Voluntary Activity Modulates Sugar-Induced Elastic Fiber Remodeling in the Alveolar Region of the Mouse Lung

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Body Weight and Blood Glucose Concentrations
2.2. Lung Mechanics
2.3. Lung Structure
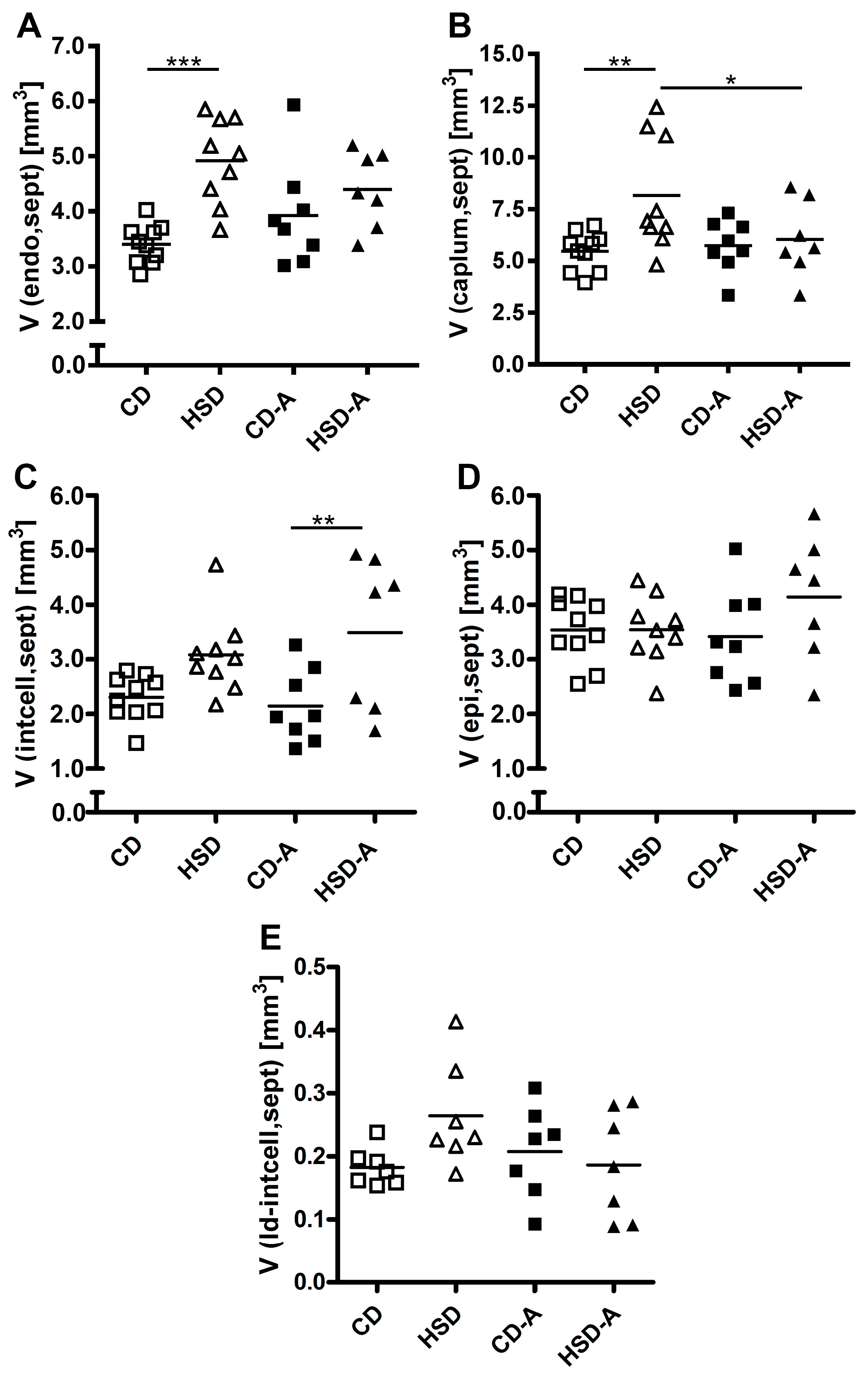
2.4. Parenchyma Composition
2.5. Septal Composition
2.6. Extracellular Matrix Composition
3. Discussion
4. Materials and Methods
4.1. Animals and Study Design
4.2. Lung Mechanics
4.3. Lung Fixation, Sampling, and Processing
4.4. Stereological Analysis—Light Microscopy
4.5. Stereological Analysis—Transmission Electron Microscopy
4.6. Scoring of Structural Elastic Fiber Appearance
4.7. Protein Islolation And Western Blot
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CD | Control diet |
| HSD | High sucrose diet |
| ECM | Extracellular matrix |
| MetS | Metabolic syndrome |
| COPD | Chronic obstructive pulmonary disease |
| CST | Static lung compliance |
| IC | Inspiratory capacity |
References
- Lustig, R.H.; Schmidt, L.A.; Brindis, C.D. The toxic truth about sugar. Nature 2012, 482, 27. [Google Scholar] [CrossRef] [PubMed]
- Lustig, R.H. Fructose: Metabolic, Hedonic, and Societal Parallels with Ethanol. J. Am. Diet. Assoc. 2010, 110, 1307–1321. [Google Scholar] [CrossRef]
- Malik, V.S.; Popkin, B.M.; Bray, G.A.; Després, J.-P.; Willett, W.C.; Hu, F.B. Sugar-Sweetened Beverages and Risk of Metabolic Syndrome and Type 2 Diabetes: A meta-analysis. Diabetes Care 2010, 33, 2477–2483. [Google Scholar] [CrossRef]
- Baffi, C.W.; Wood, L.; Winnica, D.; Strollo, P.J., Jr.; Gladwin, M.T.; Que, L.G.; Holguin, F. Metabolic Syndrome and the Lung. Chest 2016, 149, 1525–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, W.A.; Knuiman, M.; Kendall, P.; Grange, V.; Davis, T.M.E. Glycemic Exposure Is Associated with Reduced Pulmonary Function in Type 2 Diabetes: The Fremantle Diabetes Study. Diabetes Care 2004, 27, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Punjabi, N.M.; Beamer, B.A. Alterations in Glucose Disposal in Sleep-disordered Breathing. Am. J. Respir. Crit. Care Med. 2009, 179, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Cazzola, M.; Rogliani, P.; Calzetta, L.; Lauro, D.; Page, C.; Matera, M.G. Targeting Mechanisms Linking COPD to Type 2 Diabetes Mellitus. Trends Pharmacol. Sci. 2017, 38, 940–951. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.M.; Cutts, N.R.; Brody, J.S. Influence of maternal diabetes on basement membranes, type 2 cells, and capillaries in the developing rat lung. Dev. Biol. 1984, 104, 469–476. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Saltin, B. Exercise as medicine—Evidence for prescribing exercise as therapy in 26 different chronic diseases. Scand. J. Med. Sci. Sports 2015, 25, 1–72 doi 101111/sms12581. [Google Scholar] [CrossRef]
- Lemes, Í.R.; Turi-Lynch, B.C.; Cavero-Redondo, I.; Linares, S.N.; Monteiro, H.L. Aerobic training reduces blood pressure and waist circumference and increases HDL-c in metabolic syndrome: A systematic review and meta-analysis of randomized controlled trials. J. Am. Soc. Hypertens. 2018, 12, 580–588. [Google Scholar] [CrossRef]
- Salcedo, P.A.; Lindheimer, J.B.; Klein-Adams, J.C.; Sotolongo, A.M.; Falvo, M.J. Effects of Exercise Training on Pulmonary Function in Adults with Chronic Lung Disease: A Meta-Analysis of Randomized Controlled Trials. Arch. Phys. Med. Rehabil. 2018, 99, 2561.e7–2569.e7. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.M.W.; Pedersen, B.K. The anti-inflammatory effect of exercise. J. Appl. Physiol. 2005, 98, 1154–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toledo, A.C.; Magalhaes, R.M.; Hizume, D.C.; Vieira, R.P.; Biselli, P.J.C.; Moriya, H.T.; Mauad, T.; Lopes, F.D.T.Q.S.; Martins, M.A. Aerobic exercise attenuates pulmonary injury induced by exposure to cigarette smoke. Eur. Respir. J. 2012, 39, 254–264. [Google Scholar] [CrossRef]
- Schipke, J.; Vital, M.; Schnapper-Isl, A.; Pieper, D.H.; Mühlfeld, C. Spermidine and Voluntary Activity Exert Differential Effects on Sucrose- Compared with Fat-Induced Systemic Changes in Male Mice. J. Nutr. 2019, 149, 451–462. [Google Scholar] [CrossRef]
- Burgstaller, G.; Oehrle, B.; Gerckens, M.; White, E.S.; Schiller, H.B.; Eickelberg, O. The instructive extracellular matrix of the lung: Basic composition and alterations in chronic lung disease. Eur. Respir. J. 2017, 50. [Google Scholar] [CrossRef]
- Kielty, C.M.; Sherratt, M.J.; Shuttleworth, C.A. Elastic fibres. J. Cell Sci. 2002, 115, 2817–2828. [Google Scholar]
- Shamhart, P.E.; Luther, D.J.; Adapala, R.K.; Bryant, J.E.; Petersen, K.A.; Meszaros, J.G.; Thodeti, C.K. Hyperglycemia enhances function and differentiation of adult rat cardiac fibroblasts. Can. J. Physiol. Pharmacol. 2014, 92, 598–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, H.; Fricovsky, E.; Ihm, S.; Schimke, M.; Maya-Ramos, L.; Aroonsakool, N.; Ceballos, G.; Dillmann, W.; Villarreal, F.; Ramirez-Sanchez, I. Role for high-glucose-induced protein O-GlcNAcylation in stimulating cardiac fibroblast collagen synthesis. Am. J. Physiol. Cell Physiol. 2014, 306, C794–C804. [Google Scholar] [CrossRef] [Green Version]
- Sedgwick, B.; Riches, K.; Bageghni, S.A.; O’Regan, D.J.; Porter, K.E.; Turner, N.A. Investigating inherent functional differences between human cardiac fibroblasts cultured from nondiabetic and Type 2 diabetic donors. Cardiovasc. Pathol. 2014, 23, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, K.R.; Lord, C.K.; West, T.A.; Stewart, J.A., Jr. Cardiac fibroblast-dependent extracellular matrix accumulation is associated with diastolic stiffness in type 2 diabetes. PLoS ONE 2013, 8, e72080. [Google Scholar] [CrossRef]
- Gewolb, I.; Torday, J. High glucose inhibits maturation of the fetal lung in vitro. Morphometric analysis of lamellar bodies and fibroblast lipid inclusions. Lab. Investig. 1995, 73, 59–63. [Google Scholar] [PubMed]
- Goldstein, R.; Poliks, C.F.; Pilch, P.F.; Smith, B.D.; Fine, A. Stimulation of Collagen Formation by Insulin and Insulin-Like Growth Factor I in Cultures of Human Lung Fibroblasts. Endocrinology 1989, 124, 964–970. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Inui, N.; Hakamata, A.; Suzuki, Y.; Enomoto, N.; Fujisawa, T.; Nakamura, Y.; Watanabe, H.; Suda, T. Changes in pulmonary endothelial cell properties during bleomycin-induced pulmonary fibrosis. Respir. Res. 2018, 26, 127. [Google Scholar] [CrossRef] [PubMed]
- Shifren, A.; Mecham, R.P. The stumbling block in lung repair of emphysema: Elastic fiber assembly. Proc. Am. Thorac. Soc. 2006, 3, 428–433. [Google Scholar] [CrossRef]
- Burgess, J.K.; Mauad, T.; Tjin, G.; Karlsson, J.C.; Westergren-Thorsson, G. The extracellular matrix—The under-recognized element in lung disease? J. Pathol. 2016, 240, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Schuyler, M.R.; Niewoehner, D.E.; Inkley, S.R.; Kohn, R. Abnormal Lung Elasticity in Juvenile Diabetes Mellitus. Am. Rev. Respir. Dis. 1976, 113, 37–41. [Google Scholar] [CrossRef]
- Sandler, M.; Bunn, A.E.; Stewart, R.I. Cross-Section Study of Pulmonary Function in Patients with Insulin-dependent Diabetes Mellitus. Am. Rev. Respir. Dis. 1987, 135, 223–229. [Google Scholar] [CrossRef]
- Ehrlich, S.F.; Quesenberry, C.P., Jr.; Van Den Eeden, S.K.; Shan, J.; Ferrara, A. Patients diagnosed with diabetes are at increased risk for asthma, chronic obstructive pulmonary disease, pulmonary fibrosis, and pneumonia but not lung cancer. Diabetes Care 2010, 33, 55–60. [Google Scholar] [CrossRef]
- Pitocco, D.; Fuso, L.; Conte, E.G.; Zaccardi, F.; Condoluci, C.; Scavone, G.; Incalzi, R.A.; Ghirlanda, G. Thediabetic lung-a new target organ? Rev. Diabet. Stud. 2012, 9, 23–35. [Google Scholar] [CrossRef]
- Zheng, H.; Wu, J.; Jin, Z.; Yan, L.-J. Potential Biochemical Mechanisms of Lung Injury in Diabetes. Aging Dis. 2017, 8, 7–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kida, K.; Utsuyama, M.; Takizawa, T.; Thurlbeck, W.M. Changes in Lung Morphologic Features and Elasticity Caused by Streptozotocin-induced Diabetes Mellitus in Growing Rats. Am. Rev. Respir. Dis. 1983, 128, 125–131. [Google Scholar] [CrossRef]
- Ofulue, A.F.; Kida, K.; Thurlbeck, W.M. Experimental Diabetes and the Lung: I. Changes in Growth, Morphometry, and Biochemistry. Am. Rev. Respir. Dis. 1988, 137, 162–166. [Google Scholar] [CrossRef]
- Ofulue, A.F.; Thurlbeck, W.M. Experimental Diabetes and the Lung: II. In Vivo Connective Tissue Metabolism. Am. Rev. Respir. Dis. 1988, 138, 284–289. [Google Scholar] [CrossRef]
- World Health Organization. Fact Sheet: Diabetes. 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 12 May 2019).
- Lee, Y.-S.; Eun, H.S.; Kim, S.Y.; Jeong, J.-M.; Seo, W.; Byun, J.-S.; Jeong, W.-I.; Yi, H.-S. Hepatic immunophenotyping for streptozotocin-induced hyperglycemia in mice. Sci. Rep. 2016, 6, 30656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colberg, S.R.; Sigal, R.J.; Fernhall, B.; Regensteiner, J.G.; Blissmer, B.J.; Rubin, R.R.; Chasan-Taber, L.; Albright, A.L.; Braun, B.; Medicine, A.C.; et al. Exercise and type 2 diabetes: The American College of Sports Medicine and the American Diabetes Association: Joint position statement. Diabetes Care 2010, 33, e147–e167. [Google Scholar] [CrossRef] [PubMed]
- Jelleyman, C.; Yates, T.; O’Donovan, G.; Gray, L.J.; King, J.A.; Khunti, K.; Davies, M.J. The effects of high-intensity interval training on glucose regulation and insulin resistance: A meta-analysis. Obes. Rev. 2015, 16, 942–961. [Google Scholar] [CrossRef] [PubMed]
- Rullman, E.; Norrbom, J.; Strömberg, A.; Wågsäter, D.; Rundqvist, H.; Haas, T.; Gustafsson, T. Endurance exercise activates matrix metalloproteinases in human skeletal muscle. J. Appl. Physiol. 2009, 106, 804–812. [Google Scholar] [CrossRef] [Green Version]
- Koskinen, S.O.A.; Heinemeier, K.M.; Olesen, J.L.; Langberg, H.; Kjaer, M. Physical exercise can influence local levels of matrix metalloproteinases and their inhibitors in tendon-related connective tissue. J. Appl. Physiol. 2004, 96, 861–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadoglou, N.P.E.; Vrabas, I.S.; Sailer, N.; Kapelouzou, A.; Fotiadis, G.; Noussios, G.; Karayannacos, P.E.; Angelopoulou, N. Exercise ameliorates serum MMP-9 and TIMP-2 levels in patients with type 2 diabetes. Diabetes Metab. 2010, 36, 144–151. [Google Scholar] [CrossRef]
- Lopez-Rodriguez, E.; Boden, C.; Echaide, M.; Perez-Gil, J.; Kolb, M.; Gauldie, J.; Maus, U.A.; Ochs, M.; Knudsen, L. Surfactant dysfunction during overexpression of TGF-β1 precedes profibrotic lung remodeling in vivo. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L1260–L1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudsen, L.; Atochina-Vasserman, E.N.; Massa, C.B.; Birkelbach, B.; Guo, C.-J.; Scott, P.; Haenni, B.; Beers, M.F.; Ochs, M.; Gow, A.J. The role of inducible nitric oxide synthase for interstitial remodeling of alveolar septa in surfactant protein D-deficient mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2015, 309, L959–L969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherle, W. A simple method for volumetry of organs in quantitative stereology. Mikroskopie 1970, 26, 57–60. [Google Scholar]
- Ochs, M.; Mühlfeld, C. Quantitative microscopy of the lung: A problem-based approach. Part 1: Basic principles of lung stereology. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 305, L15–L22. [Google Scholar] [CrossRef]
- Schneider, J.P.; Ochs, M. Alterations of mouse lung tissue dimensions during processing for morphometry: A comparison of methods. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 306, L341–L350. [Google Scholar] [CrossRef] [PubMed]
- Tschanz, S.A.; Burri, P.H.; Weibel, E.R. A simple tool for stereological assessment of digital images: The STEPanizer. J. Microsc. 2011, 243, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Curran-Everett, D.; Benos, D.J. Guidelines for reporting statistics in journals published by the American Physiological Society. Am. J. Physiol. Metab. 2004, 287, E189–E191. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Gel Concentration | Blocking Conditions | Primary Antibody, Dilution, Incubation Conditions | Secondary Antibody, Dilution, Incubation Conditions |
|---|---|---|---|---|
| Target Proteins | ||||
| Fibrillin-1 | 5% | drying and equilibration in 3% BSA for 10 min | Anti-Fibrillin 1 antibody | Peroxidase-AffiniPure F(ab’)2 Fragment Goat Anti-Rabbit IgG |
| (ab231094; Abcam, Cambridge, Great Britain) | (111-036-045, Dianova, Hamburg, Germany) | |||
| 1:1000 | 1:20,000 | |||
| overnight, 4 °C | 1 h, room temperature | |||
| Elastin | 10% | drying and equilibration in 3% BSA for 10 min | Anti-Elastin antibody | Peroxidase-AffiniPure F(ab’)2 Fragment Goat Anti-Rabbit IgG |
| (ab217356, Abcam) | (111-036-045, Dianova) | |||
| 1:2000 | 1:20,000 | |||
| overnight, 4 °C | 1 h, room temperature | |||
| Collagen I | 10% | 1 h 3% BSA/TBS | COL1A1 (3G3) | m-IgGκ BP-HRP |
| (sc-293182, Santa Cruz Biotechnology, Dallas, TX, USA) | (sc-516102, Santa Cruz Biotechnology) | |||
| 1:500 | 1:20,000 | |||
| overnight, 4 °C | 1 h, room temperature | |||
| Collagen III | 8% | 1 h 3% BSA/TBS | COL3A1 (B-10) | m-IgGκ BP-HRP |
| (sc-271249, Santa Cruz Biotechnology) | (sc-516102, Santa Cruz Biotechnology) | |||
| 1:500 | 1:20,000 | |||
| overnight, 4 °C | 1 h, room temperature | |||
| Loading Controls | ||||
| α-Actin | α-Actin Antibody (G-12) | Peroxidase-AffiniPure F(ab’)2 Fragment Goat Anti-Rabbit IgG | ||
| (sc-130619, Santa Cruz Biotechnology) | (111-036-045, Dianova) | |||
| 1:2000 | 1:20,000 | |||
| 1 h, room temperature | 1 h, room temperature | |||
| Vinculin | Vinculin antibody (7F9) | m-IgGκ BP-HRP | ||
| (sc-73614, Santa Cruz Biotechnology) | (sc-516102, Santa Cruz Biotechnology) | |||
| 1:2000 | 1:20,000 | |||
| 1 h, room temperature | 1 h, room temperature | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hollenbach, J.; Lopez-Rodriguez, E.; Mühlfeld, C.; Schipke, J. Voluntary Activity Modulates Sugar-Induced Elastic Fiber Remodeling in the Alveolar Region of the Mouse Lung. Int. J. Mol. Sci. 2019, 20, 2438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102438
Hollenbach J, Lopez-Rodriguez E, Mühlfeld C, Schipke J. Voluntary Activity Modulates Sugar-Induced Elastic Fiber Remodeling in the Alveolar Region of the Mouse Lung. International Journal of Molecular Sciences. 2019; 20(10):2438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102438
Chicago/Turabian StyleHollenbach, Julia, Elena Lopez-Rodriguez, Christian Mühlfeld, and Julia Schipke. 2019. "Voluntary Activity Modulates Sugar-Induced Elastic Fiber Remodeling in the Alveolar Region of the Mouse Lung" International Journal of Molecular Sciences 20, no. 10: 2438. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102438