6-SFT, a Protein from Leymus mollis, Positively Regulates Salinity Tolerance and Enhances Fructan Levels in Arabidopsis thaliana

 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Sequence Analysis of 6-SFT
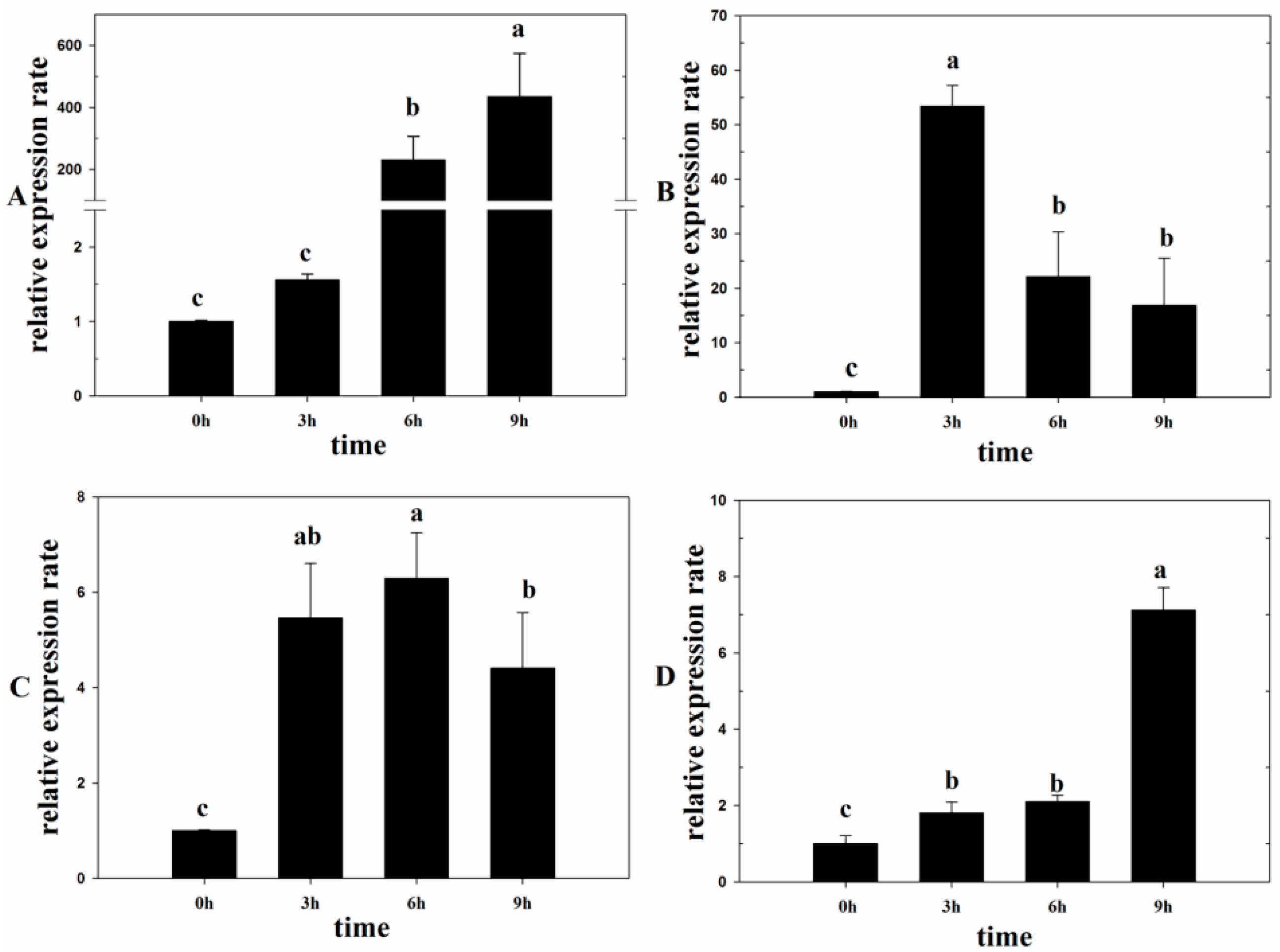
2.2. Lm-6-SFT Was Upregulated by the Exogenous ABA, SA, Light, and NaCl Treatments
2.3. Overexpression of 6-SFT Enhanced the Salt Tolerance of Transgenic Arabidopsis
2.4. Analysis of the Promoter of Lm-6-SFT
2.5. GUS Analysis of the Lm-6-SFT Expression Pattern
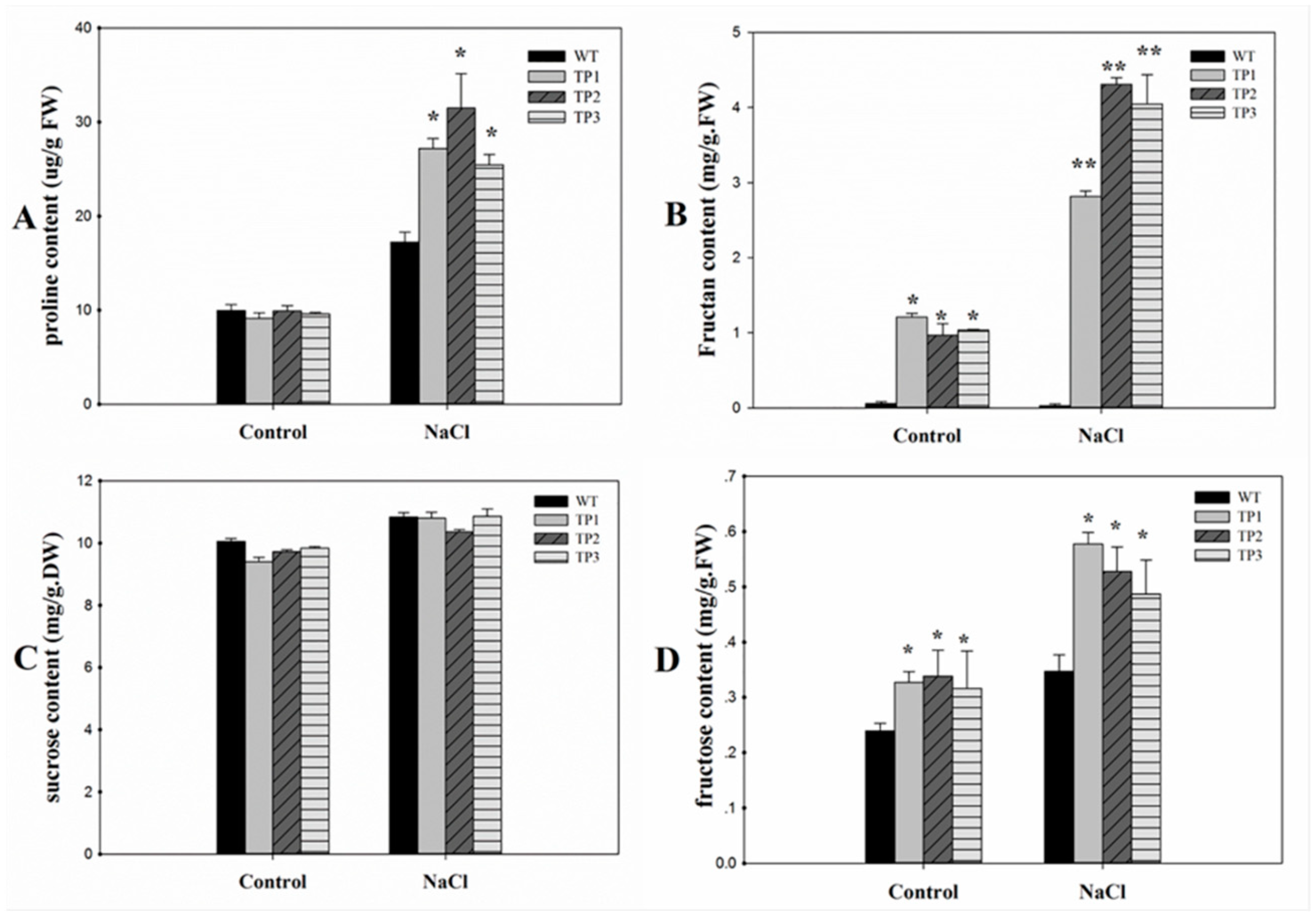
2.6. Expression of 6-SFT Increased the Accumulation of Proline and the Capacity for Osmotic Adjustment in Transgenic Arabidopsis Plants
2.7. Overexpression of Lm-6-SFT Upregulated Stress-Related Genes in Transgenic Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Plant Growth and Treatments
4.2. Isolation of Lm-6-SFT
4.3. Isolation and Analysis of the Promoter of Lm-6-SFT
4.4. Expression Pattern of Lm-6-SFT under Treatment with ABA, SA, Light, and NaCl
4.5. Vector Construction and Arabidopsis Transformation
4.6. GUS Histochemical Staining Analysis
4.7. Characterization of TPs and Tolerance of Salt Stress in T3 Arabidopsis TPs
4.8. Measurement of Proline, Fructan, Sucrose, and Fructose Levels in T3 Arabidopsis TPs and WT Plants
4.9. qPCR Analysis of T3-Generation TPs and WT Plants
4.10. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 6-SFT | Sucrose fructan 6-fructosyltransferase |
| TPs | transgenic plants |
| WT | wild type |
| RACE | rapid amplification of cDNA ends |
| GUS | β-glucuronidase |
| qPCR | quantitative PCR |
References
- Vinocur, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: Achievements and limitations. Curr. Opin. Biotechnol. 2005, 16, 123–132. [Google Scholar] [CrossRef]
- Hendry, G.A.F. Evolutionary Origins and Natural Functions of Fructans—A Climatological, Biogeographic and Mechanistic Appraisal. New Phytol. 1993, 123, 3–14. [Google Scholar] [CrossRef]
- Puebla, A.F.; Battaglia, M.E.; Salerno, G.L.; Pontis, H.G. Sucrose-sucrose fructosyl transferase activity: A direct and rapid colorimetric procedure for the assay of plant extracts. Plant Physiol. Biochem. 1999, 37, 699–702. [Google Scholar] [CrossRef]
- Xue, G.-P.; McIntyre, C.L.; Jenkins, C.L.D.; Glassop, D.; van Herwaarden, A.F.; Shorter, R. Molecular Dissection of Variation in Carbohydrate Metabolism Related to Water-Soluble Carbohydrate Accumulation in Stems of Wheat. Plant Physiol. 2008, 146, 441–454. [Google Scholar] [CrossRef]
- Lidgett, A. Isolation and characterisation of a sucrose: Sucrose 1-fructosyltransferase gene from perennial ryegrass (Lolium perenne). J. Plant Physiol. 2003, 160, 1385–1391. [Google Scholar]
- Xue, G.P.; McIntyre, C.L.; Glassop, D.; Shorter, R. Use of expression analysis to dissect alterations in carbohydrate metabolism in wheat leaves during drought stress. Plant Mol. Biol. 2008, 67, 197–214. [Google Scholar] [CrossRef]
- Meng, G.; Fütterer, K. Structural framework of fructosyl transfer in Bacillus subtilis levansucrase. Nat. Struct. Biol. 2003, 10, 935. [Google Scholar] [CrossRef]
- Trevisan, F.; Chu, E.P.; Gaspar, M.; Carvalho, M.A.M. In vitro culture and fructan production by Vernonia herbacea (Asteraceae). Acta Physiol. Plant. 2014, 36, 2299–2307. [Google Scholar] [CrossRef]
- Trevisan, F.; Oliveira, V.F.; Carvalho, M.A.; Gaspar, M. Effects of different carbohydrate sources on fructan metabolism in plants of Chrysolaena obovata grown in vitro. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Van den Ende, W.; Yoshida, M.; Clerens, S.; Vergauwen, R.; Kawakami, A. Cloning, characterization and functional analysis of novel 6-kestose exohydrolases (6-KEHs) from wheat (Triticum aestivum). New Phytol. 2005, 166, 917–932. [Google Scholar] [CrossRef] [PubMed]
- Wim, V.D.E.; Marlies, C.; Stefan, C.; Rudy, V.; Katrien, L.R.; Willem, L.; André, V.L. Unexpected presence of graminan- and levan-type fructans in the evergreen frost-hardy eudicot Pachysandra terminalis (Buxaceae): Purification, cloning, and functional analysis of a 6-SST/6-SFT enzyme. Plant Physiol. 2011, 155, 603–614. [Google Scholar]
- Lidgett, A.; Jennings, K.; Johnson, X.; Guthridge, K.; Jones, E.; Spangenberg, G. Isolation and characterisation of a fructosyltransferase gene from perennial ryegrass ( Lolium perenne ). J. Plant Physiol. 2002, 159, 1037–1043. [Google Scholar] [CrossRef]
- Iii, D.P.L.; Hincha, D.K.; Heyer, A.G. Fructan and its relationship to abiotic stress tolerance in plants. Cell. Mol. Life Sci. 2009, 66, 2007–2023. [Google Scholar] [CrossRef] [Green Version]
- Garcia, P.M.; Asega, A.F.; Silva, E.A.; Carvalho, M.A. Effect of drought and re-watering on fructan metabolism in Vernonia herbacea (Vell.) Rusby. Plant Physiol. Biochem. 2011, 49, 664. [Google Scholar] [CrossRef] [PubMed]
- Bereswill, S.; Jock, S.; Aldridge, P.; Janse, J.D.; Geider, K. Molecular characterization of naturalErwinia amylovorastrains deficient in levan synthesis. Physiol. Mol. Plant Pathol. 1997, 51, 215–225. [Google Scholar] [CrossRef]
- Ende, W.V.D.; Michiels, A.; Wonterghem, D.V.; Vergauwen, R.; Laere, A.V. Cloning, developmental, and tissue-specific expression of sucrose:sucrose 1-fructosyl transferase from Taraxacum officinale. Fructan localization in roots. Plant Physiol. 2000, 123, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Cairns, A.J. Fructan biosynthesis in transgenic plants. J. Exp. Bot. 2003, 54, 549–567. [Google Scholar] [CrossRef]
- Tamura, K.; Sanada, Y.; Tase, K.; Kawakami, A.; Yoshida, M.; Yamada, T. Comparative study of transgenic Brachypodium distachyon expressing sucrose:fructan 6-fructosyltransferases from wheat and timothy grass with different enzymatic properties. Planta 2014, 239, 783–792. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.Z.; Chatterton, N.J. Fructan biosynthesis and fructosyltransferase evolution: Expression of the 6SFT (sucrose: Fructan 6-fructosyltransferase) gene in crested wheatgrass (Agropyron cristatum). J. Plant Physiol. 2001, 158, 1203–1213. [Google Scholar] [CrossRef]
- Xue, G.P.; Mcintyre, C.L.; Rattey, A.R.; Herwaarden, A.F.V.; Shorter, R. Use of dry matter content as a rapid and low-cost estimate for ranking genotypic differences in water-soluble carbohydrate concentrations in the stem and leaf sheath of Triticum aestivum. Crop Past. Sci. 2009, 60, 51–59. [Google Scholar] [CrossRef]
- Kawakami, A.; Yoshida, M. Molecular Characterization of Sucrose:Sucrose 1-Fructosyltransferase and Sucrose:Fructan 6-Fructosyltransferase Associated with Fructan Accumulation in Winter Wheat during Cold Hardening. Biosci. Biotechnol. Biochem. 2002, 66, 2297–2305. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, W.; Dell, B.; Vergauwen, R.; Zhang, X.; Mayer, J.; Van den Ende, W. Wheat genotypic variation in dynamic fluxes of WSC components in different stem segments under drought during grain filling. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Iii, D.P.L.; Knievel, D.P.; Gildow, F.E. Fructan synthesis in oat: I. Oligomer accumulation in stems during cold hardening and their in vitro synthesis in a crude enzyme extract. New Phytol. 2010, 127, 27–36. [Google Scholar] [CrossRef]
- Hochstrasser, U.; Lüscher, M.; De Virgilio, C.; Boller, T.; Wiemken, A. Expression of a functional barley sucrose-fructan 6-fructosyltransferase in the methylotrophic yeast Pichia pastoris. FEBS Lett. 1998, 440, 356–360. [Google Scholar] [CrossRef]
- Hisano, H.; Kanazawa, A.; Kawakami, A.; Yoshida, M.; Shimamoto, Y.; Yamada, T. Transgenic perennial ryegrass plants expressing wheat fructosyltransferase genes accumulate increased amounts of fructan and acquire increased tolerance on a cellular level to freezing. Plant Sci. 2004, 167, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Bie, X.; Wang, K.; She, M.; Du, L.; Zhang, S.; Li, J.; Gao, X.; Lin, Z.; Ye, X. Combinational transformation of three wheat genes encoding fructan biosynthesis enzymes confers increased fructan content and tolerance to abiotic stresses in tobacco. Plant Cell Rep. 2012, 31, 2229–2238. [Google Scholar] [CrossRef]
- He, X.; Chen, Z.; Wang, J.; Li, W.; Zhao, J.; Wu, J.; Wang, Z.; Chen, X. A sucrose:fructan-6-fructosyltransferase (6-SFT) gene from Psathyrostachys huashanica confers abiotic stress tolerance in tobacco. Gene 2015, 570, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Honermeier, B. Effect of water stress on proline accumulation of genetically modified potatoes (Solanum tuberosum L.) generating fructans. J. Plant Physiol. 2006, 163, 392. [Google Scholar] [CrossRef]
- Roover, J.D.; Laere, A.V.; Ende, W.V.D. Effect of defoliation on fructan pattern and fructan metabolizing enzymes in young chicory plants (Cichorium intybus). Physiol. Plant. 1999, 106, 158–163. [Google Scholar] [CrossRef]
- Lothier, J.; Lasseur, B.; Roy, K.L.; Laere, A.V.; Prud’Homme, M.P.; Barre, P.; Ende, W.V.D.; Morvan-Bertrand, A. Cloning, gene mapping, and functional analysis of a fructan 1-exohydrolase (1-FEH) from Lolium perenne implicated in fructan synthesis rather than in fructan mobilization. J. Exp. Bot. 2007, 58, 1969–1983. [Google Scholar] [CrossRef]
- Fusari, C.M.; del Viso, F.; Puebla, A.F.; Hopp, H.E.; Heinz, R.A.; Pontis, H.G.; Casabuono, A.C.; Couto, A.S. Molecular Characterization of a Putative Sucrose:Fructan 6-Fructosyltransferase (6-SFT) of the Cold-Resistant Patagonian Grass Bromus pictus Associated With Fructan Accumulation Under Low Temperatures. Plant Cell Physiol. 2009, 50, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Abeynayake, S.W.; Etzerodt, T.P.; Jonavičienė, K.; Byrne, S.; Asp, T.; Boelt, B. Fructan metabolism and changes in fructan composition during cold acclimation in perennial ryegrass. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigui, A.P.; Gaspar, M.; Oliveira, V.F.; Purgatto, E.; Carvalho, M.A.M.d. Endogenous hormone concentrations correlate with fructan metabolism throughout the phenological cycle in Chrysolaena obovata. Ann. Bot. 2015, 115, 1163–1175. [Google Scholar] [CrossRef] [PubMed]
- Valluru, R. Fructan and hormone connections. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, G.P.; Kooiker, M.; Drenth, J.; McIntyre, C.L. TaMYB13 is a transcriptional activator of fructosyltransferase genes involved in β-2, 6-linked fructan synthesis in wheat. Plant J. 2011, 68, 857–870. [Google Scholar] [CrossRef]
- Nagaraj, V.J.; Riedl, R.; Boller, T.; Wiemken, A.; Meyer, A.D. Light and sugar regulation of the barley sucrose: Fructan 6-fructosyltransferase promoter. J. Plant Physiol. 2001, 158, 1601–1607. [Google Scholar] [CrossRef]
- Lu, C.; Koroleva, O.A.; Farrar, J.F.; Gallagher, J.; Pollock, C.J.; Tomos, A.D. Rubisco Small Subunit, Chlorophylla/b-Binding Protein and Sucrose:Fructan-6-Fructosyl Transferase Gene Expression and Sugar Status in Single Barley Leaf Cells in Situ. Cell Type Specificity and Induction by Light. Plant Physiol. 2002, 130, 1335–1348. [Google Scholar] [CrossRef] [Green Version]
- Kishii, M.; Wang, R.R.; Tsujimoto, H. Characteristics and behaviour of the chromosomes of Leymus mollis and L. racemosus (Triticeae, Poaceae) during mitosis and meiosis. Chromosome Res. 2003, 11, 741–748. [Google Scholar] [CrossRef]
- Zhao, J.X.; Du, W.L.; Wu, J.; Cheng, X.N.; Gao, Y.; Pang, Y.H.; Chen, X.H.; Liu, S.H.; Yang, Q.H.; Fu, J. Development and identification of a wheat- Leymus mollis multiple alien substitution line. Euphytica 2013, 190, 45–52. [Google Scholar] [CrossRef]
- Habora, M.E.E.; Eltayeb, A.E.; Tsujimoto, H.; Tanaka, K. Identification of osmotic stress-responsive genes from Leymus mollis, a wild relative of wheat (Triticum aestivum L.). Breed. Sci. 2012, 62, 78–86. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Ritsema, T.; Hernández, L.; Verhaar, A.; Altenbach, D.; Boller, T.; Wiemken, A.; Smeekens, S. Developing fructan-synthesizing capability in a plant invertase via mutations in the sucrose-binding box. Plant J. 2006, 48, 228–237. [Google Scholar] [CrossRef]
- Lee, K.H.; Piao, H.L.; Kim, H.-Y.; Choi, S.M.; Jiang, F.; Hartung, W.; Hwang, I.; Kwak, J.M.; Lee, I.-J.; Hwang, I. Activation of Glucosidase via Stress-Induced Polymerization Rapidly Increases Active Pools of Abscisic Acid. Cell 2006, 126, 1109–1120. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, R.K.; Gupta, A.; Soni, D.; Garg, R.; Pathre, U.V.; Nath, P.; Sane, A.P. Ectopic expression of a tomato DREB gene affects several ABA processes and influences plant growth and root architecture in an age-dependent manner. J. Plant Physiol. 2017, 214, 97–107. [Google Scholar] [CrossRef]
- Diedhiou, C.; Gaudet, D.; Liang, Y.; Sun, J.; Lu, Z.-X.; Eudes, F.; Laroche, A. Carbohydrate profiling in seeds and seedlings of transgenic triticale modified in the expression of sucrose:sucrose-1-fructosyltransferase (1-SST) and sucrose:fructan-6-fructosyltransferase (6-SFT). J. Biosci. Bioeng. 2012, 114, 371–378. [Google Scholar] [CrossRef]
- Jiang, F.; Hartung, W. Long-distance signalling of abscisic acid (ABA): The factors regulating the intensity of the ABA signal. J. Exp. Bot. 2007, 59, 37–43. [Google Scholar] [CrossRef]
- Suárez-González, E.M.; López, M.G.; Délano-Frier, J.P.; Gómez-Leyva, J.F. Expression of the 1-SST and 1-FFT genes and consequent fructan accumulation in Agave tequilana and A. inaequidens is differentially induced by diverse (a) biotic-stress related elicitors. J. Plant Physiol. 2014, 171, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.-Z.; Chatterton, N.J.; Larson, S.R. Expression of sucrose: Fructan 6-fructosyltransferase (6-SFT) and myo-inositol 1-phosphate synthase (MIPS) genes in barley (Hordeum vulgare) leaves. J. Plant Physiol. 2001, 158, 635–643. [Google Scholar] [CrossRef]
- He, X.L.; Wang, J.W.; Li, W.X.; Chen, Z.Z.; Wu, J.; Zhao, J.X.; Su, J.N.; Wang, Z.H.; Chen, X.H. An intronless sucrose:fructan-6-fructosyltransferase (6-SFT) gene from Dasypyrum villosum enhances abiotic tolerance in tobacco. Biol. Plant. 2017, 61, 235–245. [Google Scholar] [CrossRef]
- Van den Ende, W.; De Coninck, B.; Van Laere, A. Plant fructan exohydrolases: A role in signaling and defense? Trends Plant Sci. 2004, 9, 523–528. [Google Scholar] [CrossRef]
- De Coninck, B.; Le Roy, K.; Francis, I.; Clerens, S.; Vergauwen, R.; Halliday, A.M.; Smith, S.M.; Van Laere, A.; Van Den Ende, W. Arabidopsis AtcwINV3 and 6 are not invertases but are fructan exohydrolases (FEHs) with different substrate specificities. Plant Cell Environ. 2005, 28, 432–443. [Google Scholar] [CrossRef]
- Van den Ende, W.; El-Esawe, S.K. Sucrose signaling pathways leading to fructan and anthocyanin accumulation: A dual function in abiotic and biotic stress responses? Environ. Exp. Bot. 2014, 108, 4–13. [Google Scholar] [CrossRef]
- Ebskamp, M.J.; Im, V.D.M.; Spronk, B.A.; Weisbeek, P.J.; Smeekens, S.C. Accumulation of fructose polymers in transgenic tobacco. Bio/technology 1995, 12, 272–275. [Google Scholar] [CrossRef]
- Van den Ende, W. Multifunctional fructans and raffinose family oligosaccharides. Front. Plant Sci. 2013, 4, 247. [Google Scholar] [Green Version]
- Vogt, L.; Ramasamy, U.; Meyer, D.; Pullens, G.; Venema, K.; Faas, M.M.; Schols, H.A.; de Vos, P. Immune modulation by different types of β2→ 1-fructans is toll-like receptor dependent. PLoS ONE 2013, 8, e68367. [Google Scholar] [CrossRef]
- Ross, A.R.S.; Ambrose, S.J.; Cutler, A.J.; Allan Feurtado, J.; Kermode, A.R.; Nelson, K.; Zhou, R.; Abrams, S.R. Determination of endogenous and supplied deuterated abscisic acid in plant tissues by high-performance liquid chromatography-electrospray ionization tandem mass spectrometry with multiple reaction monitoring. Anal. Biochem. 2004, 329, 324–333. [Google Scholar] [CrossRef]
- Denoux, C.; Galletti, R.; Mammarella, N.; Gopalan, S.; Werck, D.; De Lorenzo, G.; Ferrari, S.; Ausubel, F.M.; Dewdney, J. Activation of Defense Response Pathways by OGs and Flg22 Elicitors in Arabidopsis Seedlings. Mol. Plant 2008, 1, 423–445. [Google Scholar] [CrossRef] [Green Version]
- Ariga, H.; Katori, T.; Yoshihara, R.; Hase, Y.; Nozawa, S.; Narumi, I.; Iuchi, S.; Kobayashi, M.; Tezuka, K.; Sakata, Y.; et al. Arabidopsis sos1 mutant in a salt-tolerant accession revealed an importance of salt acclimation ability in plant salt tolerance. Plant Signal. Behav. 2013, 8, e24779. [Google Scholar] [CrossRef]
- Hamam, A.M.; Britto, D.T.; Flam-Shepherd, R.; Kronzucker, H.J. Measurement of Differential Na+ Efflux from Apical and Bulk Root Zones of Intact Barley and Arabidopsis Plants. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Ullah, A.; Dutta, D.; Fliegel, L. Expression and characterization of the SOS1 Arabidopsis salt tolerance protein. Mol. Cell. Biochem. 2016, 415, 133–143. [Google Scholar] [CrossRef]
- Li, X.-Y.; Liu, X.; Yao, Y.; Li, Y.-H.; Liu, S.; He, C.-Y.; Li, J.-M.; Lin, Y.-Y.; Li, L. Overexpression of Arachis hypogaea AREB1 gene enhances drought tolerance by modulating ROS scavenging and maintaining endogenous ABA content. Int. J. Mol. Sci. 2013, 14, 12827–12842. [Google Scholar] [CrossRef] [PubMed]
- Hiroaki, F.; Verslues, P.E.; Jian-Kang, Z. Identification of two protein kinases required for abscisic acid regulation of seed germination, root growth, and gene expression in Arabidopsis. Plant Cell 2007, 19, 485–494. [Google Scholar]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Lakra, N.; Nutan, K.K.; Das, P.; Anwar, K.; Singla-Pareek, S.L.; Pareek, A. A nuclear-localized histone-gene binding protein from rice (OsHBP1b) functions in salinity and drought stress tolerance by maintaining chlorophyll content and improving the antioxidant machinery. J. Plant Physiol. 2015, 176, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant. Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Cheikh, N.; Brenner, M.L. Regulation of key enzymes of sucrose biosynthesis in soybean leaves: Effect of dark and light conditions and role of gibberellins and abscisic acid. Plant Physiol. 1992, 100, 1230–1237. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequences |
|---|---|
| PBI121R | GACCACCCGGGGATCCTCTAGATTGAAGAAACAAGTCATCGTCC |
| PBI121F | AGAGAACACGGGGGACTCTAGAATGGGGTCACACGGCAAG |
| DP6-SFT-F | AAYGARATGYTNCARTGG |
| DP6-SFT-R | NCCRTCNARNACNGGNAC |
| GSOP3′-LL3 | TGAGGCTGATGTGGGCTAT |
| GSIP3′-LL3 | CCTCGTCCTCGCTGCTGGTA |
| GSOP5′-LL3 | GGTGACCAGATGACGGGATT |
| GSIP5′-LL3 | TGCACTGGACCTCAACAGC |
| SFTF | ATGGGGTCACACGGCAAGCCA |
| SFTR | TCATTGAAGAAACAAGTCATC |
| AtactinF | GCACCCTGTTCTTCTTACCGAG |
| AtactinR | AGTAAGGTCACGTCCAGCAAGG |
| RD29AF | TAATCGGAAGACACGACAGG |
| RD29AR | GATGTTTAGGAAAGTAAAGGCTAG |
| p5CS1F | AGCTTGATGACGTTATCGATCT |
| p5CS1R | AGATTCCATCAGCATGACCTAG |
| RD22F | CATGAGTCTCCGGGAGGAAGTG |
| RD22R | CGGCTGGGGTAAAGAAGTTGTC |
| QSFTF | TGTTGAAGACGAGCATGGAC |
| QSFTR | TGGACGCATAAAACTTACCCC |
| Lm-actinF | CTCCCTCACAACAACAACCGC |
| Lm-actinR | TACCAGGAACTTCCATACCAAC |
| sp1 | CGACAACCACCACCACCATGG |
| sp2 | GCCACCTCGTACAGCCGGTCC |
| sp3 | CTTGTACGCGTACGGTAGTGG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; He, X.; Hao, D.; Wu, J.; Zhao, J.; Yang, Q.; Chen, X. 6-SFT, a Protein from Leymus mollis, Positively Regulates Salinity Tolerance and Enhances Fructan Levels in Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 2691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20112691
Li M, He X, Hao D, Wu J, Zhao J, Yang Q, Chen X. 6-SFT, a Protein from Leymus mollis, Positively Regulates Salinity Tolerance and Enhances Fructan Levels in Arabidopsis thaliana. International Journal of Molecular Sciences. 2019; 20(11):2691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20112691
Chicago/Turabian StyleLi, Mao, Xiaolan He, Dongdong Hao, Jun Wu, Jixin Zhao, Qunhui Yang, and Xinhong Chen. 2019. "6-SFT, a Protein from Leymus mollis, Positively Regulates Salinity Tolerance and Enhances Fructan Levels in Arabidopsis thaliana" International Journal of Molecular Sciences 20, no. 11: 2691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20112691