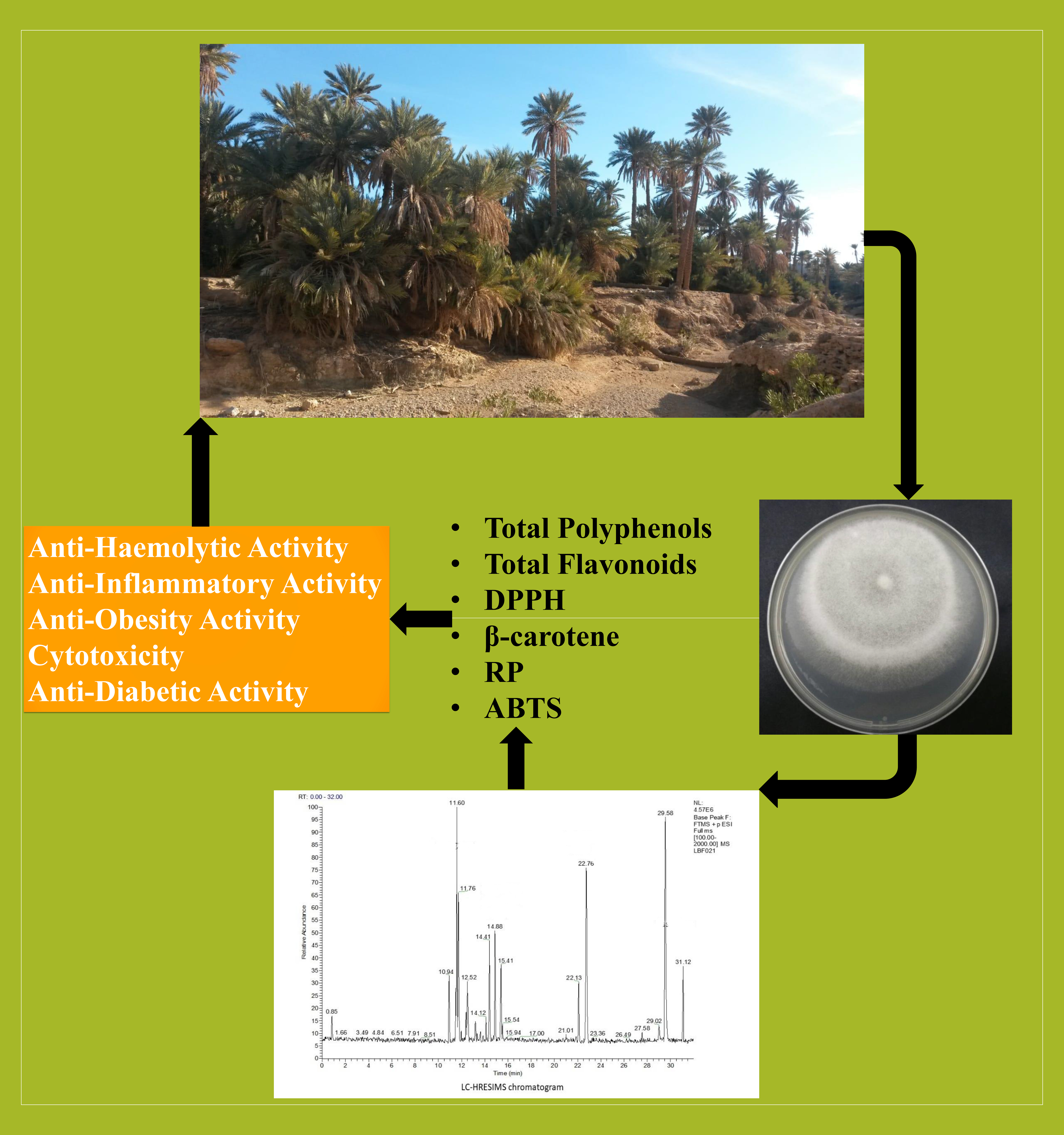
Date Palm Trees Root-Derived Endophytes as Fungal Cell Factories for Diverse Bioactive Metabolites


 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Results
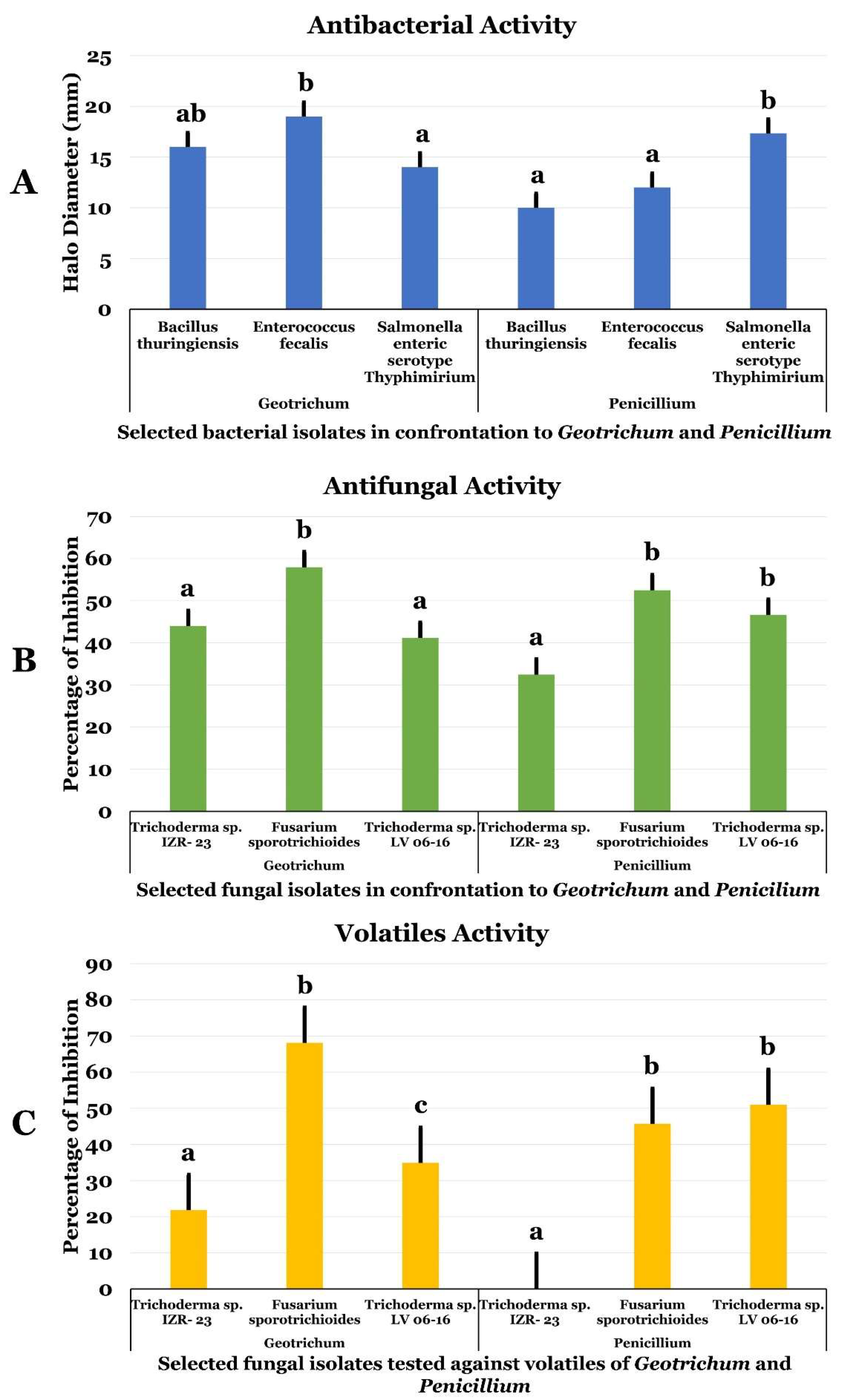
2.1. Antimicrobial (Antagonistic) Activities of G. candidum and P. citrinum and Their Volatiles
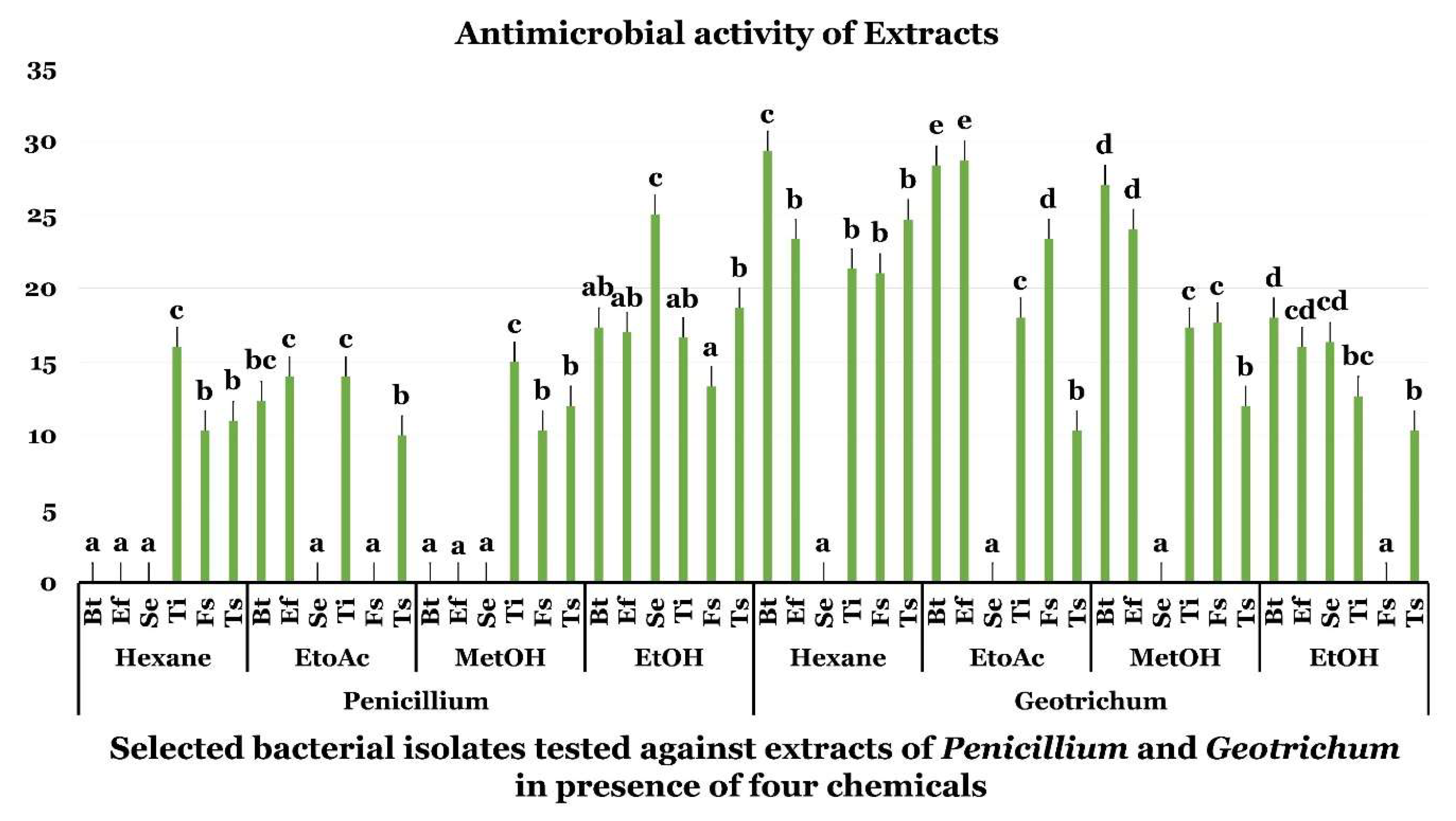
2.2. Antimicrobial Assay of G. candidum and P. citrinum Extracts
2.3. Biological Activities of G. candidum and P. citrinum Extracts
2.4. Chemical Composition of G. candidum and P. citrinum Extracts
Polyphenol and Flavonoid Contents
2.5. In Vitro Antioxidant Activity of Date Palm Endophytic Fungi
2.5.1. DPPH Free Radical-Scavenging Activity
2.5.2. β-Carotene-Linoleic Acid Assay
2.5.3. Reducing Power Assay
2.5.4. ABTS Radical-Scavenging Activity
2.6. LC-HRMS Analysis of G. candidum and P. citrinum Extracts
2.7. GC-MS Analysis of G. candidum and P. citrinum Extracts
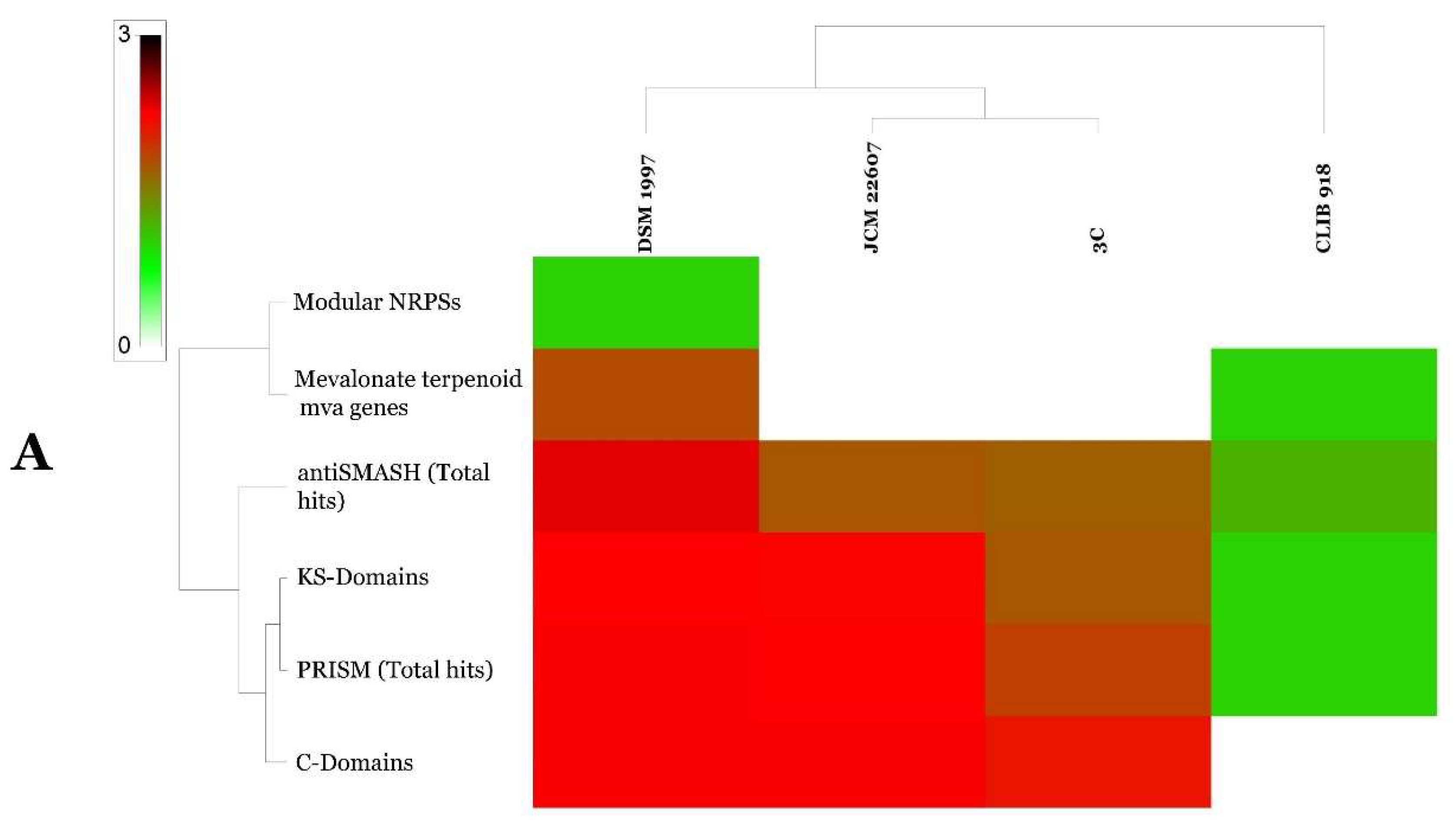
2.8. Genome Mining of G. candidum and P. citrinum Strains
3. Discussion
4. Materials and Methods
4.1. Antimicrobial Screening G. candidum and P. citrinum Fungi and Their Extracts
4.1.1. Microbial Strains and Growth Conditions
4.1.2. Antagonistic Assay of G. candidum and P. citrinum Fungi
4.1.3. Antagonistic Assay of G. candidum and P. citrinum Volatiles
4.1.4. Fermentation and Preparation of Extracts of G. candidum and P. citrinum
4.1.5. Antimicrobial Activity of G. candidum and P. citrinum Extracts
4.2. In Vitro Biological Activities of G. candidum and P. citrinum Extracts
4.2.1. Cytotoxic Activity
4.2.2. Anti-Inflammatory Activity
4.2.3. Anti-Haemolytic Activity
4.2.4. Anti-Diabetic Activity
4.2.5. Anti-Obesity Activity
4.3. In Vitro Assay of Antioxidant Activity of G. candidum and P. citrinum Extracts
4.3.1. DPPH Free Radical-Scavenging Activity
4.3.2. β-Carotene-Linoleic Acid Assay
4.3.3. Reducing Power Assay
4.3.4. ABTS Radical-Scavenging Activity
4.4. Phytochemical Composition of G. candidum and P. citrinum Extracts
4.4.1. Total Phenolic and Flavonoid Content Assay
4.4.2. Samples Preparation for GCMS and LCMS Analysis
4.4.3. LC-MS Analysis of G. candidum and P. citrinum Extracts
4.4.4. GC-MS Analysis of G. candidum and P. citrinum Cultures
4.4.5. Genome Mining of G. candidum and P. citrinum Strains
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mefteh, F.B.; Daoud, A.; Chenari Bouket, A.; Alenezi, F.N.; Luptakova, L.; Rateb, M.E.; Kadri, A.; Gharsallah, N.; Belbahri, L. Fungal root microbiome from healthy and brittle leaf diseased date palm trees (Phoenix dactylifera L.) reveals a hidden untapped arsenal of antibacterial and broad spectrum antifungal secondary metabolites. Front. Microbiol. 2017, 8, 307. [Google Scholar] [CrossRef] [PubMed]
- Cairns, T.; Vera, M. In silico prediction and characterization of secondary metabolite biosynthetic gene clusters in the wheat pathogen Zymoseptoria tritici. BMC Genom, 2017, 18, 631. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, I.K.; Chettri, P.; Dupont, P.Y.; Barnes, I.; McDougal, R.L.; Moore, G.G.; Sim, A.; Bradshaw, R.E. Evolution of polyketide synthesis in a Dothideomycete forest pathogen. Fungal Genet. Biol. 2017, 106, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Sharma, P.; Kumar, A.; Chadha, P.; Kaur, R.; Kaur, A. Antioxidant and in vivo genoprotective effects of phenolic compounds identified from an endophytic Cladosporium velox and their relationship with its host plant Tinospora cordifolia. J. Ethnopharmacol. 2016, 194, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Arora, D.S.; Chandra, P. Antioxidant Activity of Aspergillus fumigatus. ISRN Pharmacol. 2011, 2011, 619395. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Fusetani, N. Marine pharmacology in 2012–2013: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar. Drugs 2017, 15, 273. [Google Scholar] [CrossRef]
- Sharma, M.; Sharma, R. Drugs and drug intermediates from fungi: Striving for greener processes. Crit. Rev. Microbiol. 2016, 42, 322–338. [Google Scholar] [CrossRef] [PubMed]
- Noinart, J.; Buttachon, S.; Dethoup, T.; Gales, L.; Pereira, J.A.; Urbatzka, R.; Freitas, S.; Lee, M.; Silva, A.M.S.; Pinto, M.M.M.; et al. A new ergosterol analog, a new bis-anthraquinone and anti-obesity activity of anthraquinones from the marine sponge-associated fungus Talaromyces stipitatus KUFA 0207. Mar. Drugs 2017, 15, 139. [Google Scholar] [CrossRef] [PubMed]
- Li, C.S.; Yang, B.J.; Turkson, J.; Cao, S.G. Anti-proliferative ambuic acid derivatives from Hawaiian endophytic fungus Pestalotiopsis sp. FT172. Phytochemistry 2017, 140, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Katoch, M.; Phull, S.; Vaid, S.; Singh, S. Diversity, phylogeny, anticancer and antimicrobial potential of fungal endophytes associated with Monarda citriodora L. BMC Microbiol. 2017, 17, 44. [Google Scholar] [CrossRef] [PubMed]
- Jahn, L.; Schafhauser, T.; Wibberg, D.; Ruckert, C.; Winkler, A.; Kulik, A.; Weber, T.; Flor, L.; van Pee, K.H.; Kalinowski, J.; et al. Linking secondary metabolites to biosynthesis genes in the fungal endophyte Cyanodermella asteris: The anti-cancer bisanthraquinone skyrin. J. Biotechnol. 2017, 257, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Monggoot, S.; Popluechai, S.; Gentekaki, E.; Pripdeevech, P. Fungal endophytes: An alternative source for production of volatile compounds from agarwood oil of Aquilaria subintegra. Microb. Ecol. 2017, 74, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Klimova, E.; Rodriguez-Pena, K.; Sanchez, S. Endophytes as sources of antibiotics. Biochem. Pharmacol. 2017, 134, 1–17. [Google Scholar] [CrossRef] [PubMed]
- McMullin, D.R.; Green, B.D.; Prince, N.C.; Tanney, J.B.; Miller, J.D. Natural products of Picea endophytes from the Acadian forest. J. Nat. Prod. 2017, 80, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Chhipa, H.; Kaushik, N. Fungal and bacterial diversity isolated from Aquilaria malaccensis tree and soil, induces agarospirol formation within 3 months after artificial infection. Front. Microbiol. 2017, 8, 1286. [Google Scholar] [CrossRef] [PubMed]
- Belbahri, L.; Chenari Bouket, A.; Rekik, I.; Alenezi, F.N.; Vallat, A.; Luptakova, L.; Petrovova, E.; Oszako, T.; Cherrad, S.; Vacher, S.; et al. Comparative genomics of Bacillus amyloliquefaciens strains reveals a core genome with traits for habitat adaptation and a secondary metabolites rich accessory genome. Front. Microbiol. 2017, 8, 1438. [Google Scholar] [CrossRef] [PubMed]
- Bailly, A.; Weisskopf, L. Mining the volatilomes of plant-associated microbiota for new biocontrol solutions. Front. Microbiol. 2017, 8, 1638. [Google Scholar] [CrossRef] [PubMed]
- Grijseels, S.; Nielsen, J.C.; Nielsen, J.; Larsen, T.O.; Frisvad, J.C.; Nielsen, K.F.; Frandsen, R.J.N.; Workman, M. Physiological characterization of secondary metabolite producing Penicillium cell factories. Fungal Biol. Biotechnol. 2017, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.N.; Liang, Y.J.; Yang, Y.; Lu, X.F. Single-step production of the simvastatin precursor monacolin J by engineering of an industrial strain of Aspergillus terreus. Metab. Eng. 2017, 42, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Lubeck, M.; Lubeck, P.S. Aspergillus as a versatile cell factory for organic acid production. Fungal Biol. Rev. 2017, 31, 33–49. [Google Scholar] [CrossRef]
- Pardo-Planas, O.; Prade, R.A.; Muller, M.; Atiyeh, H.K. Prevention of melanin formation during aryl alcohol oxidase production under growth-limited conditions using an Aspergillus nidulans cell factory. Bioresour. Technol. 2017, 243, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Anyaogu, D.C.; Mortensen, U.H. Heterologous production of fungal secondary metabolites in Aspergilli. Front. Microbiol. 2015, 6, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, H.U.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.A.; Müller, R.; Wohlleben, W.; et al. antiSMASH 3.0—A comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res. 2015, 43, W237–W243. [Google Scholar] [CrossRef] [PubMed]
- Skinnider, M.A.; Dejong, C.A.; Rees, P.N.; Johnston, C.W.; Li, H.; Webster, A.L.; Wyatt, M.A.; Magarvey, N.A. Genomes to natural products prediction informatics for secondary metabolomes (PRISM). Nucleic Acids Res. 2015, 43, 9645–9662. [Google Scholar] [CrossRef] [PubMed]
- Ziemert, N.; Podell, S.; Penn, K.; Badger, J.H.; Allen, E.; Jensen, P.R. The natural product domain seeker NaPDoS: A phylogeny based bioinformatic tool to classify secondary metabolite gene diversity. PLoS ONE 2012, 7, e34064. [Google Scholar] [CrossRef] [PubMed]
- Li, M.H.T.; Ung, P.M.U.; Zajkowski, J.; Garneau-Tsodikova, S.; Sherman, D.H. Automated genome mining for natural products. BMC Bioinform. 2009, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- Van Heel, A.J.; de Jong, A.; Montalban-Lopez, M.; Kok, J.; Kuipers, O.P. BAGEL3: Automated identification of genes encoding bacteriocins and (non-)bactericidal post translationally modified peptides. Nucleic Acids Res. 2013, 41, W448–W453. [Google Scholar] [CrossRef] [PubMed]
- Clevenger, K.D.; Bok, J.W.; Ye, R.; Miley, G.P.; Verdan, M.H.; Velk, T.; Chen, C.; Yang, K.; Robey, M.T.; Gao, P.; et al. A scalable platform to identify fungal secondary metabolites and their gene clusters. Nat. Chem. Biol. 2017, 13, 895–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandel, S.L.; Firrincieli, A.; Joubert, P.M.; Okubara, P.A.; Leston, N.D.; McGeorge, K.M.; Mugnozza, G.S.; Harfouche, A.; Kim, S.-H.; Doty, S.L. An In vitro study of bio-control and plant growth promotion potential of Salicaceae endophytes. Front. Microbiol. 2017, 8, 386. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Shen, Z.; Ruan, Y.; Tao, C.; Chao, Y.; Li, R.; Shen, Q. Isolation of antagonistic endophytes from banana roots against Meloidogyne javanica and their effects on soil nematode community. Front. Microbiol. 2017, 8, 2070. [Google Scholar] [CrossRef] [PubMed]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.-S.; Patra, J.K. Endophytes: A treasure house of bioactive compounds of medicinal importance. Front. Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef] [PubMed]
- Jalgaonwala, R.E.; Mohite, B.V.; Mahajan, R.T. A review: Natural products from plant associated endophytic fungi. J. Microbiol. Biotechnol. Res. 2011, 1, 21–32. [Google Scholar]
- Nair, D.N.; Padmavathy, S. Impact of endophytic microorganisms on plants, environment and humans. Sci. World J. 2014, 250693. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Tan, Y.; Liu, N.; Liao, Y.; Sun, C.; Wang, S.; Wu, A. Functional agents to biologically control deoxynivalenol contamination in cereal grains. Front. Microbiol. 2016, 7, 395. [Google Scholar] [CrossRef] [PubMed]
- Gos, F.M.W.R.; Savi, D.C.; Shaaban, K.A.; Thorson, J.S.; Aluizio, R.; Possiede, Y.M.; Rohr, J.; Glienke, C. Antibacterial activity of endophytic Actinomycetes isolated from the medicinal plant Vochysia divergens (Pantanal, Brazil). Front. Microbiol. 2017, 8, 1642. [Google Scholar] [CrossRef] [PubMed]
- Bibi, Y.; Nisa, S.; Zia, M.; Waheed, A.; Ahmed, S.; Chaudhary, M.F. The study of anticancer and antifungal activities of Pistacia integerrima extract in vitro. Indian J. Pharm. Sci. 2012, 74, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Daoud, A.; Ben Mefteh, F.; Mnafgui, K.; Turki, M.; Jmal, S.; Ben Amar, R.; Ayadi, F.; ElFeki, A.; Abid, L.; Rateb, M.E.; et al. Cardiopreventive effect of ethanolic extract of date palm pollen against isoproterenol induced myocardial infarction in rats through the inhibition of the angiotensin-converting enzyme. Exp. Toxicol. Pathol. 2017, 69, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Patil, D.; Rajamohanan, P.R.; Ahmad, A. Isolation, purification and characterization of vinblastine and vincristine from endophytic fungus Fusarium oxysporum isolated from Catharanthus roseus. PLoS ONE 2013, 8, e71805. [Google Scholar] [CrossRef] [PubMed]
- Wattana-Amorn, P.; Charoenwongsa, W.; Williams, C.; Crump, M.P.; Apichaisataienchote, B. Antibacterial activity of cyclo(L-Pro-L-Tyr) and cyclo(D-Pro-L-Tyr) from Streptomyces sp. strain 22-4 against phytopathogenic bacteria. Nat. Prod. Res. 2016, 30, 1980–1983. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.A.; Sharma, S.; Mittra, S.; Sujatha, S.; Kanaujia, A.; Shukla, G.; Katiyar, C.; Lakshmi, B.S.; Bansal, V.S.; Bhatnagar, P.K. Antihyperglycemic effect of Annona squamosa hexane extract in type 2 diabetes animal model: PTP1B inhibition, a possible mechanism of action? Indian J. Pharmacol. 2012, 44, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Dutraa, F.F.; Alvesa, L.S.; Rodriguesa, D.; Fernandez, P.L.; de Oliveirac, R.B.; Golenbockc, D.T.; Zambonid, D.S.; Bozza, M.T. Hemolysis-induced lethality involves inflammasome activation by heme. Proc. Natl. Acad. Sci. USA 2014, E4110–E4118. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-S.; Wang, S.-W.; Wang, G.-J.; Pang, K.-L.; Lee, C.-K.; Kuo, Y.-H.; Cha, H.-J.; Lin, R.-K.; Lee, T.-H. Angiogenesis inhibitors and anti-Inflammatory agents from Phoma sp. NTOU4195. J. Nat. Prod. 2016, 79, 2983–2990. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, Q.; Liu, B.; Xie, C.-L.; Cao, M.-J.; Yang, X.-W.; Liu, G.-M. Inhibitory activities of compounds from the marine actinomycete Williamsia sp. MCCC 1A11233 variant on IgEmediated mast cells and passive cutaneous anaphylaxis. J. Agric. Food Chem. 2017, 65, 10749–10756. [Google Scholar] [CrossRef] [PubMed]
- Alberts, A.W.; Chen, J.; Kuron, G.; Hunt, V.; Huff, J.; Hoffman, C.; Rothrock, J.; Lopez, M.; Joshua, H.; Harris, E.; et al. Mevinolin: A highly potent competitive inhibitor of hydroxymethylglutaryl-coenzyme a reductase and a cholesterol-lowering agent. Proc. Natl. Acad. Sci. USA 1980, 77, 3957–3961. [Google Scholar] [CrossRef] [PubMed]
- Jouda, J.-B.; Tamokou, J.-D.; Mbazoa, C.D.; Douala-Meli, C.; Sarkar, P.; Bag, P.K.; Wandji, J. Antibacterial and cytotoxic cytochalasins from the endophytic fungus Phomopsis sp. harbored in Garcinia kola (Heckel) nut. BMC Complement. Altern. Med. 2016, 16, 462. [Google Scholar] [CrossRef] [PubMed]
- Ebada, M.E. Cuminaldehyde: A potential drug candidate. Pharmacol. Clin. Res. 2017, 2, 555585. [Google Scholar] [CrossRef]
- Quintans-Junior, L.J.; Barreto, R.S.; Menezes, P.P.; Almeida, J.R.; Viana, A.F.; Oliveira, R.C.; Oliveira, A.P.; Gelain, D.P.; de Lucca Júnior, W.; Araújo, A.A. β-Cyclodextrin-complexed (-)-linalool produces antinociceptive effect superior to that of (-)-linalool in experimental pain protocols. Basic Clin. Pharmacol. Toxicol. 2013, 113, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.; Xiao, J.G.; Pei, G.X. Medicinal Plants: Chemistry, Biology and Omics; Elsevier Science: New York, NY, USA, 2015; p. 694. [Google Scholar]
- Mangamuri, U.K.; Muvva, V.; Poda, S.; Manavathi, B.; Bhujangarao, C.; Yenamandra, V. Chemical characterization and bioactivity of diketopiperazine derivates from the mangrove derived Pseudonocardia endophytica. Egypt. J. Aquat. Res. 2016, 42, 169–175. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed]
- Hautbergue, T.; Jamin, E.L.; Debrauwer, L.; Puel, O.; Oswald, I.P. From genomics to metabolomics, moving toward an integrated strategy for the discovery of fungal secondary metabolites. Nat. Prod. Rep. 2018, 35, 147–173. [Google Scholar] [CrossRef] [PubMed]
- Alenezi, F.N.; Rekik, I.; Belka, M.; Ibrahim, A.F.; Luptakova, L.; Jaspars, M.; Woodward, S.; Belbahri, L. Strain-level diversity of secondary metabolism in the biocontrol species Aneurinibacillus migulanus. Microbiol. Res. 2016, 182, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Medina-Romero, Y.M.; Roque-Flores, G.; Macias-Rubalcava, M.L. Volatile organic compounds from endophytic fungi as innovative postharvest control of Fusarium oxysporum in cherry tomato fruits. Appl. Microbiol. Biotechnol. 2017, 101, 8209–8222. [Google Scholar] [CrossRef] [PubMed]
- Schalchli, H.; Hormazabal, E.; Becerra, J.; Birkett, M.; Alvear, M.; Vidal, J.; Quiroz, A. Antifungal activity of volatile metabolites emitted by mycelial cultures of saprophytic fungi. Chem. Ecol. 2011, 27, 503–513. [Google Scholar] [CrossRef]
- Zhao, J.T.; Ma, D.H.; Luo, M.; Wang, W.; Zhao, C.J.; Zu, Y.G.; Fu, Y.J.; Wink, M. In vitro antioxidant activities and antioxidant enzyme activities in HepG2 cells and main active compounds of endophytic fungus from pigeon pea [Cajanus cajan (L.) Millsp.]. Food Res. Int. 2014, 56, 243–251. [Google Scholar] [CrossRef]
- Sakat, S.; Juvekar, A.R.; Gambhire, M.N. In vitro antioxidant and anti-inflammatory activity of methanol extract of Oxalis corniculata Linn. Int. J. Pharm. Pharm. Sci. 2010, 2, 146–155. [Google Scholar]
- Hasnat, M.; Mehnaz, P.; Beong, O.L. Acetylcholinesterase inhibition and in vitro and in vivo antioxidant activities of Ganoderma lucidum grown on germinated brown rice. Molecules 2013, 18, 6663–6678. [Google Scholar] [CrossRef] [PubMed]
- Sathiavelu, A.; Sangeetha, S.; Archit, R.; Mythili, S. In vitro anti-diabetic activity of aqueous extract of the medicinal plants Nigella sativa, Eugenia jambolana, Andrographis paniculata and Gymnema sylvestre. Int. J. Drug Dev. Res. 2013, 5, 323–328. [Google Scholar]
- Jeong, S.M.; Kim, S.Y.; Kim, D.R.; Namk, C.; Ahn, D.U.; Lee, S.C. Effect of seed roasting conditions on the antioxidant activity of defatted sesame meal extracts. J. Food Sci. 2004, 69, C377–C381. [Google Scholar] [CrossRef]
- Kabouche, A.; Kabouche, Z.; Ozturk, M.; Kolal, U.; Topcu, G. Antioxidant abietane diterpenoids from Salvia barrelieri. Food. Chem. 2007, 102, 1281–1287. [Google Scholar] [CrossRef]
- Gao, Y.; Zhao, J.; Zu, Y.; Fu, Y.; Liang, L.; Luo, M.; Wang, W.; Efferth, T. Antioxidant properties, superoxide dismutase and glutathione reductase activities in HepG2 cells with a fungal endophyte producing apigenin from pigeon pea [Cajanus cajan (L.) Millsp.]. Food Res. Int. 2012, 49, 147–152. [Google Scholar] [CrossRef]
- Zhao, X.; Sun, H.; Hou, A.; Zhao, Q.; Wei, T.; Xin, W. Antioxidant properties of two gallotannins isolated from the leaves of Pistacia weinmannifolia. Biochim. Biophys. Acta 2005, 1725, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Suksomtip, M.; Pongsamart, S. Protective effect against oxidation of human low-density lipoprotein and plasmid DNA strand scission of tamarind seed coat extract in vitro. LWT-Food Sci. Technol. 2008, 41, 2002–2007. [Google Scholar] [CrossRef]
- Djeridane, A.; Yousfi, M.; Nadjemi, B.; Boutassouna, D.; Stocker, P.; Vidal, N. Antioxidant activity of some Algerian medicinal plants extracts containing phenolic compounds. Food Chem. 2006, 97, 654–660. [Google Scholar] [CrossRef]
- Ozer, E.A.; Allen, J.P.; Hauser, A.R. Characterization of the core and accessory genomes of Pseudomonas aeruginosa using bioinformatic tools Spine and AGEnt. BMC Genom. 2014, 15, 737. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
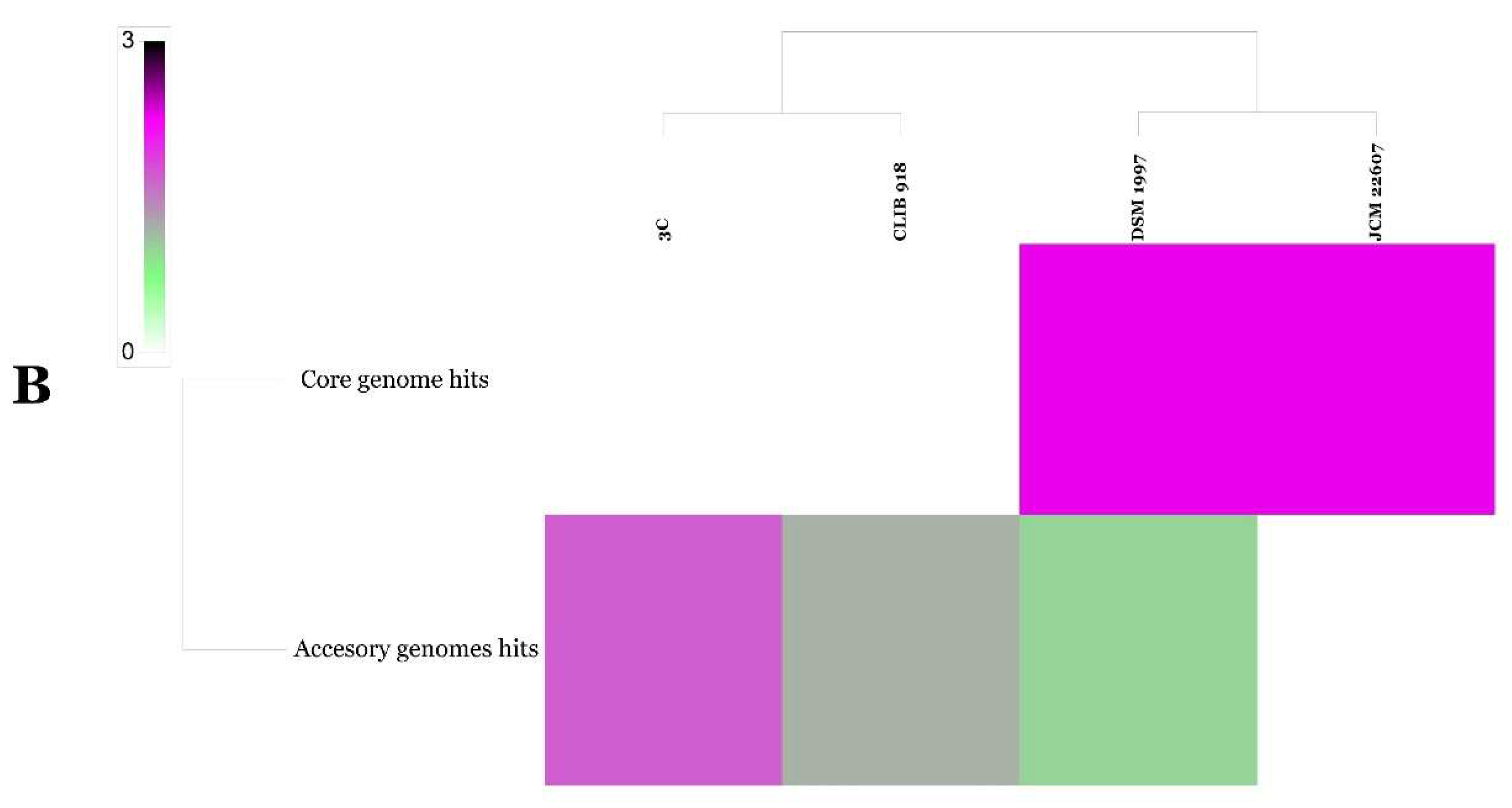{kind=link}
| Geotrichum candidum | |||||
| Extracts | IC50 (mg/mL) | IC50 (µg/mL) | |||
| Anti-Haemolytic Activity | Anti-Inflammatory Activity | Anti-Obesity Activity | Cytotoxicity | Anti-Diabetic Activity | |
| EtOH | 1.53 ± 0.17 (a) | 1.37 ± 0.08 (b) | 170.29 ± 0.83 (c) | - | 162.56 ± 2.91 (d) |
| EtOAc | 1.35 ± 0.07 (a) | 0.47 ± 0.006 (a) | 160.08 ± 0.86 (b) | 145.55 ± 23.55 (b) | 118.84 ± 0.83 (b) |
| Hexane | 0.96 ± 0.03 (a) | 2.3 ± 0.08 (c) | 169.47 ± 1.04 (c) | 163.73 ± 1.76 (d) | |
| MetOH | 2.47 ± 0.24 (a) | 4.31 ± 0.3 (d) | 222.45 ± 4.29 (d) | 177.97 ± 22.94 (b) | 147.36 ± 1.38 (c) |
| Acarbose 1 | - | - | - | - | 17.94 ± 0.33 (a) |
| Aspirin 2 | 0.34 ± 0.1 (b) | - | - | - | - |
| Sodium Diclofenac 3 | - | 0.41 ± 0.01 (a) | - | - | - |
| Orlistat 4 | - | - | 12.86 ± 0.05 (a) | - | - |
| Paclitaxel 5 | - | - | - | 0.52 ± 0.03 (a) | - |
| Penicillium citrinum | |||||
| Extracts | IC50 (mg/mL) | IC50 (µg/mL) | |||
| Anti-Haemolytic Activity | Anti-Inflammatory Activity | Anti-Obesity Activity | Cytotoxicity | Anti-Diabetic Activity | |
| EtOH | 5.25 ± 1.84 (a) | 1.88 ± 0.15 (b) | 175.83 ± 1.34 (b) | - | 188.69 ± 1.66 (c) |
| EtOAc | 2.55 ± 0.42 (a) | 3.67 ± 0.32 (d) | 173.87 ± 0.35 (b) | - | 177.27 ± 2.09 (b) |
| Hexane | 1.64 ± 0.15 (a) | 2.32 ± 0.36 (bc) | 314.29 ± 7.46 (d) | - | 329.73 ± 4.37 (d) |
| MetOH | 1.55 ± 0.12 (a) | 2.64 ± 0.08 (c) | 210.14 ± 4.47 (c) | - | 174.79 ± 3.97 (b) |
| Acarbose 1 | - | - | - | - | 17.94 ± 0.33 (a) |
| Aspirin 2 | 0.34 ± 0.1 (b) | - | - | - | - |
| Diclofenac Sodium Diclofenac 3 | - | 0.41 ± 0.01 (a) | - | - | - |
| Orlistat 4 | - | - | 12.86 ± 0.05 (a) | - | - |
| Paclitaxel 5 | - | - | - | 0.52 ± 0.03 | - |
| Positive controls | |||||
| Geotrichum candidum | ||||||
| Extracts | Total Polyphenols (mg GAE/g) | Total Flavonoids (mg QE/g) | IC50 (µg/mL) | |||
| DPPH | β-carotene | RP | ABTS | |||
| EtOH | 28.93 ± 0.89 (ab) | 14.41 ± 0.16 (c) | 222.16 ± 3.08 (d) | 219.14 ± 1.02 (d) | 211.48 ± 0.37 (c) | 191.36 ± 0.41 (e) |
| EtOAc | 33.39 ± 0.49 (b) | 11.25 ± 1.03 (ab) | 177.55 ± 0.96 (b) | 162.86 ± 0.34 (b) | 190.3 ± 3.36 (b) | 151.31 ± 4.2 (c) |
| Hexane | 27.51 ± 2.78 (a) | 10.03 ± 0.27 (a) | 223.14 ± 2.71 (d) | 220.27 ± 2.07 (d) | 218.71 ± 3.24 (d) | 205.98 ± 8.35 (f) |
| MetOH | 31.77 ± 2.23 (ab) | 12.46 ± 0.6 (b) | 207.44 ± 1.48 (c) | 171.75 ± 1.08 (c) | 205.93 ± 0.41 (c) | 177.9 ± 1.71 (d) |
| BHT 1 | - | - | 34.33 ± 0.03 (a) | 50.11 ± 0.18 (a) | - | - |
| Trolox 2 | - | - | - | - | - | 132.98 ± 0.95 (b) |
| Ascorbic Acid | - | - | - | - | 182.08 ± 1.12 (a) | 116.75 ± 0.32 (a) |
| Penicillium citrinum | ||||||
| Extracts | Total Polyphenols (mg GAE/g) | Total Flavonoids (mg QE/g) | IC50 (µg/mL) | |||
| DPPH | β-carotene | RP | ABTS | |||
| EtOH | 18.87 ± 0.36 (a) | 5.11 ± 0.19 (d) | 268.4 ± 4.46 (d) | 252.84 ± 1.66 (c) | 272.49 ± 4.44 (c) | 266.4 ± 6.41 (e) |
| EtOAc | 21.24 ± 1.4 (ab) | 2.29 ± 0.06 (b) | 246.36 ± 0.45 (c) | 239.42 ± 1.71 (b) | 264.76 ± 4.43 (c) | 227.03 ± 1.37 (d) |
| Hexane | 16.69 ± 1.51 (a) | 1.41 ± 0.33 (a) | 370.8 ± 3.39 (e) | 264.55 ± 2.59 (d) | 301.78 ± 5.81 (d) | 277.02 ± 6.58 (f) |
| MetOH | 24.12 ± 3.36 (b) | 4.07 ± 0.08 (c) | 227.87 ± 0.49 (b) | 235.28 ± 1.15 (b) | 233.89 ± 3.32 (b) | 215.04 ± 1.31 (c) |
| BHT 1 | - | - | 34.33 ± 0.03 (a) | 50.11 ± 0.18 (a) | - | - |
| Trolox 2 | - | - | - | - | - | 132.98 ± 0.95 (b) |
| Ascorbic Acid | - | - | - | - | 182.08 ± 1.12 (a) | 116.75 ± 0.32 (a) |
| Positive control | ||||||
| Query Genome | Reference Genome | Formula 2 (Identities/HSP Length) | ||
|---|---|---|---|---|
| DDH | Distance | Prob. DDH ≥ 70% | ||
| Penicillium citrinum JCM 22607 | Penicillium citrinum DSM 1997 | 97.3 | 0.0040 | 97.7 |
| Geotrichum candidum 3C | Geotrichum candidum CLIB 918 | 18.6 | 0.2367 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ben Mefteh, F.; Daoud, A.; Chenari Bouket, A.; Thissera, B.; Kadri, Y.; Cherif-Silini, H.; Eshelli, M.; Alenezi, F.N.; Vallat, A.; Oszako, T.; et al. Date Palm Trees Root-Derived Endophytes as Fungal Cell Factories for Diverse Bioactive Metabolites. Int. J. Mol. Sci. 2018, 19, 1986. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071986
Ben Mefteh F, Daoud A, Chenari Bouket A, Thissera B, Kadri Y, Cherif-Silini H, Eshelli M, Alenezi FN, Vallat A, Oszako T, et al. Date Palm Trees Root-Derived Endophytes as Fungal Cell Factories for Diverse Bioactive Metabolites. International Journal of Molecular Sciences. 2018; 19(7):1986. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071986
Chicago/Turabian StyleBen Mefteh, Fedia, Amal Daoud, Ali Chenari Bouket, Bathini Thissera, Yamina Kadri, Hafsa Cherif-Silini, Manal Eshelli, Faizah N. Alenezi, Armelle Vallat, Tomasz Oszako, and et al. 2018. "Date Palm Trees Root-Derived Endophytes as Fungal Cell Factories for Diverse Bioactive Metabolites" International Journal of Molecular Sciences 19, no. 7: 1986. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071986