Genome-Wide Analysis, Expression Profile, and Characterization of the Acid Invertase Gene Family in Pepper

Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of CaAINV Genes in Pepper
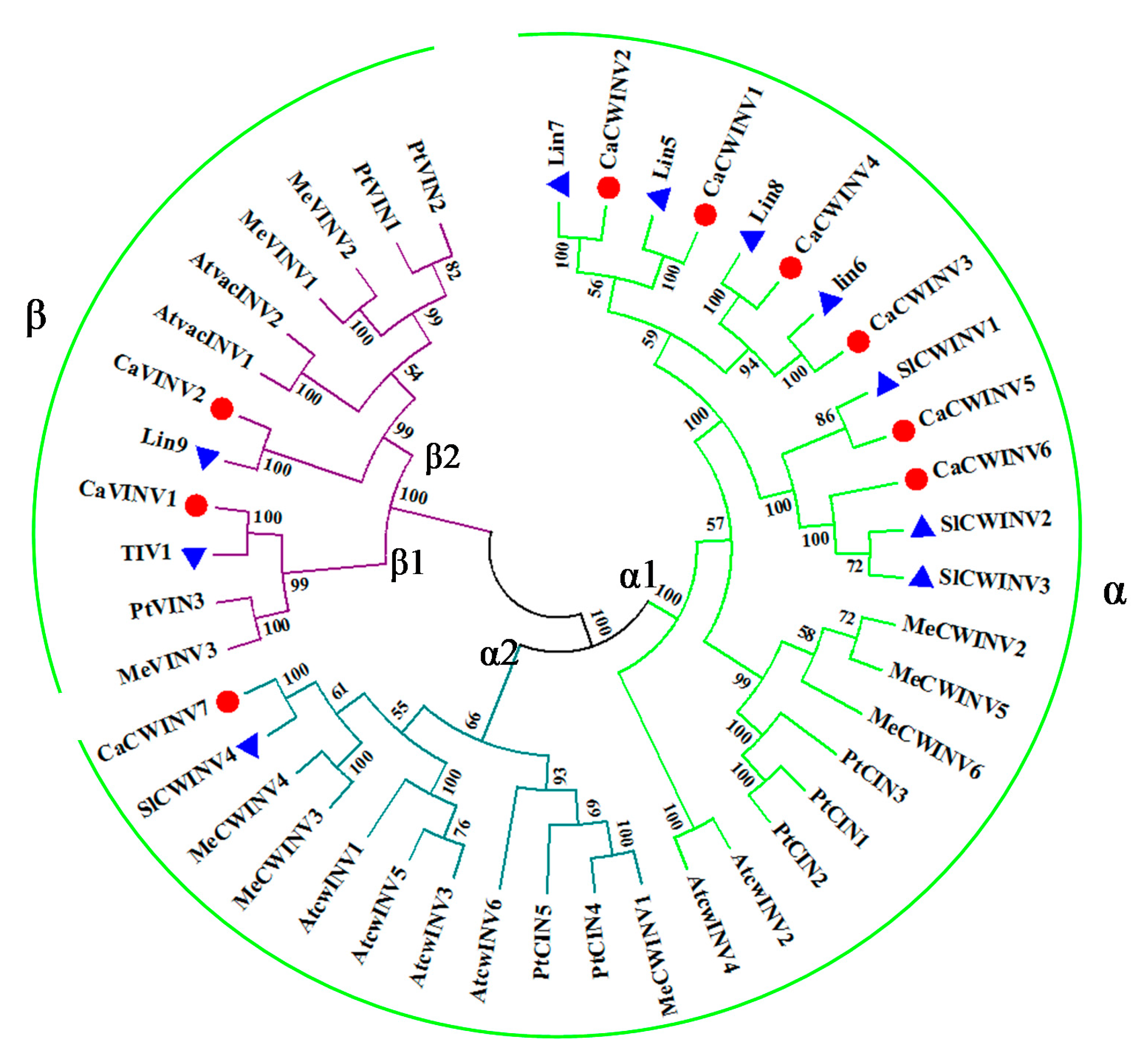
2.2. Phylogenetic Analysis of Acid Invertase Genes in Pepper
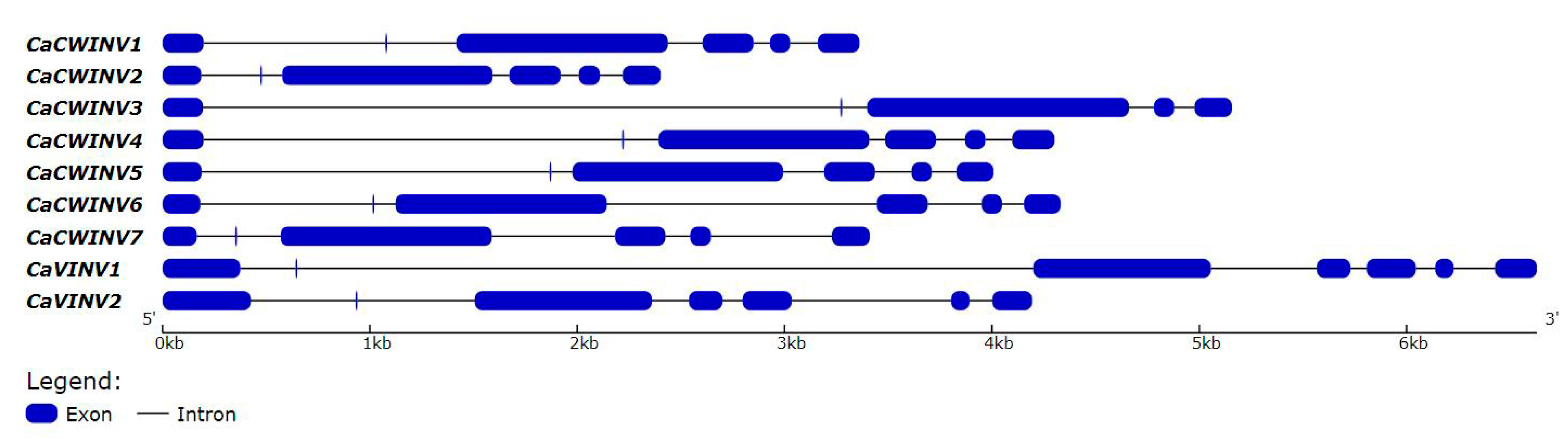
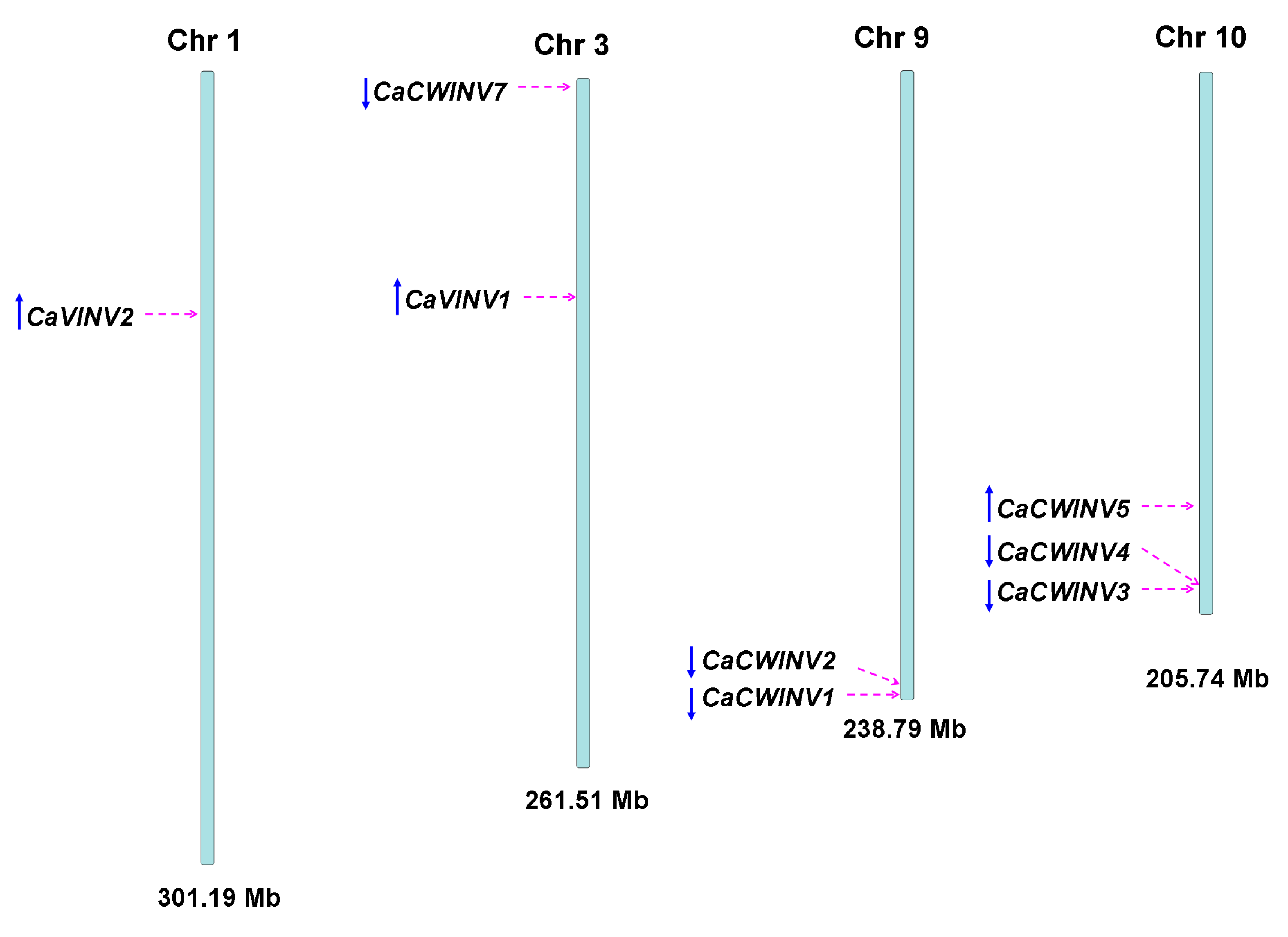
2.3. Structural Analysis and Chromosomal Distribution of CaAINV Family Genes
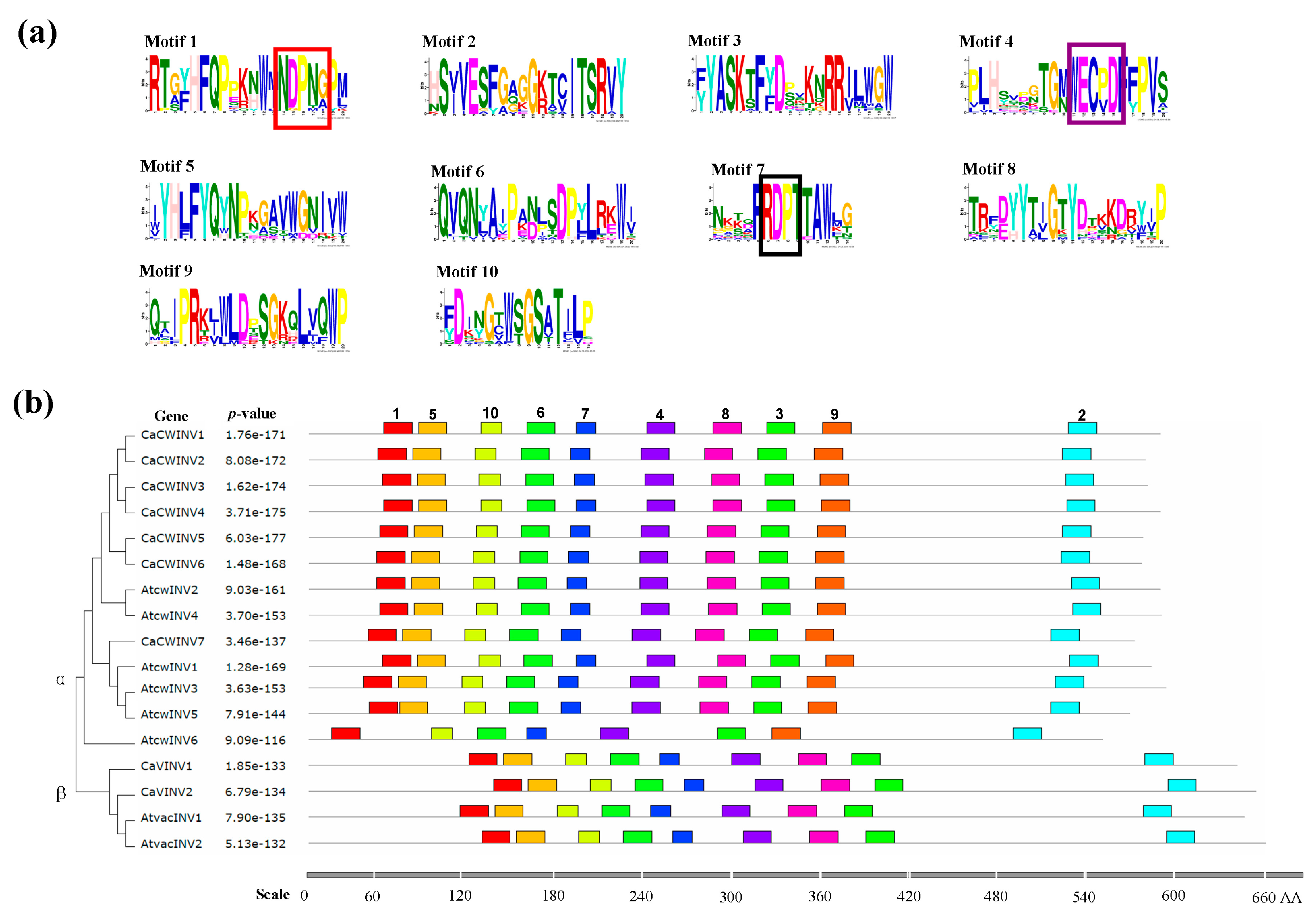
2.4. Motif Distribution in CaAINV Proteins
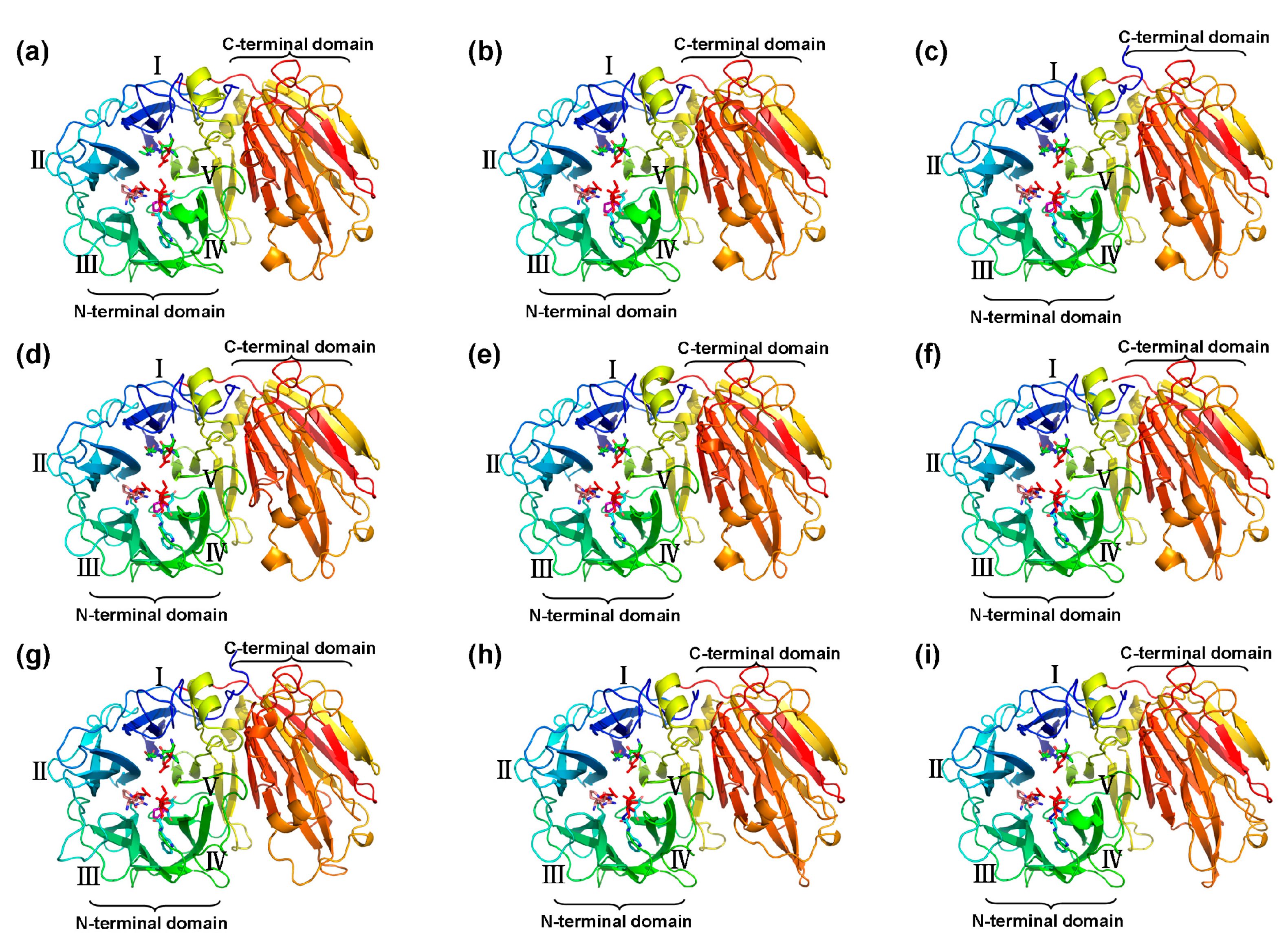
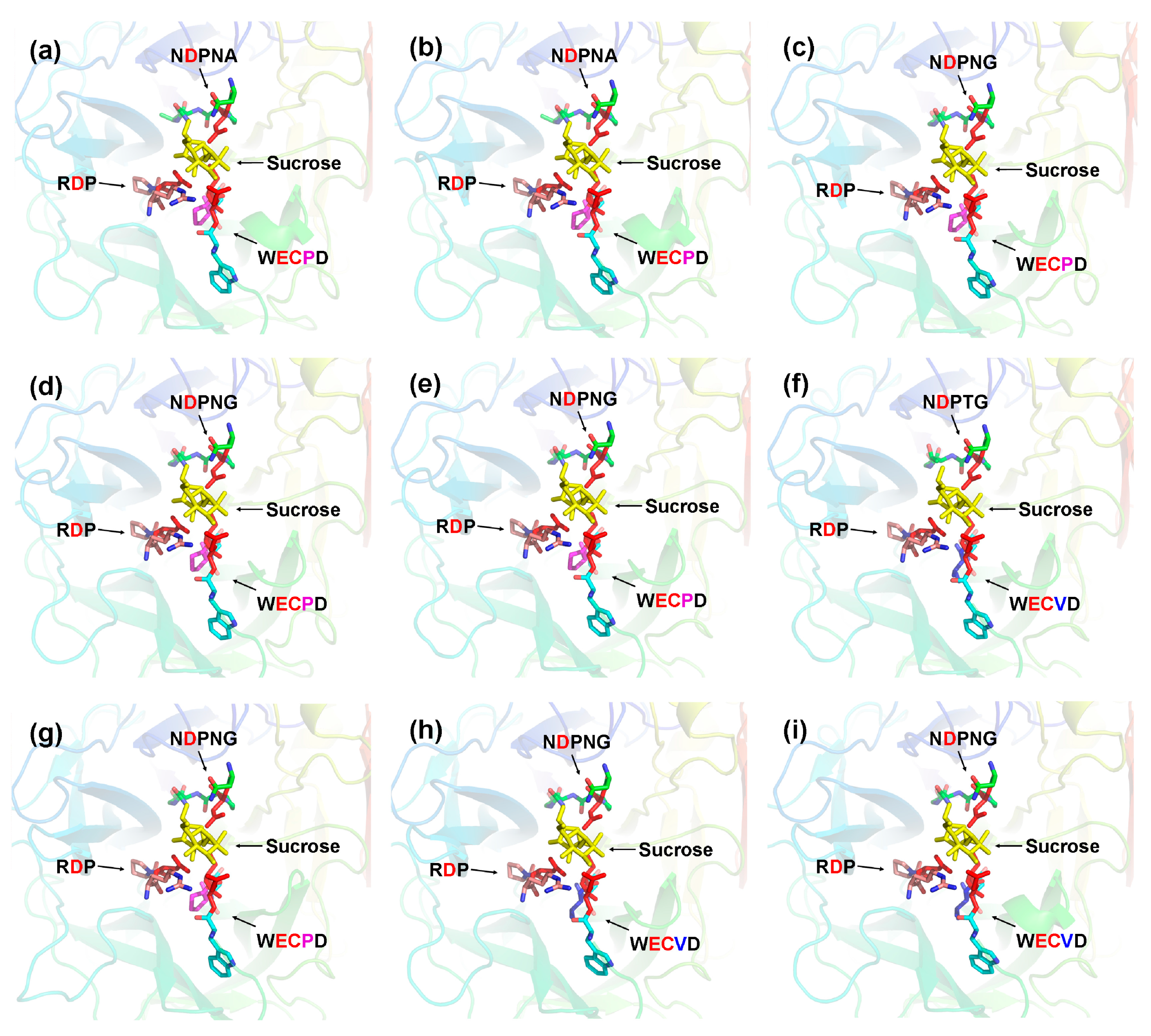
2.5. Three-Dimensional Structure of CaAINV Proteins
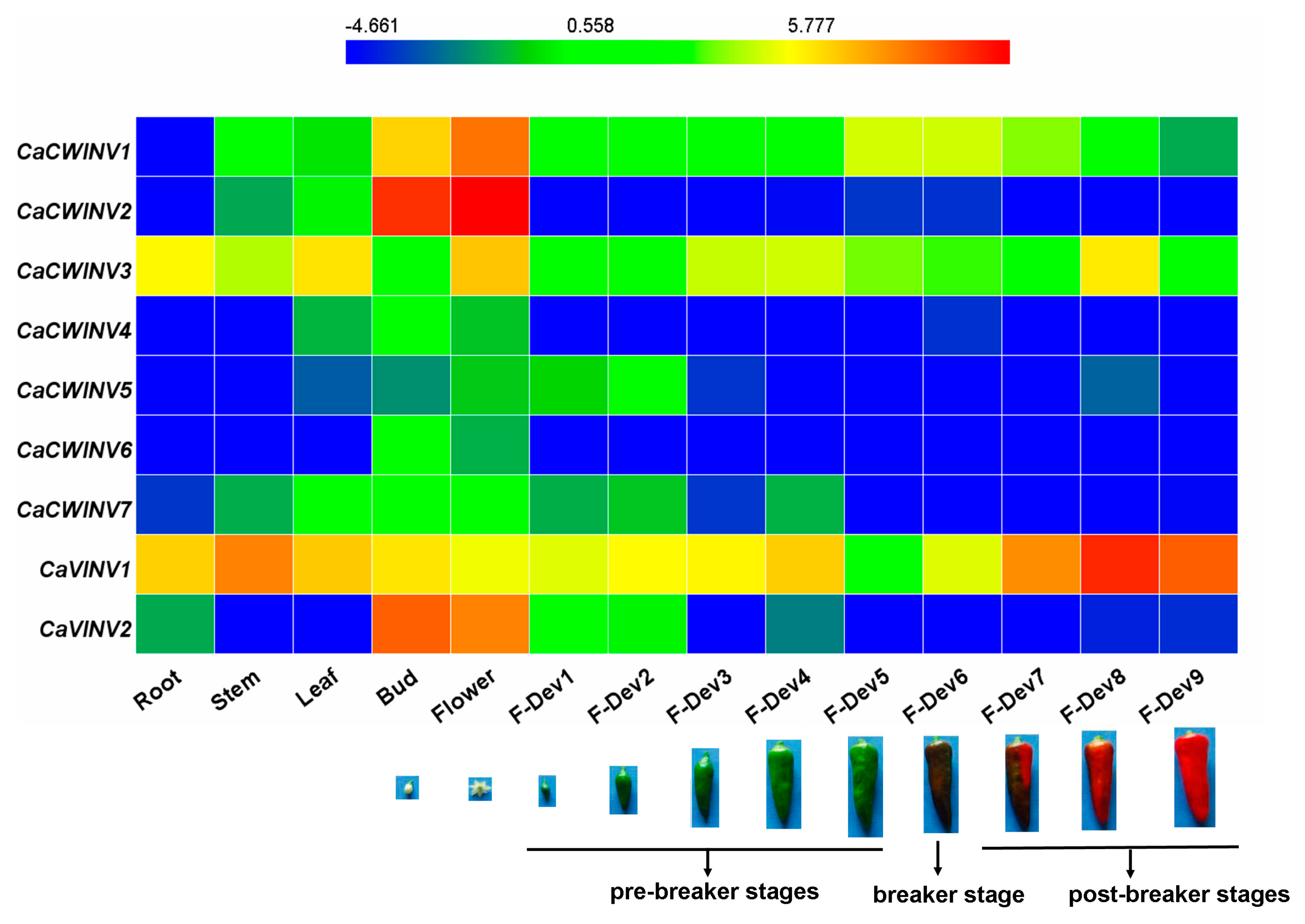
2.6. Expression Analysis of CaAINV Genes for Different Tissues and Developmental Stages
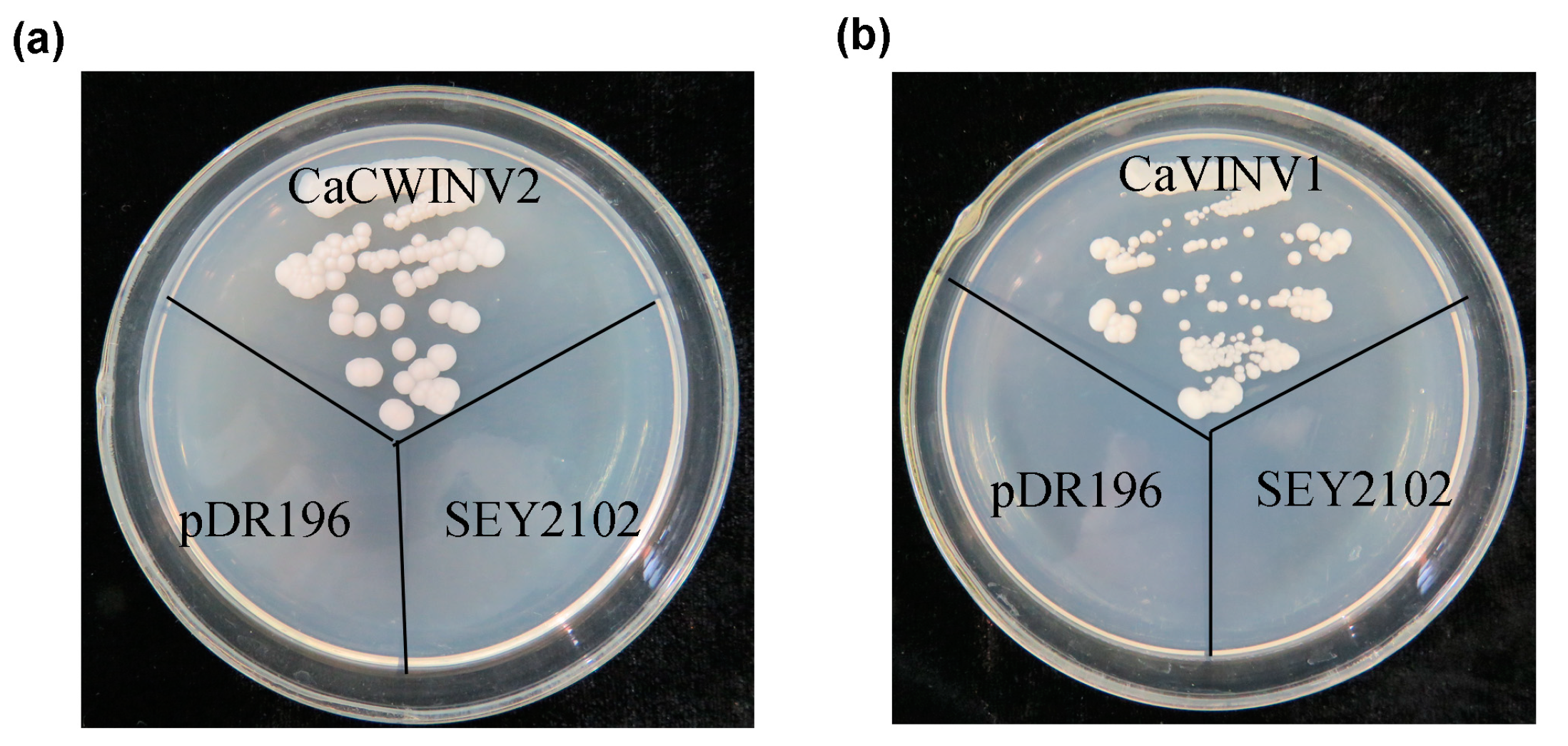
2.7. Yeast Complementation of CaCWINV-2 and CaVINV-1
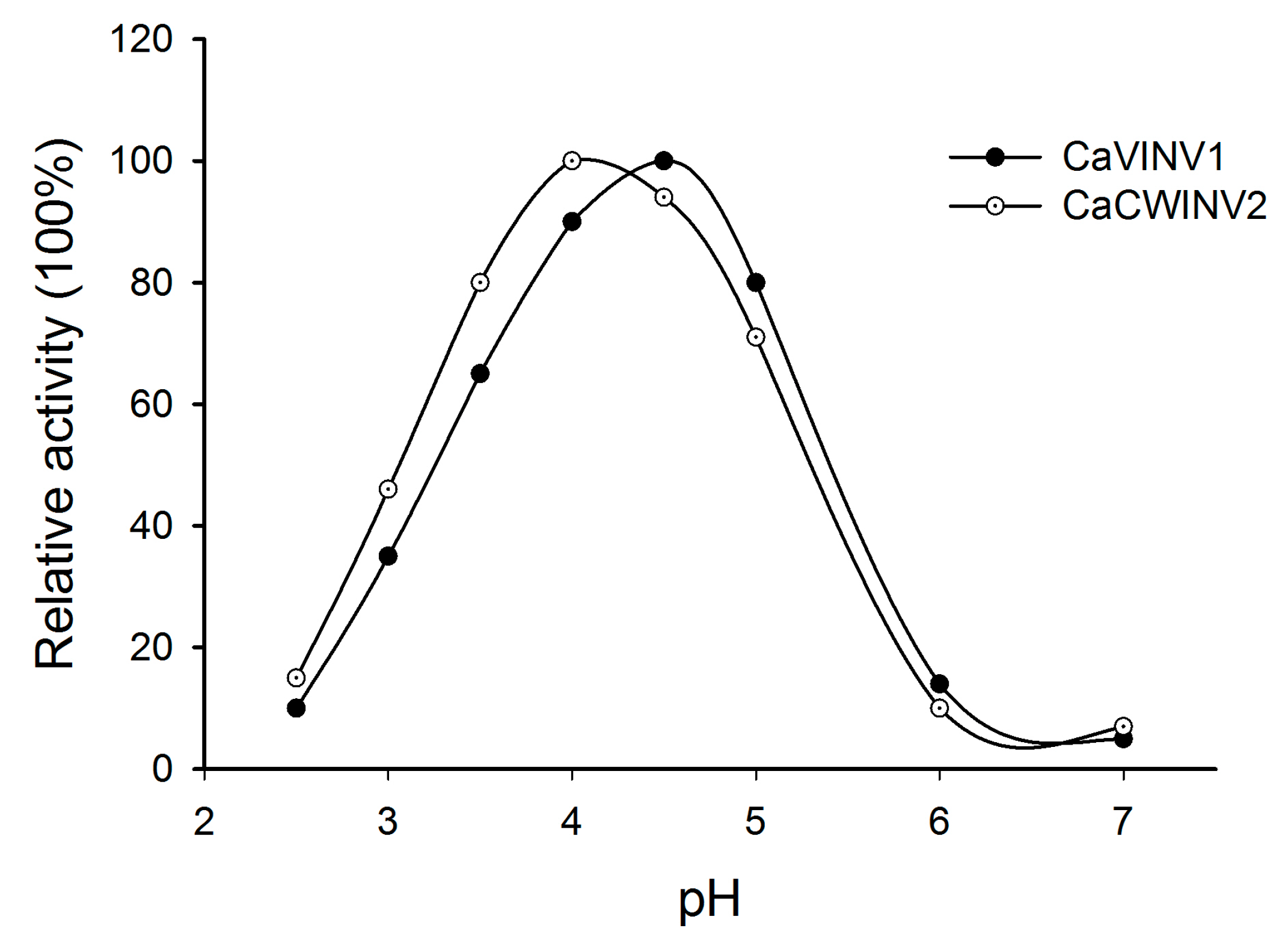
2.8. Optimum pH Determination for CaCWINV2 and CaVINV1
3. Discussion
3.1. Identification and Characterization of CaAINV Genes
3.2. Differential Expression and Enzymatic Activities of CaAINVs
4. Materials and Methods
4.1. Identification and Sequence Analysis of CaAINV Proteins in Pepper
4.2. Phylogenetic Analyses
4.3. Exon–Intron Structure Analysis and Chromosomal Mapping
4.4. Conserved Motif Analysis
4.5. Prediction of Three-Dimensional Structure of the CaAINV Proteins
4.6. Analysis of Expression Characteristics of CaAINV Genes
4.7. Yeast Complementation and Enzymatic Analysis of CaCWINV2 and CaVINV1
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Elwan, M.W.M.; El-Hamahmy, M.A.M. Improved productivity and quality associated with salicylic acid application in greenhouse pepper. Sci. Hortic. 2009, 122, 521–526. [Google Scholar] [CrossRef]
- Marcelis, L.F.M.; Heuvelink, E.; Hofman-Eijer, L.R.B.; Bakker, J.D.; Xue, L.B. Flower and fruit abortion in sweet pepper in relation to source and sink strength. J. Exp. Bot. 2004, 55, 2261–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aloni, B.; Karni, L.; Zaidman, Z.; Schaffer, A.A. Changes of Carbohydrates in Pepper (Capsicum annuum L.) Flowers in Relation to Their Abscission Under Different Shading Regimes. Ann. Bot. 1996, 78, 163–168. [Google Scholar] [CrossRef]
- Hubbard, N.L.; Pharr, D.M. Developmental changes in carbohydrate concentration and activaties of sucrose metabolizing enzymes in fruits of two Capsicum annuum L. genotypes. Plant Sci. 1992, 86, 33–39. [Google Scholar] [CrossRef]
- Aloni, B.; Peet, M.; Pharr, M.; Karni, L. The effect of high temperature and high atmospheric CO2 on carbohydrate changes in bell pepper (Capsicum annuum) pollen in relation to its germination. Physiol. Plant. 2001, 112, 505–512. [Google Scholar] [CrossRef]
- Fotopoulos, V. Plant Invertases: structure, function and regulation of a diverse enzyme family. J. Biol. Res. 2005, 4, 127–137. [Google Scholar]
- Bocock, P.; Morse, A.; Dervinis, C.; Davis, J. Evolution and diversity of invertase genes in Populus trichocarpa. Planta 2008, 227, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; van den Ende, W.; van Laere, A.; Cheng, S.; Bennett, J. Structure, evolution, and expression of the two invertase gene families of rice. J. Mol. Evol. 2005, 60, 615–634. [Google Scholar] [CrossRef]
- Xiang, L.; van den Ende, W. Trafficking of plant vacuolar invertases: From a membrane-anchored to a soluble status. understanding sorting information in their complex N-Terminal motifs. Plant Cell Physiol. 2013, 54, 1263–1277. [Google Scholar] [CrossRef]
- Chen, T.H.; Huang, Y.C.; Yang, C.S.; Yang, C.C.; Wang, A.Y.; Sung, H.Y. Insights into the catalytic properties of bamboo vacuolar invertase through mutational analysis of active site residues. Phytochemistry 2009, 70, 25–31. [Google Scholar] [CrossRef]
- Cho, J.I.; Lee, S.K.; Ko, S.; Kim, H.K.; Jun, S.H.; Lee, Y.H.; Bhoo, S.H.; Lee, K.W.; An, G.; Hahn, T.R. Molecular cloning and expression analysis of the cell-wall invertase gene family in rice (Oryza sativa L.). Plant Cell Rep. 2005, 24, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed]
- Tarchoun, N.; Rezgui, S.; Mougou, A. Sucrose synthase and acid invertase activities in relation to the floral structures abortion in pepper (Capsicum annuum L.) grown under low night temperature. J. Appl. Hortic. 2009, 11, 35–40. [Google Scholar]
- Yao, Y.; Wu, X.H.; Geng, M.T.; Li, R.M.; Liu, J.; Hu, X.W.; Guo, J.C. Cloning, 3D modeling and expression analysis of three vacuolar invertase genes from cassava (Manihot esculenta Crantz). Molecules 2014, 19, 6228–6245. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Geng, M.T.; Wu, X.H.; Liu, J.; Li, R.M.; Hu, X.W.; Guo, J.C. Genome-wide identification, 3D modeling, expression and enzymatic activity analysis of cell wall invertase gene family from cassava (Manihot esculenta Crantz). Int. J. Mol. Sci. 2014, 15, 7313–7331. [Google Scholar] [CrossRef] [PubMed]
- Fridman, E.; Zamir, D. Functional divergence of a syntenic invertase gene family in tomato, potato, and Arabidopsis. Plant Physiol. 2003, 131, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Verhaest, M.; Lammens, W.; Le Roy, K.; de Coninck, B.; de Ranter, C.J.; van Laere, A.; van den Ende, W.; Rabijns, A. X-ray diffraction structure of a cell-wall invertase from Arabidopsis thaliana. Acta Crystallogr. Sect. D 2006, 62, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Lammens, W.; Le Roy, K.; van Laere, A.; Rabijns, A.; van den Ende, W. Crystal structures of Arabidopsis thaliana cell-wall invertase mutants in complex with sucrose. J. Mol. Biol. 2008, 377, 378–385. [Google Scholar] [CrossRef]
- Klopotek, Y.; Franken, P.; Klaering, H.P.; Fischer, K.; Hause, B.; Hajirezaei, M.R.; Druege, U. A higher sink competitiveness of the rooting zone and invertases are involved in dark stimulation of adventitious root formation in Petunia hybrida cuttings. Plant Sci. 2016, 243, 10–22. [Google Scholar] [CrossRef]
- Lontom, W.; Kosittrakun, M.; Lingle, S.E. Relationship of acid invertase activities to sugar content in sugarcane internodes during ripening and after harvest. Thai J. Agri. Sci. 2008, 41, 143–151. [Google Scholar]
- Jin, Y.; Ni, D.-A.; Ruan, Y.L. Posttranslational elevation of cell wall invertase activity by silencing its inhibitor in tomato delays leaf senescence and increases seed weight and fruit hexose level. Plant Cell 2009, 21, 2072–2089. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Palmer, W.M.; Martin, A.P.; Wang, R.; Rainsford, F.; Jin, Y.; Patrick, J.W.; Yang, Y.; Ruan, Y.L. High invertase activity in tomato reproductive organs correlates with enhanced sucrose import into, and heat tolerance of, young fruit. J. Exp. Bot. 2012, 63, 1155–1166. [Google Scholar] [CrossRef]
- Palmer, W.M.; Ru, L.; Jin, Y.; Patrick, J.W.; Ruan, Y.L. Tomato ovary-to-fruit transition is characterized by a spatial shift of mrnas for cell wall invertase and its inhibitor with the encoded proteins localized to sieve elements. Mol. Plant 2015, 8, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Zhu, Z.; Wang, W.; Cai, J.; Chen, Y.; Li, L.; Tian, S. A tomato vacuolar invertase inhibitor mediates sucrose metabolism and influences fruit ripening. Plant Physiol. 2016, 172, 1596–1611. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Wu, L.; Yang, Y.; Zhou, G.; Ruan, Y.L. Evolution of sucrose metabolism: The dichotomy of invertases and beyond. Trends Plant Sci. 2018, 23, 163–177. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Han, L.; Huai, B.; Zheng, P.; Chang, Q.; Guan, T.; Li, D.; Huang, L.; Kang, Z. Down-regulation of a wheat alkaline/neutral invertase correlates with reduced host susceptibility to wheat stripe rust caused by Puccinia striiformis. J. Exp. Bot. 2015, 66, 7325–7338. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | ORF Length (bp) | Protein Length (aa) | Mw (kDa) | PI | Localization |
|---|---|---|---|---|---|---|
| CaCWINV1 | Capana09g002436 | 1773 | 590 | 67.41 | 9.23 | Cell wall |
| CaCWINV2 | Capana09g002437 | 1743 | 580 | 65.86 | 9.07 | Cell wall |
| CaCWINV3 | Capana10g002008 | 1746 | 581 | 65.78 | 9.60 | Cell wall |
| CaCWINV4 | Capana10g002007 | 1773 | 590 | 66.80 | 9.09 | Cell wall |
| CaCWINV5 | Capana10g001560 | 1737 | 578 | 66.16 | 9.16 | Cell wall |
| CaCWINV6 | Capana00g001535 | 1734 | 577 | 66.17 | 6.92 | Cell wall |
| CaCWINV7 | Capana03g000156 | 1719 | 572 | 64.36 | 6.52 | Cell wall |
| CaVINV1 | Capana03g002552 | 1932 | 643 | 70.94 | 5.97 | Vacuole |
| CaVINV2 | Capana01g000522 | 1971 | 656 | 73.29 | 5.51 | Vacuole |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, L.-B.; Qin, Y.-L.; Qi, Z.-Q.; Niu, Y.; Liu, Z.-J.; Liu, W.-X.; He, H.; Cao, Z.-M.; Yang, Y. Genome-Wide Analysis, Expression Profile, and Characterization of the Acid Invertase Gene Family in Pepper. Int. J. Mol. Sci. 2019, 20, 15. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20010015
Shen L-B, Qin Y-L, Qi Z-Q, Niu Y, Liu Z-J, Liu W-X, He H, Cao Z-M, Yang Y. Genome-Wide Analysis, Expression Profile, and Characterization of the Acid Invertase Gene Family in Pepper. International Journal of Molecular Sciences. 2019; 20(1):15. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20010015
Chicago/Turabian StyleShen, Long-Bin, Yu-Ling Qin, Zhi-Qiang Qi, Yu Niu, Zi-Ji Liu, Wei-Xia Liu, Huang He, Zhen-Mu Cao, and Yan Yang. 2019. "Genome-Wide Analysis, Expression Profile, and Characterization of the Acid Invertase Gene Family in Pepper" International Journal of Molecular Sciences 20, no. 1: 15. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20010015