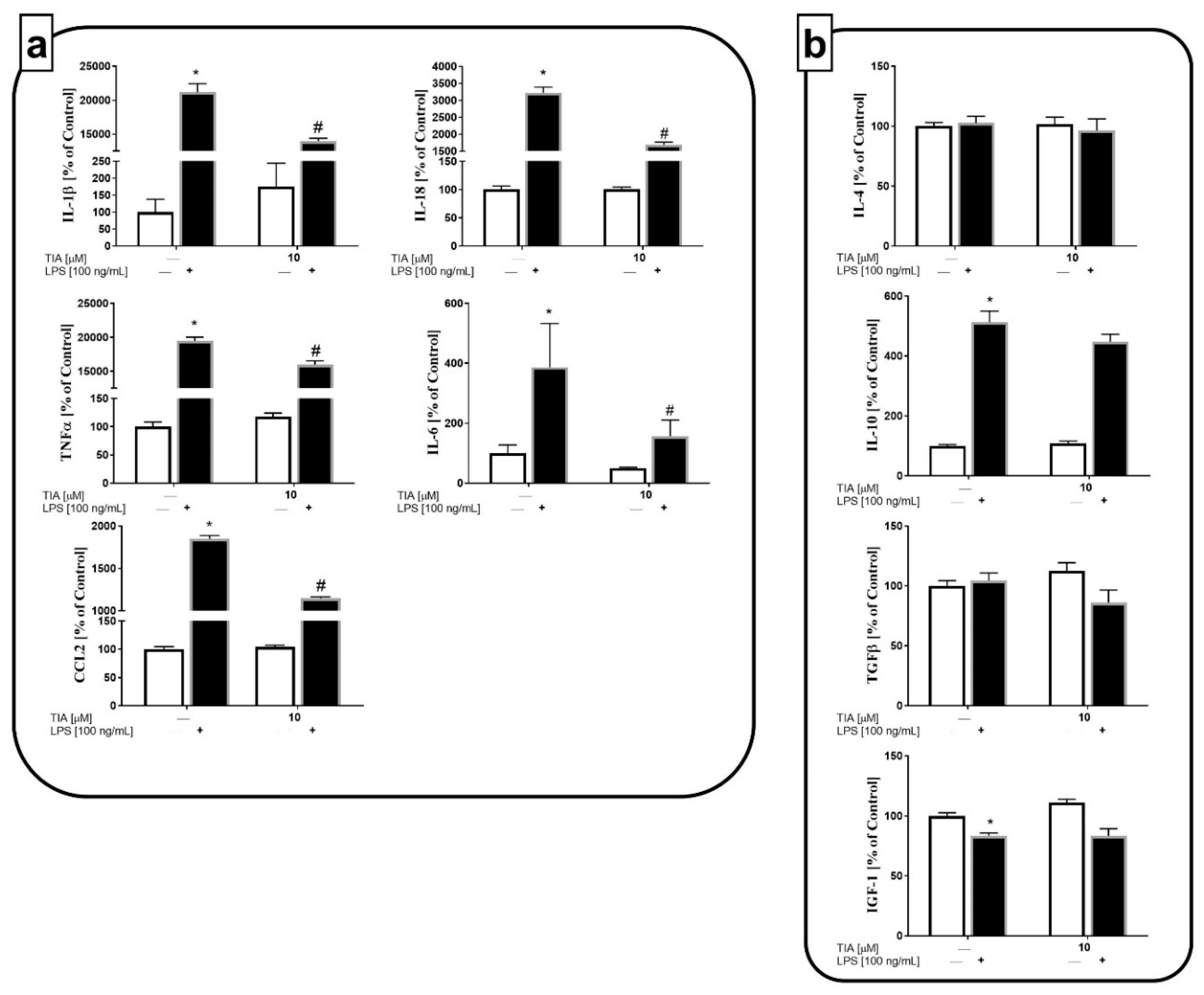
2.4. Tianeptine Reduces the Expression of Microglial Activation Markers in LPS-Stimulated Cells
2.5. Tianeptine Suppresses Microglia Polarization to the M1 but Not the M2 Phenotype in LPS-Stimulated Cells
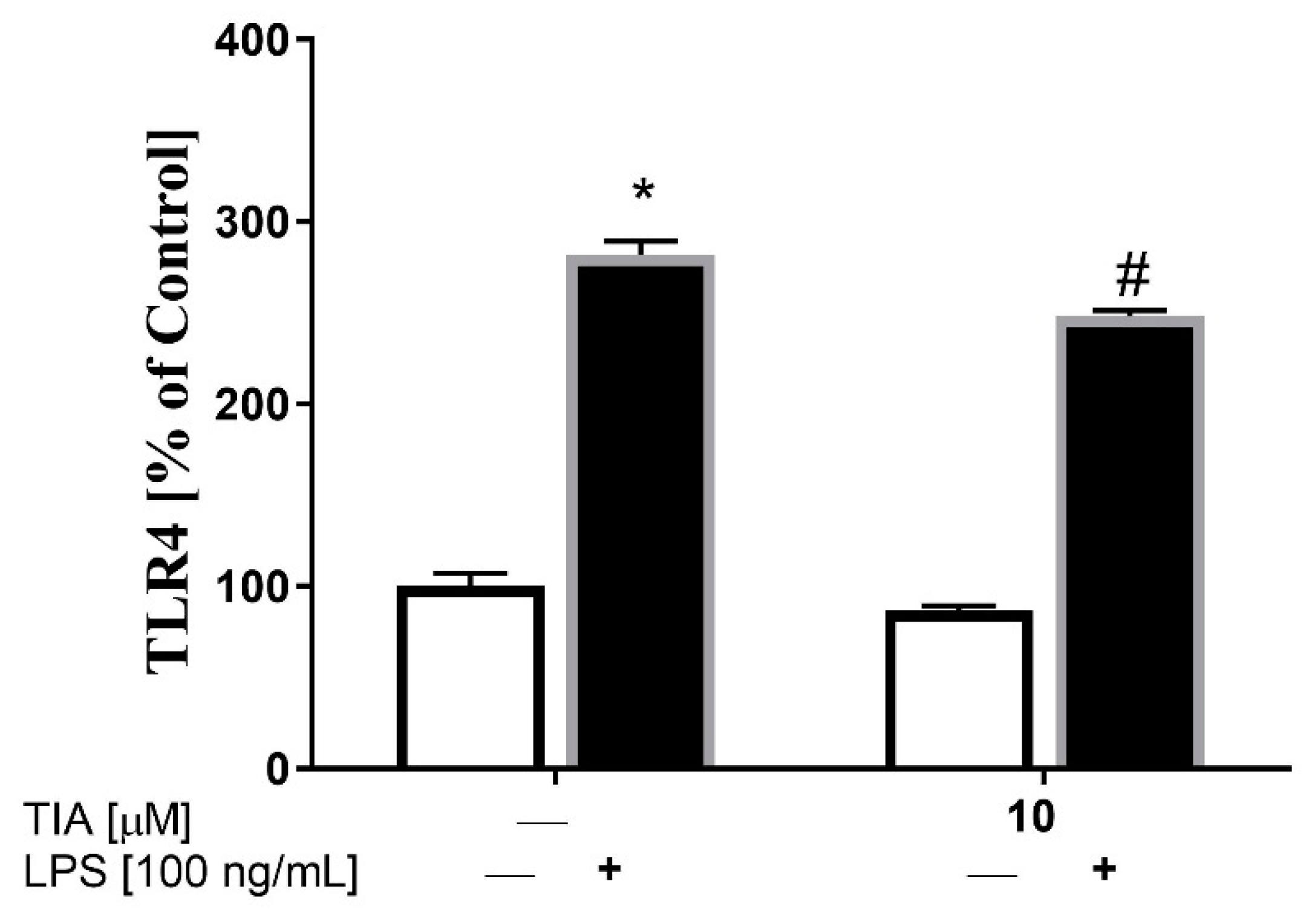
2.6. Tianeptine Reduces the Level of TLR4 in LPS-Activated Microglia Cells
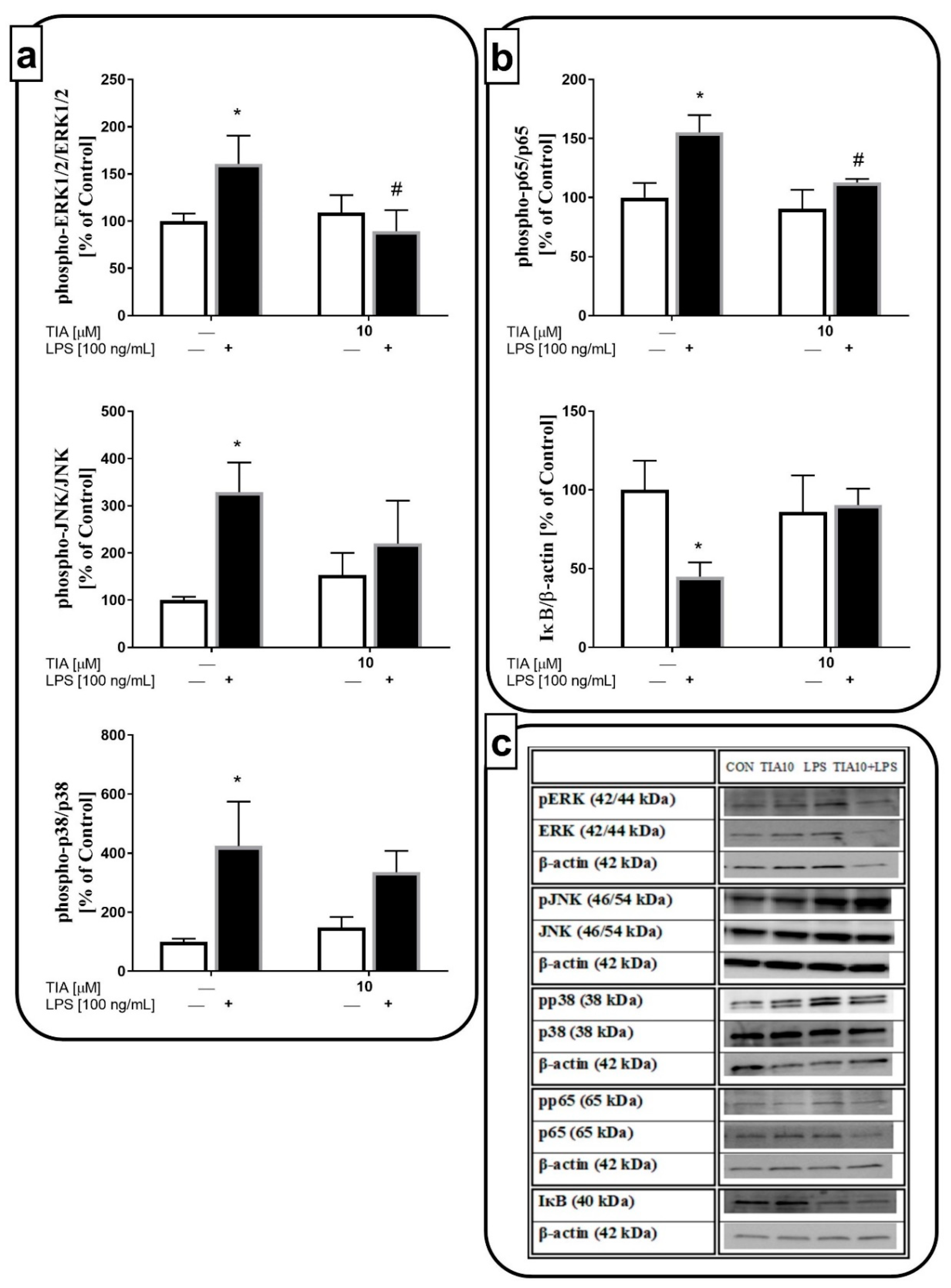
2.7. Tianeptine Inhibits LPS-Induced Microglial Activation through the ERK1/2 and NF-κB Pathways
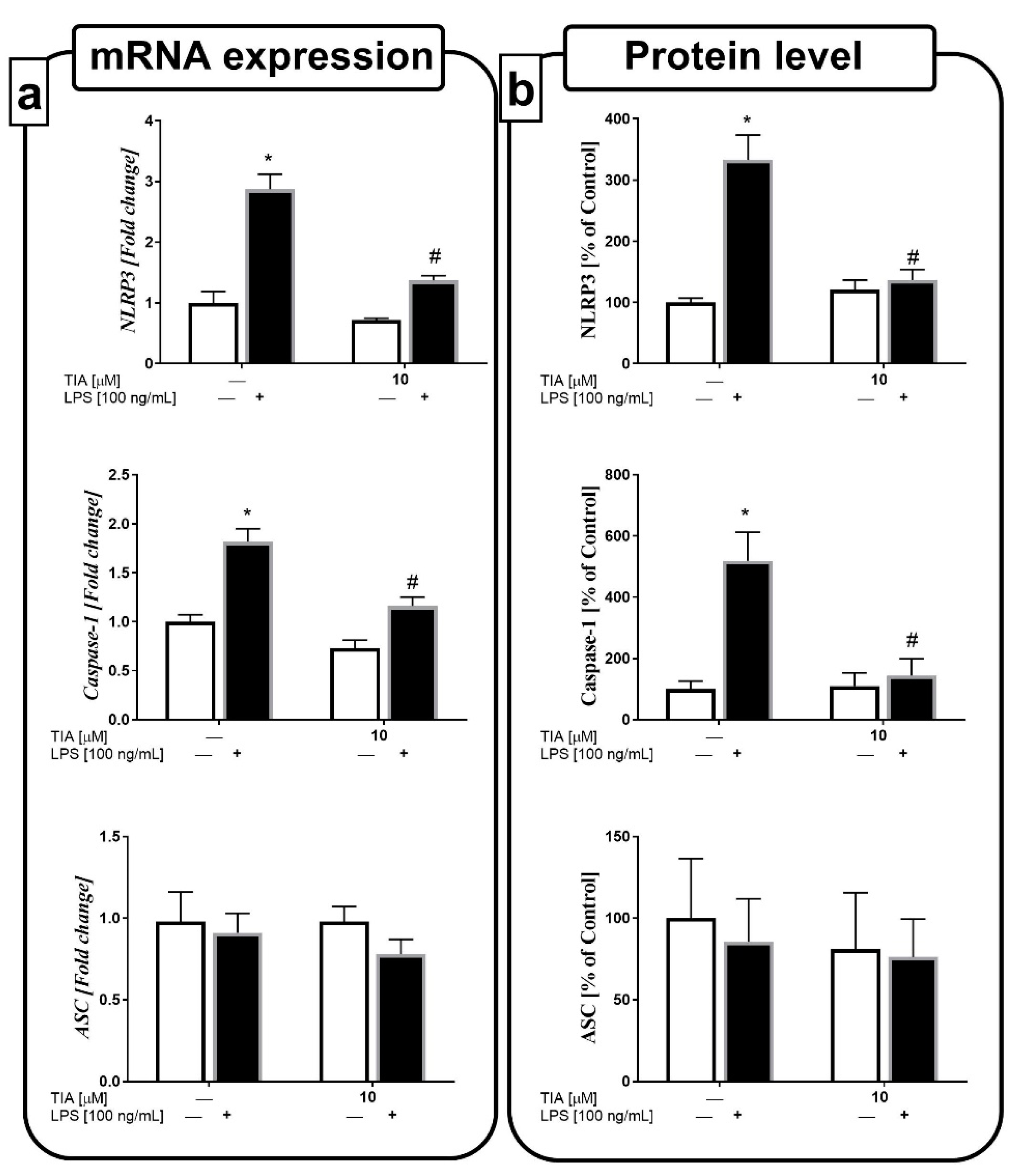
2.8. Tianeptine Diminishes NLRP3 Inflammasome Activation in LPS-Stimulated Microglial Cells
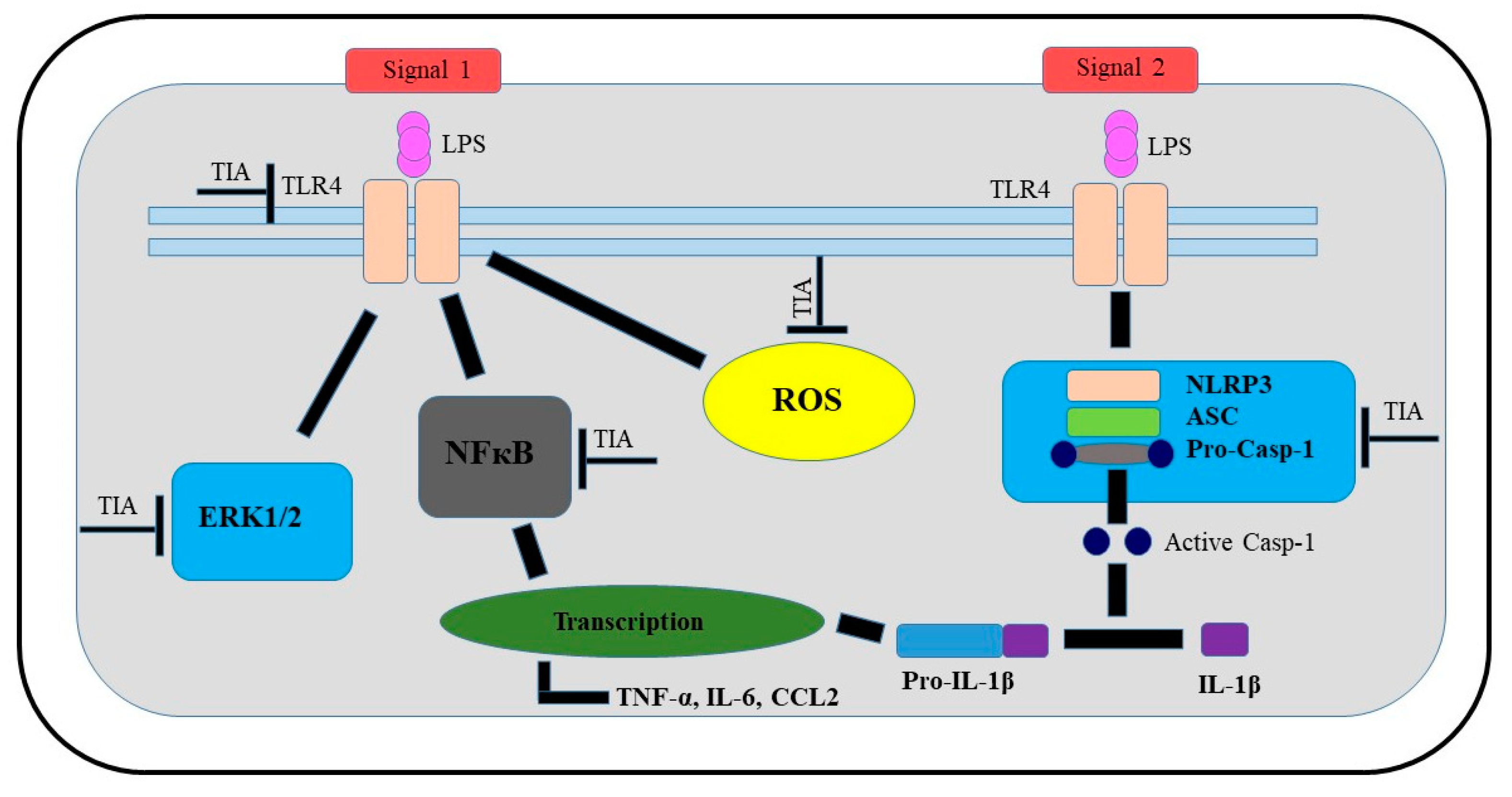
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Culture
4.3. Cell Treatment
4.4. Cell Viability Test
4.5. Flow Cytometry Analysis
4.6. Nitric Oxide Release Assay
4.7. Measurement of Intracellular ROS Formation
4.8. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.9. Western Blotting Analysis
4.10. Enzyme-Linked Immunosorbent Assay (ELISA)
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AP-1 | activator protein-1 |
| ASC | apoptosis-associated speck-like protein containing a caspase recruitment domain |
| Casp-1 | caspase-1 |
| CD40 | cluster of differentiation 40 |
| CNS | central nervous system |
| CX3CL1 | fractalkine |
| CX3CR1 | fractalkine receptor |
| DCFH | 2′,7′-dichlorodihydrofluorescin |
| DCFH-DA | 2′,7′-dichlorofluorescin diacetate |
| DMEM | Dulbecco’s modified Eagle medium |
| ERK 1/2 | extracellular signal–regulated kinases |
| FBS | fetal bovine serum |
| HBSS | Hank’s balanced salt solution |
| HPA | hypothalamus-pituitary-adrenal |
| IκB | inhibitor of κB kinases |
| IGF-1 | insulin-like growth factor |
| IL | interleukin |
| iNOS | inducible nitric oxide synthase |
| JNK | c-Jun N-terminal kinases |
| CCL2/MCP-1 | chemokine CC motif ligand 2/monocyte chemoattractant protein-1 |
| MD-2 | myeloid differentiation protein-2 |
| MHC II | major histocompatibility complex class II |
| MTT | tetrazolium salt 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NLRP3 | NOD-like receptor pyrin-containing 3 inflammasome |
| NO | nitric oxide |
| PBS | phosphate-buffered saline |
| PKCδ | protein kinase C δ type |
| RIPA | radioimmunoprecipitation assay |
| ROS | reactive oxygen species |
| SSRIs | selective serotonin uptake inhibitors |
| TBS | Tris-buffered saline |
| TCAs | tricyclic antidepressants |
| TGF-β | transforming growth factor β |
| TLR | Toll-like receptor |
| TNF-α | tumor necrosis factor α |
References
- Maes, M.; Yirmyia, R.; Noraberg, J.; Brene, S.; Hibbeln, J.; Perini, G.; Kubera, M.; Bob, P.; Lerer, B.; Maj, M. The inflammatory & neurodegenerative (I&ND) hypothesis of depression: Leads for future research and new drug developments in depression. Metab. Brain Dis. 2009, 24, 27–53. [Google Scholar] [CrossRef] [PubMed]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctôt, K.L. A Meta-Analysis of Cytokines in Major Depression. Biol. Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Szczesny, E.; Basta-Kaim, A.; Slusarczyk, J.; Trojan, E.; Glombik, K.; Regulska, M.; Leskiewicz, M.; Budziszewska, B.; Kubera, M.; Lason, W. The impact of prenatal stress on insulin-like growth factor-1 and pro-inflammatory cytokine expression in the brains of adult male rats: The possible role of suppressors of cytokine signaling proteins. J. Neuroimmunol. 2014, 276, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Galecki, P.; Szemraj, J.; Florkowski, A.; Talarowska, M.; Bienkiewicz, M.; Galecka, E.; Lewinski, A. Single nucleotide polymorphism of the KIBRA gene in recurrent depressive disorders. Neuro Endocrinol. Lett. 2010, 31, 97–102. [Google Scholar] [PubMed]
- Nakagawa, Y.; Chiba, K. Role of microglial M1/M2 polarization in relapse and remission of psychiatric disorders and diseases. Pharmaceuticals 2014, 7, 1028–1048. [Google Scholar] [CrossRef] [PubMed]
- Joca, S.R.L.; Ferreira, F.R.; Guimarães, F.S. Modulation of stress consequences by hippocampal monoaminergic, glutamatergic and nitrergic neurotransmitter systems. Stress 2007, 10, 227–249. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, U.K. Microglia as a source and target of cytokines. Glia 2002, 40, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Goldmann, T.; Prinz, M. Role of Microglia in CNS Autoimmunity. Clin. Dev. Immunol. 2013, 2013, 208093. [Google Scholar] [CrossRef] [PubMed]
- Polazzi, E.; Monti, B. Microglia and neuroprotection: From in vitro studies to therapeutic applications. Prog. Neurobiol. 2010, 92, 293–315. [Google Scholar] [CrossRef] [PubMed]
- Kohman, R.A.; Rhodes, J.S. Neurogenesis, inflammation and behavior. Brain Behav. Immun. 2013, 27, 22–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frick, L.R.; Williams, K.; Pittenger, C. Microglial dysregulation in psychiatric disease. Clin. Dev. Immunol. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, M.-E.; Stevens, B.; Sierra, A.; Wake, H.; Bessis, A.; Nimmerjahn, A. The Role of Microglia in the Healthy Brain. J. Neurosci. 2011, 31, 16064–16069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryanpour, R.; Pasbakhsh, P.; Zibara, K.; Namjoo, Z.; Beigi Boroujeni, F.; Shahbeigi, S.; Kashani, I.R.; Beyer, C.; Zendehdel, A. Progesterone therapy induces an M1 to M2 switch in microglia phenotype and suppresses NLRP3 inflammasome in a cuprizone-induced demyelination mouse model. Int. Immunopharmacol. 2017, 51, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.M.; Yang, S.; Huang, S.S.; Tang, B.S.; Guo, J.F. Microglial activation in the pathogenesis of Huntington’s Disease. Front. Aging Neurosci. 2017, 9, 193. [Google Scholar] [CrossRef] [PubMed]
- Ślusarczyk, J.; Trojan, E.; Głombik, K.; Budziszewska, B.; Kubera, M.; Lasoń, W.; Popiołek-Barczyk, K.; Mika, J.; Wędzony, K.; Basta-Kaim, A. Prenatal stress is a vulnerability factor for altered morphology and biological activity of microglia cells. Front. Cell. Neurosci. 2015, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Jana, M.; Jana, A.; Liu, X.; Ghosh, S.; Pahan, K. Involvement of Phosphatidylinositol 3-Kinase-Mediated Up-Regulation of IκBα in Anti-Inflammatory Effect of Gemfibrozil in Microglia. J. Immunol. 2007, 179, 4142–4152. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.Y.; Park, J.S.; Jung, J.S.; Kim, D.H.; Kim, H.S. Anti-inflammatory effect of ginsenoside Rg5 in lipopolysaccharide-stimulated BV2 microglial cells. Int. J. Mol. Sci. 2013, 14, 9820–9833. [Google Scholar] [CrossRef] [PubMed]
- Franco, R.; Fernández-Suárez, D. Alternatively activated microglia and macrophages in the central nervous system. Prog. Neurobiol. 2015, 131, 65–86. [Google Scholar] [CrossRef] [PubMed]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial M1/M2 polarization and metabolic states. Br. J. Pharmacol. 2016, 173, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Liang, Z.; Yang, H.; Pan, Y.; Zheng, Y.; Wang, X. Tenuigenin protects dopaminergic neurons from inflammation via suppressing NLRP3 inflammasome activation in microglia. J. Neuroinflamm. 2017, 14, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kettenmann, H.; Hanisch, U.-K.; Noda, M.; Verkhratsky, A. Physiology of Microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseau, M.C.; Hsu, R.Y.C.; Spicer, J.D.; McDonald, B.; Chan, C.H.F.; Perera, R.M.; Giannias, B.; Chow, S.C.; Rousseau, S.; Law, S.; et al. Lipopolysaccharide-induced toll-like receptor 4 signaling enhances the migratory ability of human esophageal cancer cells in a selectin-dependent manner. Surgery 2013, 154, 69–77. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Chattarji, S.; Diamond, D.M.; Jay, T.M.; Reagan, L.P.; Svenningsson, P.; Fuchs, E. The neurobiological propierties of Tianeptine (Stablon): From monoamine hypothesis to glutamtergic modulation. Mol. Psychiatry 2010, 15, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Uzbay, T.I. Tianeptine: Potential influences on neuroplasticity and novel pharmacological effects. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2008, 32, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, S.D.; Trentani, A.; Van Der Zee, E.A.; Den Boer, J.A. Chronic stress-induced changes in the rat brain: Role of sex differences and effects of long-term tianeptine treatment. Neuropharmacology 2013, 75, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Jantas, D.; Krawczyk, S.; Lason, W. The predominant protective effect of tianeptine over other antidepressants in models of neuronal apoptosis: The effect blocked by inhibitors of MAPK/ERK1/2 and PI3-K/Akt pathways. Neurotox. Res. 2014, 25, 208–225. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Mateo, C.; Darias, V.; Albertos, L.; Expo’sito-Orta, M. Psychopharmacological Effects of Tianeptine Analogous Hetero[2,1] benzothiazepine Derivatives. Arzneimittelforschung 2011, 53, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Muniz, P.S.; Nardi, A.E. Refractory Pain–Depression Syndrome Treated with Tianeptine: A Case Report. Aust. N. Z. J. Psychiatry 2004, 38, 176. [Google Scholar] [CrossRef] [PubMed]
- Castanon, N.; Konsman, J.P.; Médina, C.; Chauvet, N.; Dantzer, R. Chronic treatment with the antidepressant tianeptine attenuates lipopolysaccharide-induced Fos expression in the rat paraventricular nucleus and HPA axis activation. Psychoneuroendocrinology 2003, 28, 19–34. [Google Scholar] [CrossRef]
- Trojan, E.; Głombik, K.; Ślusarczyk, J.; Budziszewska, B.; Kubera, M.; Roman, A.; Lasoń, W.; Basta-Kaim, A. The Beneficial Impact of Antidepressant Drugs on Prenatal Stress-Evoked Malfunction of the Insulin-Like Growth Factor-1 (IGF-1) Protein Family in the Olfactory Bulbs of Adult Rats. Neurotox. Res. 2016, 29, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Trojan, E.; Ślusarczyk, J.; Chamera, K.; Kotarska, K.; Glombik, K.; Kubera, M.; Basta-Kaim, A. The modulatory properties of chronic antidepressant drugs treatment on the brain chemokine—Chemokine receptor network: A molecular study in an animal model of depression. Front. Pharmacol. 2017, 8, 779. [Google Scholar] [CrossRef] [PubMed]
- Ling, E.-A.; Wong, W.-C. The origin and nature of ramified and amoeboid microglia: A historical review and current concepts. Glia 1993, 7, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Rock, R.B.; Gekker, G.; Hu, S.; Sheng, W.S.; Cheeran, M.; Lokensgard, J.R.; Phillip, K.; Peterson, P.K. Role of Microglia in Central Nervous System Infections Role. Clin. Microbiol. Rev. 2004, 17, 942–964. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.P.; Zou, M.; Wang, J.Y.; Zhu, J.J.; Lai, J.M.; Zhou, L.L.; Chen, S.F.; Zhang, X.; Zhu, J.H. Paroxetine ameliorates lipopolysaccharide-induced microglia activation via differential regulation of MAPK signaling. J. Neuroinflamm. 2014, 11, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Wang, Z.; Liu, S.; Wang, F.; Zhao, S.; Hao, A. Anti-inflammatory effects of fluoxetine in lipopolysaccharide (LPS)-stimulated microglial cells. Neuropharmacology 2011, 61, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Obuchowicz, E.; Bielecka, A.M.; Paul-Samojedny, M.; Pudełko, A.; Kowalski, J. Imipramine and fluoxetine inhibit LPS-induced activation and affect morphology of microglial cells in the rat glial culture. Pharmacol. Rep. 2014, 66, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.A.; Boddeke, H.W.G.M.; Kettenmann, H. Microglia in Physiology and Disease. Annu. Rev. Physiol. 2017, 79, 619–643. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting Microglial Cells Are Highly Dynamic Surveillants of Brain Parenchyma In Vivo. Science 2005, 308, 1314–1319. [Google Scholar] [CrossRef] [PubMed]
- Colonna, M.; Butovsky, O. Microglia Function in the Central Nervous System During Health and Neurodegeneration. Annu. Rev. Immunol. 2017, 35, 441–468. [Google Scholar] [CrossRef] [PubMed]
- Kierdorf, K.; Prinz, M. Factors regulating microglia activation. Front. Cell. Neurosci. 2013, 7, 44. [Google Scholar] [CrossRef] [PubMed]
- Salvi, V.; Sozio, F.; Sozzani, S.; Prete, A. Del Role of atypical chemokine receptors in microglial activation and polarization. Front. Aging Neurosci. 2017, 9, 148. [Google Scholar] [CrossRef] [PubMed]
- Andrzejczak, D.; Woldan-Tambor, A.; Bednarska, K.; Zawilska, J.B. The effects of topiramate on lipopolysaccharide (LPS)-induced proinflammatory cytokine release from primary rat microglial cell cultures. Epilepsy Res. 2016, 127, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Stuart, M.J.; Baune, B.T. Chemokines and chemokine receptors in mood disorders, schizophrenia, and cognitive impairment: A systematic review of biomarker studies. Neurosci. Biobehav. Rev. 2014, 42, 93–115. [Google Scholar] [CrossRef] [PubMed]
- Plaisant, F.; Dommergues, M.A.; Spedding, M.; Cecchelli, R.; Brillault, J.; Kato, G.; Muñoz, C.; Gressens, P. Neuroprotective properties of tianeptine: Interactions with cytokines. Neuropharmacology 2003, 44, 801–809. [Google Scholar] [CrossRef]
- Castanon, N.; Bluthé, R.M.; Dantzer, R. Chronic treatment with the atypical antidepressant tianeptine attenuates sickness behavior induced by peripheral but not central lipopolysaccharide and interleukin-1beta in the rat. Psychopharmacology 2001, 154, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Castanon, N.; Médina, C.; Mormède, C.; Dantzer, R. Chronic administration of tianeptine balances lipopolysaccharide-induced expression of cytokines in the spleen and hypothalamus of rats. Psychoneuroendocrinology 2004, 29, 778–790. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric Oxide and Peroxynitrite in Health and Disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, J.P. Role of nitric oxide in the regulation of monoaminergic neurotransmission. Brain Res. Bull. 2000, 52, 459–466. [Google Scholar] [CrossRef]
- Suzuki, E.; Yagi, G.; Nakaki, T.; Kanba, S.; Asai, M. Elevated plasma nitrate levels in depressive states. J. Affect. Disord. 2001, 63, 221–224. [Google Scholar] [CrossRef]
- Araújo, I.M.; Carvalho, C.M. Role of nitric oxide and calpain activation in neuronal death and survival. Curr. Drug Targets. CNS Neurol. Disord. 2005, 4, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Kamata, H.; Hirata, H. Redox regulation of cellular signalling. Cell. Signal. 1999, 11, 1–14. [Google Scholar] [CrossRef]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.; Tresini, M. Oxidative stress and gene regulation. Free Radic. Biol. Med. 2000, 28, 463–499. [Google Scholar] [CrossRef]
- Brown, G.C.; Bal-Price, A. Inflammatory Neurodegeneration Mediated by Nitric Oxide, Glutamate, and Mitochondria. Mol. Neurobiol. 2003, 27, 325–355. [Google Scholar] [CrossRef]
- Brown, G.C.; Neher, J.J. Inflammatory neurodegeneration and mechanisms of microglial killing of neurons. Mol. Neurobiol. 2010, 41, 242–247. [Google Scholar] [CrossRef] [PubMed]
- He, A.; Shao, J.; Zhang, Y.; Lu, H.; Wu, Z.; Xu, Y. CD200Fc reduces LPS-induced IL-1β activation in human cervical cancer cells by modulating TLR4-NF-κB and NLRP3 inflammasome pathway. Oncotarget 2017, 8, 33214–33224. [Google Scholar] [CrossRef] [PubMed]
- Koistinaho, M.; Koistinaho, J. Interactions between Alzheimer’s disease and cerebral ischemia—Focus on inflammation. Brain Res. Rev. 2005, 48, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Liu, W.; Dickson, D.W.; Brosnan, C.F.; Berman, J.W. Cytokine production by human fetal microglia and astrocytes. Differential induction by lipopolysaccharide and IL-1 beta. J. Immunol. 1993, 150, 2659–2667. [Google Scholar] [PubMed]
- Hwang, J.; Zheng, L.T.; Ock, J.; Lee, M.G.; Kim, S.H.; Lee, H.W.; Lee, W.H.; Park, H.C.; Suk, K. Inhibition of glial inflammatory activation and neurotoxicity by tricyclic antidepressants. Neuropharmacology 2008, 55, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Lee, M. Neurotransmitters and Microglial-Mediated Neuroinflammation. Curr. Protein Pept. Sci. 2013, 14, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.Z.; Xu, Z.Q.; Han, B.Z.; Su, D.F.; Liu, C. NLRP3 inflammasome and its inhibitors: A review. Front. Pharmacol. 2015, 6, 262. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, C.Y.; Barberi, T.J.; Ghosh, P.; Longo, D.L. Phosphorylation of Re1A/p65 on serine 536 defines an IκBα-independent NF-κB pathway. J. Biol. Chem. 2005, 280, 34538–34547. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Hu, L.; Li, D.; Wang, Y.; He, Y.; Zhu, L.; Ren, W. MD-2 regulates LPS-induced NLRP3 inflammasome activation and IL-1beta secretion by a MyD88/NF-κB-dependent pathway in alveolar macrophages cell line. Mol. Immunol. 2017, 90, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, E.; Rodhe, J.; Burguillos, M.A.; Venero, J.L.; Joseph, B. Regulation of caspase-3 processing by cIAP2 controls the switch between pro-inflammatory activation and cell death in microglia. Cell Death Dis. 2014, 5, e1565–e1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, E.; Campbell, M.; Doyle, S.L. Targeting the NLRP3 inflammasome in chronic inflammatory diseases: Current perspectives. J. Inflamm. Res. 2015, 8, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Guo, H.; Xu, C.; Wang, B.; Zhang, M.; Ding, F. Mitochondrial reactive oxygen species-mediated NLRP3 inflammasome activation contributes to aldosterone-induced renal tubular cells injury. Oncotarget 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, F.; Bartok, E.; Rieger, A.; Franchi, L.; Nunez, G.; Hornung, V. Cutting Edge: Reactive Oxygen Species Inhibitors Block Priming, but Not Activation, of the NLRP3 Inflammasome. J. Immunol. 2011, 187, 613–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leskiewicz, M.; Regulska, M.; Budziszewska, B.; Jantas, D.; Jaworska-Feil, L.; Basta-Kaim, A.; Kubera, M.; Jagla, G.; Nowak, W.; Lason, W. Effects of neurosteroids on hydrogen peroxide- and staurosporine-induced damage of human neuroblastoma SH-SY5Y cells. J. Neurosci. Res. 2008, 86, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Basta-Kaim, A.; Budziszewska, B.; Leśkiewicz, M.; Fijał, K.; Regulska, M.; Kubera, M.; Wȩdzony, K.; Lasoń, W. Hyperactivity of the hypothalamus-pituitary-adrenal axis in lipopolysaccharide-induced neurodevelopmental model of schizophrenia in rats: Effects of antipsychotic drugs. Eur. J. Pharmacol. 2011, 650, 586–595. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Gene Expression | |||
|---|---|---|---|---|
| Control | LPS | Control + TIA10 | LPS + TIA10 | |
| MHCII | 1.00 ± 0.06 | 2.38 ± 0.29 * | 1.00 ± 0.03 | 1.30 ± 0.07 # |
| CD40 | 1.00 ± 0.06 | 2.34 ± 0.15 * | 1.02 ± 0.04 | 1.46 ± 0.05 # |
| IL-1β | 1.00 ± 0.04 | 3.34 ± 0.32 * | 1.03 ± 0.03 | 1.48 ± 0.23 # |
| IL-18 | 1.00 ± 0.07 | 3.06 ± 0.27 * | 0.96 ± 0.08 | 1.39 ± 0.07 # |
| TNF-α | 1.00 ± 0.04 | 4.50 ± 0.43 * | 0.94 ± 0.05 | 2.06 ± 0.32 # |
| IL-6 | 1.00 ± 0.03 | 1.51 ± 0.10 * | 0.97 ± 0.03 | 1.26 ± 0.04 # |
| CCL2 | 1.00 ± 0.09 | 2.69 ± 0.31 * | 1.06 ± 0.09 | 1.37 ± 0.12 # |
| IL-4 | 1.00 ± 0.04 | 0.73 ± 0.05 | 1.78 ± 0.82 | 1.01 ± 0.16 |
| IL-10 | 1.01 ± 0.06 | 8.68 ± 1.31 * | 0.69 ± 0.15 | 9.74 ± 1.49 * |
| TGF-β | 1.00 ± 0.03 | 0.37 ± 0.05 * | 0.58 ± 0.05 * | 0.58 ± 0.01 * |
| IGF-1 | 1.06 ± 0.16 | 0.0081 ± 0.0022 * | 0.67 ± 0.19 | 0.0242 ± 0.0032 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ślusarczyk, J.; Trojan, E.; Głombik, K.; Piotrowska, A.; Budziszewska, B.; Kubera, M.; Popiołek-Barczyk, K.; Lasoń, W.; Mika, J.; Basta-Kaim, A. Targeting the NLRP3 Inflammasome-Related Pathways via Tianeptine Treatment-Suppressed Microglia Polarization to the M1 Phenotype in Lipopolysaccharide-Stimulated Cultures. Int. J. Mol. Sci. 2018, 19, 1965. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071965
Ślusarczyk J, Trojan E, Głombik K, Piotrowska A, Budziszewska B, Kubera M, Popiołek-Barczyk K, Lasoń W, Mika J, Basta-Kaim A. Targeting the NLRP3 Inflammasome-Related Pathways via Tianeptine Treatment-Suppressed Microglia Polarization to the M1 Phenotype in Lipopolysaccharide-Stimulated Cultures. International Journal of Molecular Sciences. 2018; 19(7):1965. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071965
Chicago/Turabian StyleŚlusarczyk, Joanna, Ewa Trojan, Katarzyna Głombik, Anna Piotrowska, Bogusława Budziszewska, Marta Kubera, Katarzyna Popiołek-Barczyk, Władysław Lasoń, Joanna Mika, and Agnieszka Basta-Kaim. 2018. "Targeting the NLRP3 Inflammasome-Related Pathways via Tianeptine Treatment-Suppressed Microglia Polarization to the M1 Phenotype in Lipopolysaccharide-Stimulated Cultures" International Journal of Molecular Sciences 19, no. 7: 1965. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071965