Metabolomics Studies on Cytoplasmic Male Sterility during Flower Bud Development in Soybean

Abstract
:1. Introduction
2. Results
2.1. Metabolite Profiling of Soybean Flower Buds
2.2. Principal Component Analysis (PCA) and Orthogonal Projections to Latent Structures-Discriminant Analysis (OPLS-DA) for the NJCMS5A Group Versus NJCMS5B Group
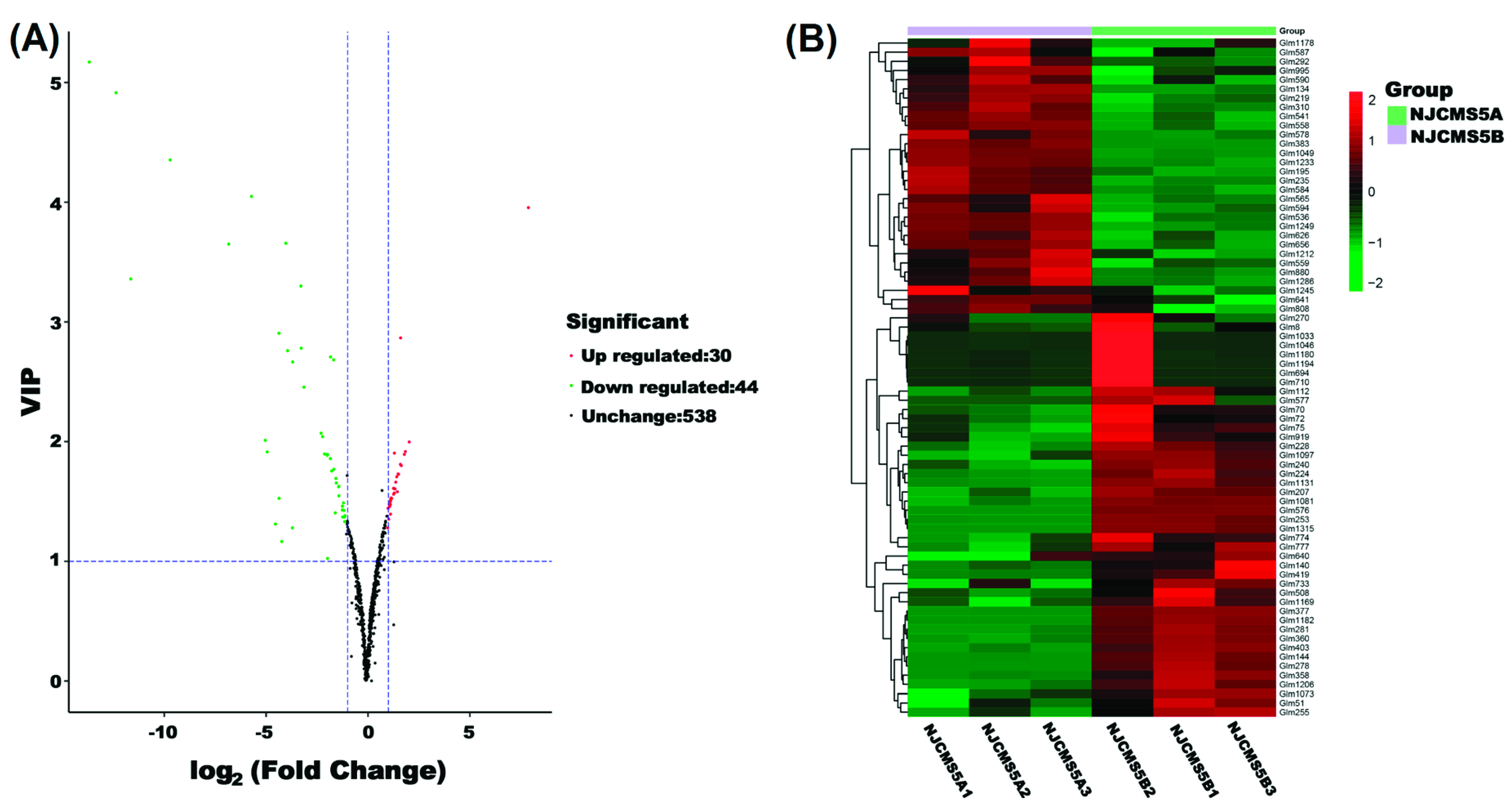
2.3. Metabolic Changes in Flower Bud of Soybean CMS Line
2.4. Altered Functional Pathways in Soybean CMS Line
2.5. Activities of ROS Scavenging Enzymes
3. Discussion
3.1. Differential Metabolites between Soybean CMS Line and Its Maintainer
3.2. Disorder of Metabolites in Non-enZymatic ROS Scavenging System of Soybean CMS Line
3.3. Altered Enzymatic ROS Scavenging System Exist in Flower Buds of Soybean CMS Line
3.4. Proposed Model for ROS Burst and ROS Scavenging System Deficiency in Soybean CMS-N8855
4. Materials and Methods
4.1. Plant Materials
4.2. Plant Traits Investigation
4.3. Sample Preparation and Extraction
4.4. UPLC-MS/MS Conditions
4.5. Qualitative and Quantitative Determination of Metabolite
4.6. Statistical Analysis
4.7. Kyoto Encyclopaedia of Genes and Genomes (KEGG) Annotation and Metabolic Pathway Analysis of Differential Metabolites
4.8. Determination of ROS Scavenging Enzyme Activities and Flavonoid Content
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Wise, R.P.; Pring, D.R. Nuclear-mediated mitochondrial gene regulation and male fertility in higher plants: light at the end of the tunnel. Proc. Natl. Acad. Sci. USA 2002, 99, 10240–10242. [Google Scholar] [CrossRef] [PubMed]
- Gutierres, S.; Combettes, B.; De Paepe, R.; Mirande, M.; Lelandais, C.; Vedel, F.; Chétrit, P. In the Nicotiana sylvestris CMSII mutant, a recombination-mediated change 5′ to the first exon of the mitochondrial nad1 gene is associated with lack of the NADH: ubiquinone oxidoreductase (complex I) NAD1 subunit. Eur. J. Biochem. 1999, 261, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Touzet, P.; Meyer, E.H. Cytoplasmic male sterility and mitochondrial metabolism in plants. Mitochondrion 2014, 19, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.T.; Liu, Y.G. Male sterility and fertility restoration in crops. Annu. Rev. Plant Biol. 2014, 65, 579–606. [Google Scholar] [CrossRef] [PubMed]
- Green, D.; Reed, J. Mitochondria and apoptosis. Science 1998, 281, 1309–1312. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.D.; Zhang, X.Q.; Zhu, Y.G.; Zhu, W.; Xie, H.Y.; Wang, X.D. Metabolism of reactive oxygen species in cotton cytoplasmic male sterility and its restoration. Plant Cell Rep. 2007, 26, 1627–1634. [Google Scholar] [CrossRef] [PubMed]
- Lamb, C.J.; Dixon, R.A. The oxidative burst in plant disease resistance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 251–275. [Google Scholar] [CrossRef] [PubMed]
- Jabs, T. Reactive oxygen intermediates as mediators of programmed cell death in plants and animals. Biochem. Pharmacol. 1999, 57, 231–245. [Google Scholar] [CrossRef]
- Papini, A.; Mosti, S.; Brighigna, L. Programmed-cell-death events during tapetum development of angiosperms. Protoplasma 1999, 207, 213–221. [Google Scholar] [CrossRef]
- Kawanabe, T.; Ariizumi, T.; Kawai-Yamada, M.; Uchimiya, H.; Toriyama, K. Abolition of the tapetum suicide program ruins microsporogenesis. Plant Cell Physiol. 2006, 47, 784–787. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Dubey, R.S. Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul. 2005, 46, 209–221. [Google Scholar] [CrossRef]
- Srivastava, S.; Dubey, R.S. Manganese-excess induces oxidative stress, lowers the pool of antioxidants and elevates activities of key antioxidative enzymes in rice seedlings. Plant Growth Regul. 2011, 64, 1–16. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef]
- Li, S.Q.; Wan, C.X.; Kong, J.; Zhang, Z.J.; Li, Y.S.; Zhu, Y.G. Programmed cell death during microgenesis in a Hong-lian CMS line of rice is correlated with oxidative stress in mitochondria. Funct. Plant Biol. 2004, 31, 369–376. [Google Scholar] [CrossRef]
- Wan, C.X.; Li, S.Q.; Wen, L.; Kong, J.; Wang, K.; Zhu, Y.G. Damage of oxidative stress on mitochondria during microspores development in Honglian CMS line of rice. Plant Cell Rep. 2007, 26, 373–382. [Google Scholar] [CrossRef]
- Deng, M.H.; Wen, J.F.; Huo, J.L.; Zhu, H.S.; Dai, X.Z.; Zhang, Z.Q.; Zhou, H.; Zou, X.X. Relationship of metabolism of reactive oxygen species with cytoplasmic male sterility in pepper (Capsicum annuum L.). Sci. Hortic. 2012, 134, 232–236. [Google Scholar] [CrossRef]
- Ba, Q.S.; Zhang, G.S.; Wang, J.S.; Che, H.X.; Liu, H.Z.; Niu, N.; Ma, S.C.; Wang, J.W. Relationship between metabolism of reactive oxygen species and chemically induced male sterility in wheat (Triticum aestivum L.). Can. J. Plant Sci. 2013, 93, 675–681. [Google Scholar] [CrossRef]
- Qu, G.R.; Quan, S.; Mondol, P.; Xu, J.; Zhang, D.B.; Shi, J.X. Comparative metabolomic analysis of wild type and mads3 mutant rice anthers. J. Integr. Plant Biol. 2014, 56, 849–863. [Google Scholar] [CrossRef]
- Rudneva, I.I. Antioxidant system of black sea animals in early development. Comp. Biochem. Physiol. C 1999, 112, 265–271. [Google Scholar] [CrossRef]
- Bassard, J.E.; Ullmann, P.; Bernier, F.; Werck-Reichhart, D. Phenolamides: Bridging polyamines to the phenolic metabolism. Phytochemistry 2010, 71, 1808–1824. [Google Scholar] [CrossRef]
- Aceroa, N.; Gradillasb, A.; Beltrana, M.; Garcíab, A.; Mingarro, D.M. Comparison of phenolic compounds profile and antioxidant properties of different sweet cherry (Prunus avium L.) varieties. Food Chem. 2019, 279, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Edreva, A.M.; Velikova, V.B.; Tsonev, T.D. Phenylamides in plants. Russian J. Plant Physiol. 2007, 54, 287–301. [Google Scholar] [CrossRef]
- Paupière, M.J.; Müller, F.; Li, H.; Rieu, I.; Tikunov, Y.M.; Visser, R.G.F.; Bovy, A.G. Untargeted metabolomic analysis of tomato pollen development and heat stress response. Plant Reprod. 2017, 30, 81–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, N.N.; Liu, F.; Wang, P.D.; Yan, X.H.; Gao, H.F.; Zeng, X.H.; Wu, G. Overexpression of BraLTP2, a lipid transfer protein of Brassica napus, results in increased trichome density and altered concentration of secondary metabolites. Int. J. Mol. Sci. 2018, 19, 1733. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.X.; Zhao, T.J.; Yang, S.P.; Gai, J.Y. Development of a cytoplasmic male-sterile line NJCMS4A for hybrid soybean production. Plant Breed. 2017, 136, 516–525. [Google Scholar] [CrossRef]
- Li., J.J.; Han, S.H.; Ding, X.L.; He, T.T.; Dai, J.Y.; Yang, S.P.; Gai, J.Y. Comparative transcriptome analysis between the cytoplasmic male sterile line NJCMS1A and its maintainer NJCMS1B in soybean (Glycine max (L.) Merr.). PLoS ONE 2015, 10, e0126771. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.L.; Song, Y.L.; Guo, J.L.; Wang, J.W.; Zhang, L.L.; Niu, N.; Ma, S.C.; Zhang, G.S.; Zhao, H.Y. Physiological and metabolome changes during anther development in wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2018, 132, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Li, S.M.; Su, X.Q.; Abdullah, M.; Sun, Y.M.; Li, G.H.; Cheng, X.; Lin, Y.; Cai, Y.P.; Jin, Q. Effects of different pollens on primary metabolism and lignin biosynthesis in pear. Int. J. Mol. Sci. 2018, 19, 2273. [Google Scholar] [CrossRef] [PubMed]
- Shishova, M.; Puzanskiy, R.; Gavrilova, O.; Kurbanniazov, S.; Demchenko, K.; Yemelyanov, V.; Pendinen, G.; Shavarda, A.; Gavrilenko, T. Metabolic alterations in male-sterile potato as compared to male-fertile. Metabolites 2019, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Wang, J. Transcriptomics and Metabolimics Studies on Cytoplasmic Male Sterility in Cotton. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2013; pp. 67–114. [Google Scholar]
- Li, J.J.; Ding, X.L.; Han, S.H.; Zhang, H.; Yang, L.S.; Yang, S.P.; Gai, J.Y. Differential proteomics analysis to identify proteins and pathways associated with male sterility of soybean using iTRAQ-based strategy. J. Proteomics 2016, 138, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Mo, Y.; Nagel, C.; Taylor, L.P. Biochemical complementation of chalcone synthase mutants defines a role for flavonols in functional pollen. Proc. Natl. Acad. Sci. USA 1992, 89, 7213–7217. [Google Scholar] [CrossRef] [PubMed]
- Ylstra, B.; Touraev, A.; Benito Moreno, R.M.; Stöger, E.; van Tunen, A.J.; Vicente, O.; Mol, J.N.M.; Heberle-Bors, E. Flavonols stimulate development, germination and tube growth of tobacco pollen. Plant Physiol. 1992, 100, 902–907. [Google Scholar] [CrossRef]
- Wiermann, R. Stage specific phenylpropanoid metabolism during pollen development. In Regulation of Secondary Product and Plant Hormone Metabolism; Luckner, M., Schreiber, L., Eds.; Pergamon Press: Oxford, UK, 1979; pp. 231–239. [Google Scholar] [CrossRef]
- Schijlen, E.G.; de Vos, C.H.; Martens, S.; Jonker, H.H.; Rosin, F.M.; Molthoff, J.W.; Tikunov, Y.M.; Angenent, G.C.; van Tunen, A.J.; Bovy, A.G. RNA interference silencing of chalcone synthase, the first step in the flavonoid biosynthesis pathway, leads to parthenocarpic tomato fruits. Plant Physiol. 2007, 144, 1520–1530. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.P.; Jorgensen, R. Conditional male fertility in chalcone synthase-deficient Petunia. J. Hered. 1992, 83, 11–17. [Google Scholar] [CrossRef]
- Van der Meer, I.M.; Stam, M.E.; Van Tunen, A.J.; Mol, J.N.; Stuitje, A.R. Antisense inhibition of flavonoid biosynthesis in Petunia anthers results in male sterility. Plant Cell 1992, 4, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Coe, E.H.; Mccormick, S.M.; Modena, S.A. White pollen in maize. J. Hered. 1981, 72, 318–320. [Google Scholar] [CrossRef]
- Strack, D.; Eilert, U.; Wray, V.; Wolff, J.; Jaggy, H. Tricoumaroylspermidine in flowers of rosaceae. Phytochemistry 1990, 29, 2893–2896. [Google Scholar] [CrossRef]
- Tarenghi, E.; Martin-Tanguy, J. Polyamides, floral induction and floral development of strawberry (Fragaria ananassa Duch.). Plant Growth Regul. 1995, 17, 157–165. [Google Scholar] [CrossRef]
- Martin-Tanguy, J. The occurrence and possible function of hydroxycinnamoyl acid amines in plants. Plant Growth Regul. 1985, 3, 381–399. [Google Scholar] [CrossRef]
- Kakkar, R.K.; Rai, V.K. Plant polyamines in flowering and fruit ripening. Phytochemistry 1993, 33, 1281–1288. [Google Scholar] [CrossRef]
- Martin-Tanguy, J.; Perdrizet, E.; Prevost, J.; Martin, C. Hydroxycinnamic acid amides in fertile and cytoplasmic male sterile lines of maize. Phytochemistry 1982, 21, 1939–1945. [Google Scholar] [CrossRef]
- Cabanne, F.; Dalebroux, M.A.; Martin-Tanguy, J.; Martin, C. Hydroxycinnamic acid amides and ripening to flower of Nicotiana tabaccum var. Xanthi n.c. Physiol. Plant 1981, 53, 399–404. [Google Scholar] [CrossRef]
- Arora, A.; Sairam, R.K.; Srivastava, G.C. Oxidative stress and antioxidative system in plants. Curr. Sci. 2002, 82, 1227–1238. [Google Scholar]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Dewey, R.E.; Timothy, D.H.; Levings, C.S. A mitochondrial protein associated with cytoplasmic male sterility in the T cytoplasm of maize. Proc. Natl. Acad. Sci. USA 1987, 84, 5374–5378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, G.G.; Formanova, N.; Jin, H.; Wargachuk, R.; Dendy, C.; Patil, P.; Laforest, M.; Zhang, J.; Cheung, W.Y.; Landry, B.S. The radish Rfo restorer gene of Ogura cytoplasmic male sterility encodes a protein with multiple pentatricopeptide repeats. Plant J. 2003, 35, 262–272. [Google Scholar] [CrossRef]
- Linke, B.; Börner, T. Mitochondrial effects on flower and pollen development. Mitochondrion 2005, 5, 389–402. [Google Scholar] [CrossRef]
- Connettm, M.B.; Hanson, M.R. Differential mitochondrial electron transport through the cyanide sensitive and cyanide-insensitive pathways in isonuclear lines of cytoplasmic male sterile, male fertile, and restored Petunia. Plant Physiol. 1990, 93, 1634–1640. [Google Scholar] [CrossRef]
- Bergman, P.; Edqvist, J.; Farbos, I.; Glimelius, K. Male sterile tobacco displays abnormal mitochondrial atp1 transcript accumulation and reduced floral ATP/ADP ratio. Plant Mol. Biol. 2000, 42, 531–544. [Google Scholar] [CrossRef]
- Balk, J.; Leaver, C.J. The PET1-CMS mitochondrial mutation in sunflower is associated with premature programmed cell death and cytochrome c release. Plant Cell 2001, 13, 1803–1818. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.P.; Xu, H.; Liu, Z.L.; Guo, J.X.; Li, H.Y.; Chen, L.T.; Fang, C.; Zhang, Q.Y.; Bai, M.; Yao, N.; et al. A detrimental mitochondrial-nuclear interaction causes cytoplasmic male sterility in rice. Nat. Genet. 2013, 45, 573–577. [Google Scholar] [CrossRef] [PubMed]
- He, T.T. Comparative Analysis of Mitochondrial Genomes of Soybean Cytoplasmic-Nuclear Male Sterile Lines and Their Maintainers and Mapping of Fertility Restoring Gene. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2017; pp. 21–65. [Google Scholar]
- Zeng, W.Y.; Yang, S.P.; Gai, J.Y.; Yu, D.Y. Proteomic study of anther differentiation between cytoplasmic-nuclear male sterile line NJCMS1A and its maintainer in soybean [Glycine max (L) Merr.]. Sci. Agric. Sin. 2007, 40, 2679–2687. [Google Scholar]
- Fan, J.M. Studies on Cyto-Morphological and Cyto-Chemical Features of Cytoplasmic-Nuclear Male-Sterile Lines of Soybeans (Glycine max (L.) Merr.). Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2003; pp. 32–41. [Google Scholar]
- Gai, J.Y.; Cui, Z.L.; Ji, D.F.; Ren, Z.J.; Ding, D.R. A report on the nuclear cytoplasmic male sterility from a cross between two soybean cultivars. Soy. Genet. Newsl. 1995, 22, 55–58. [Google Scholar]
- Ding, D.R.; Gai, J.Y.; Cui, Z.L.; Yang, S.P.; Qiu, J.X. Development and verification of the cytoplasmic-nuclear male sterile soybean line NJCMS1A and its maintainer NJCMS1B. Chin. Sci. Bull. 1999, 44, 191–192. [Google Scholar] [CrossRef]
- Ding, D.R.; Gai, J.Y.; Cui, Z.L.; Qiu, J.X. Development of a cytoplasmic-nuclear male-sterile line of soybean. Euphytica 2002, 124, 85–91. [Google Scholar] [CrossRef]
- Li, Y.W.; Ding, X.L.; Wang, X.; He, T.T.; Zhang, H.; Yang, L.S.; Wang, T.L.; Chen, L.F.; Gai, J.Y.; Yang, S.P. Genome wide comparative analysis of DNA methylation between soybean cytoplasmic male-sterile line NJCMS5A and its maintainer NJCMS5B. BMC Genom. 2017, 18, 596. [Google Scholar] [CrossRef]
- Gai, J.Y.; Hu, Y.Z.; Chen, J.M.; Gu, Y.J. Experiments on the retaining of pollen viability of soybeans. Acta Agron. Sin. 1980, 6, 11–16. [Google Scholar]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.S.; Zhang, H.Y.; Liu, X.Q.; Yu, S.B.; Xiong, L.Z.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant. 2013, 6, 1769–1780. [Google Scholar] [CrossRef]
- Fraga, C.G.; Clowers, B.H.; Moore, R.J.; Zink, E.M. Signature-discovery approach for sample matching of a nerve-agent precursor using liquid chromatography-mass spectrometry, XCMS, and chemometrics. Anal. Chem. 2010, 82, 4165–4173. [Google Scholar] [CrossRef]
- Eriksson, L.; Johansson, E.; Lindgren, F.; Sjostrom, M.; Wold, S. Megavariate analysis of hierarchical QSAR data. J. Comput. Aided Mol. Des. 2002, 16, 711–726. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Zhang, R.P.; Song, Y.M.; He, J.M.; Sun, J.H.; Bai, J.F.; An, Z.L.; Dong, L.J.; Zhan, Q.M.; Abliz, Z. RRLC-MS/MS-based metabonomics combined with in-depth analysis of metabolic correlation network: Finding potential biomarkers for breast cancer. Analyst 2009, 134, 2003–2011. [Google Scholar] [CrossRef] [PubMed]
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the human adult urinary metabolome variations with age, body mass index, and gender by implementing a comprehensive workflow for univariate and OPLS statistical analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Index | Compounds | VIP | Fold Change | Type |
|---|---|---|---|---|---|
| Flavanone | Glm558 | Naringenin O-malonylhexoside | 1.61 | 2.51 | up |
| Glm656 | Isoliquiritigenin | 1.47 | 2.17 | up | |
| Glm1180 | Hesperetin 7-rutinoside | 1.31 | −22.16 | down | |
| Glm1194 | Hesperetin 7-O-neohesperidoside | 1.52 | −19.58 | down | |
| Flavone | Glm541 | Apigenin O-malonylhexoside | 1.52 | 2.28 | up |
| Glm559 | Luteolin O-sinapoylhexoside | 1.57 | 2.59 | up | |
| Glm565 | Tricetin O-malonylhexoside | 1.46 | 2.19 | up | |
| Glm578 | Chrysin O-hexoside | 1.92 | 3.76 | up | |
| Glm590 | Tricin O-rhamnoside | 1.61 | 2.64 | up | |
| Glm594 | Chrysin 5-O-glucoside | 2.00 | 4.28 | up | |
| Glm1233 | 7,4′-Dihydroxyflavone | 1.44 | 2.08 | up | |
| Glm403 | Selgin 5-O-hexoside | 1.86 | −3.42 | down | |
| Glm694 | Nobiletin | 1.28 | −12.40 | down | |
| Glm710 | Tangeretin | 1.16 | −17.82 | down | |
| Glm1131 | Tricin O-saccharic acid | 1.77 | −3.04 | down | |
| Glm1169 | Tricin 5-O-hexoside | 1.38 | −2.07 | down | |
| Glm1182 | Tricin di-O-hexoside | 4.91 | −5025.93 | down | |
| Flavone C-glycosides | Glm383 | Isorhamnetin C-hexoside | 3.95 | 247.04 | up |
| Glm536 | Luteolin O-feruloylhexoside | 1.66 | 2.71 | up | |
| Glm278 | 8-C-hexosyl-hesperetin O-hexoside | 5.17 | −12,477.78 | down | |
| Glm419 | 8-C-hexosyl-luteolin O-hexoside | 1.42 | −2.15 | down | |
| Flavonol | Glm587 | Kaempferol 7-O-rhamnoside | 1.39 | 2.27 | up |
| Glm1286 | Syringetin | 1.80 | 3.27 | up | |
| Glm360 | Syringetin 3-O-hexoside | 1.63 | −2.56 | down | |
| Glm577 | Quercetin-3,4′-O-di-β-glucopyranoside | 3.36 | −3033.67 | down | |
| Glm640 | Isorhamnetin | 2.68 | −3.06 | down | |
| Glm1315 | 3,7-Di-O-methylquercetin | 4.35 | −794.44 | down | |
| Flavonolignan | Glm641 | Tricin 4′-O-β-guaiacylglycerol | 1.90 | 2.59 | up |
| Isoflavone | Glm584 | Daidzein | 1.53 | 2.31 | up |
| Glm626 | Genistein (4′,5,7-Trihydroxyisoflavone) | 1.50 | 2.28 | up | |
| Glm1212 | 6-Hydroxydaidzein | 1.56 | 2.52 | up | |
| Glm1249 | Glycitein | 1.71 | 2.80 | up |
| Index | Phenolamides | VIP | Fold Change | Type |
|---|---|---|---|---|
| Glm995 | N-p-Coumaroyl hydroxyagmatine | 1.35 | 2.13 | up |
| Glm144 | N-Caffeoylspermidine | 2.45 | −8.41 | down |
| Glm207 | N-p-Coumaroyl putrescine | 3.66 | −15.56 | down |
| Glm224 | N′-Feruloyl putrescine | 2.67 | −12.33 | down |
| Glm228 | N′-p-Coumaroyl putrescine | 2.78 | −9.23 | down |
| Glm240 | N′,N″,N‴-p-coumaroyl-cinnamoyl-caffeoyl spermidine | 1.62 | −2.58 | down |
| Glm253 | N-Feruloyl putrescine | 2.76 | −14.66 | down |
| Glm255 | N-Caffeoyl agmatine | 3.30 | −9.33 | down |
| Glm270 | N-p-Coumaroyl agmatine | 1.40 | −2.90 | down |
| Glm281 | N-Feruloyl agmatine | 4.05 | −50.00 | down |
| Glm358 | N′, N″-Feruloyl,caffeoylspermidine | 2.07 | −4.70 | down |
| Glm377 | N′, N″-di-p-coumaroylspermidine | 2.90 | −19.65 | down |
| Glm576 | N′,N″-p-coumaroyl-feruloyl putrescine | 3.65 | −108.53 | down |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Wang, X.; Li, Q.; Yu, L.; Song, Q.; Gai, J.; Yang, S. Metabolomics Studies on Cytoplasmic Male Sterility during Flower Bud Development in Soybean. Int. J. Mol. Sci. 2019, 20, 2869. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122869
Ding X, Wang X, Li Q, Yu L, Song Q, Gai J, Yang S. Metabolomics Studies on Cytoplasmic Male Sterility during Flower Bud Development in Soybean. International Journal of Molecular Sciences. 2019; 20(12):2869. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122869
Chicago/Turabian StyleDing, Xianlong, Xuan Wang, Qiang Li, Lifeng Yu, Qijian Song, Junyi Gai, and Shouping Yang. 2019. "Metabolomics Studies on Cytoplasmic Male Sterility during Flower Bud Development in Soybean" International Journal of Molecular Sciences 20, no. 12: 2869. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122869