Trifluoperazine, an Antipsychotic Drug, Effectively Reduces Drug Resistance in Cisplatin-Resistant Urothelial Carcinoma Cells via Suppressing Bcl-xL: An In Vitro and In Vivo Study

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
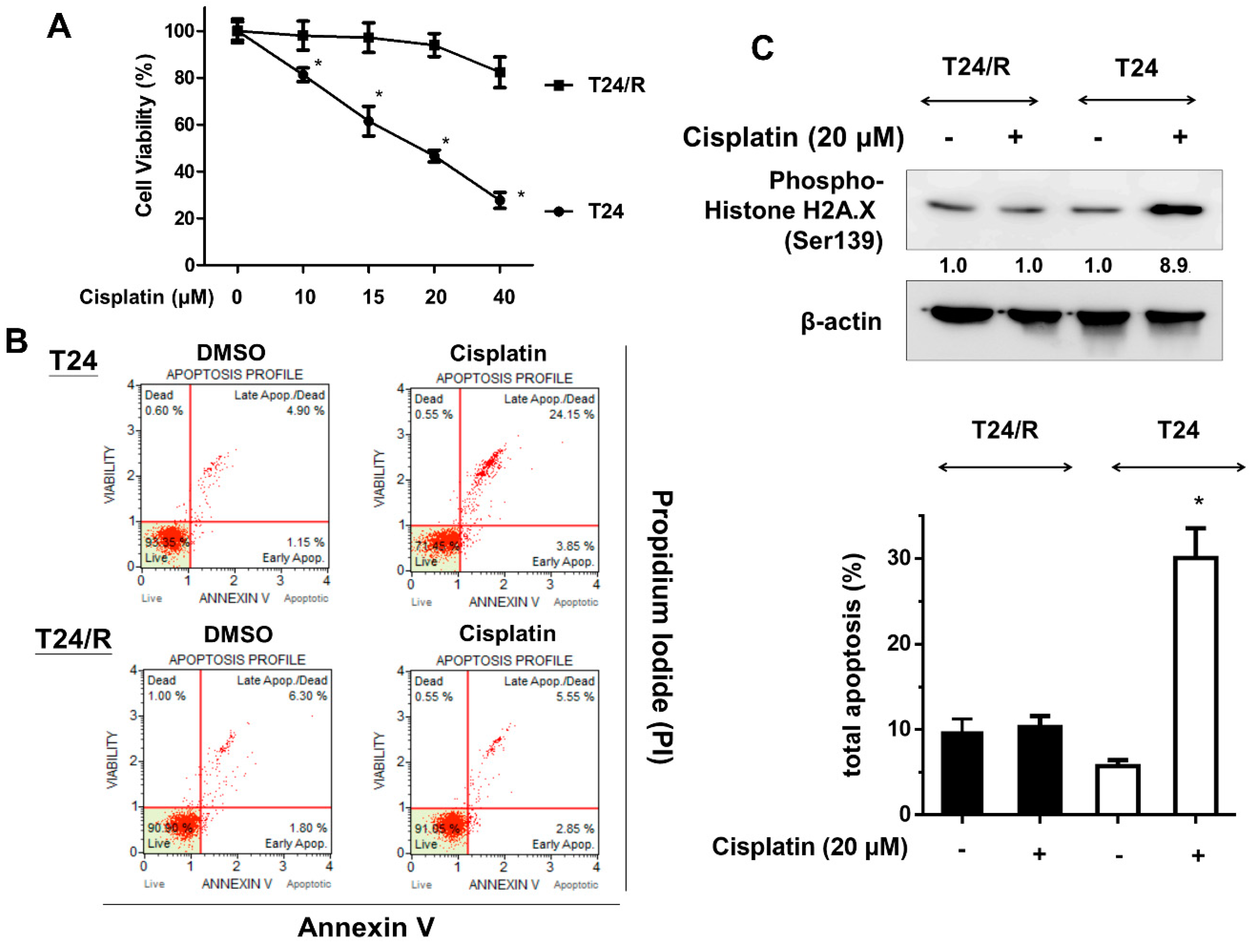
2.1. Cisplatin-Induced Cytotoxicity, Apoptosis, and DNA Damage Response Were Reduced in Cisplatin-Resistant UC Cells (T24/R) Compared with Parental T24 Cells
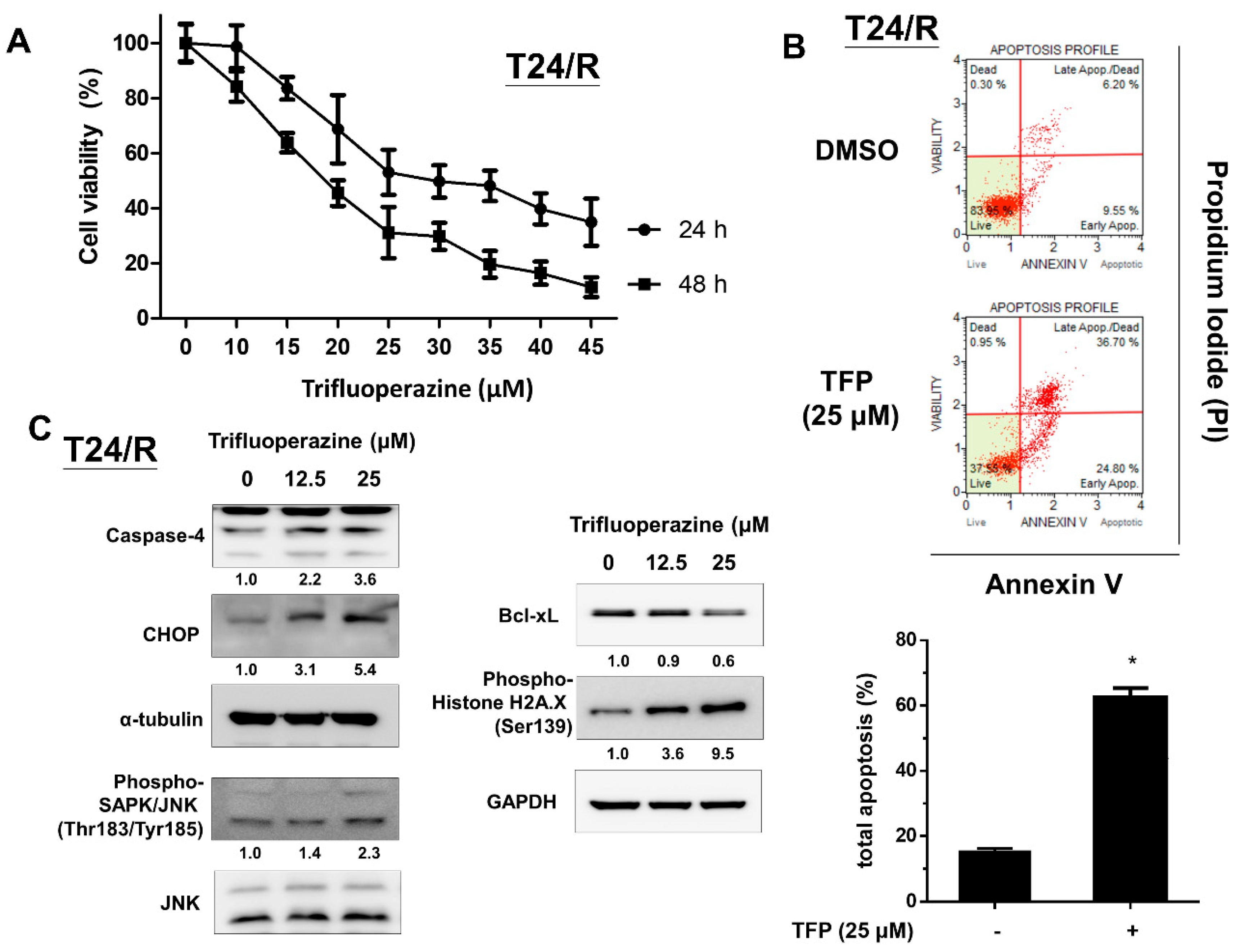
2.2. TFP Effectively Induced Cytotoxicity, Apoptosis, Endoplasmic Reticulum Stress-Related Apoptosis, and DNA Damage in Cisplatin-Resistant Human UC Cells (T24/R)
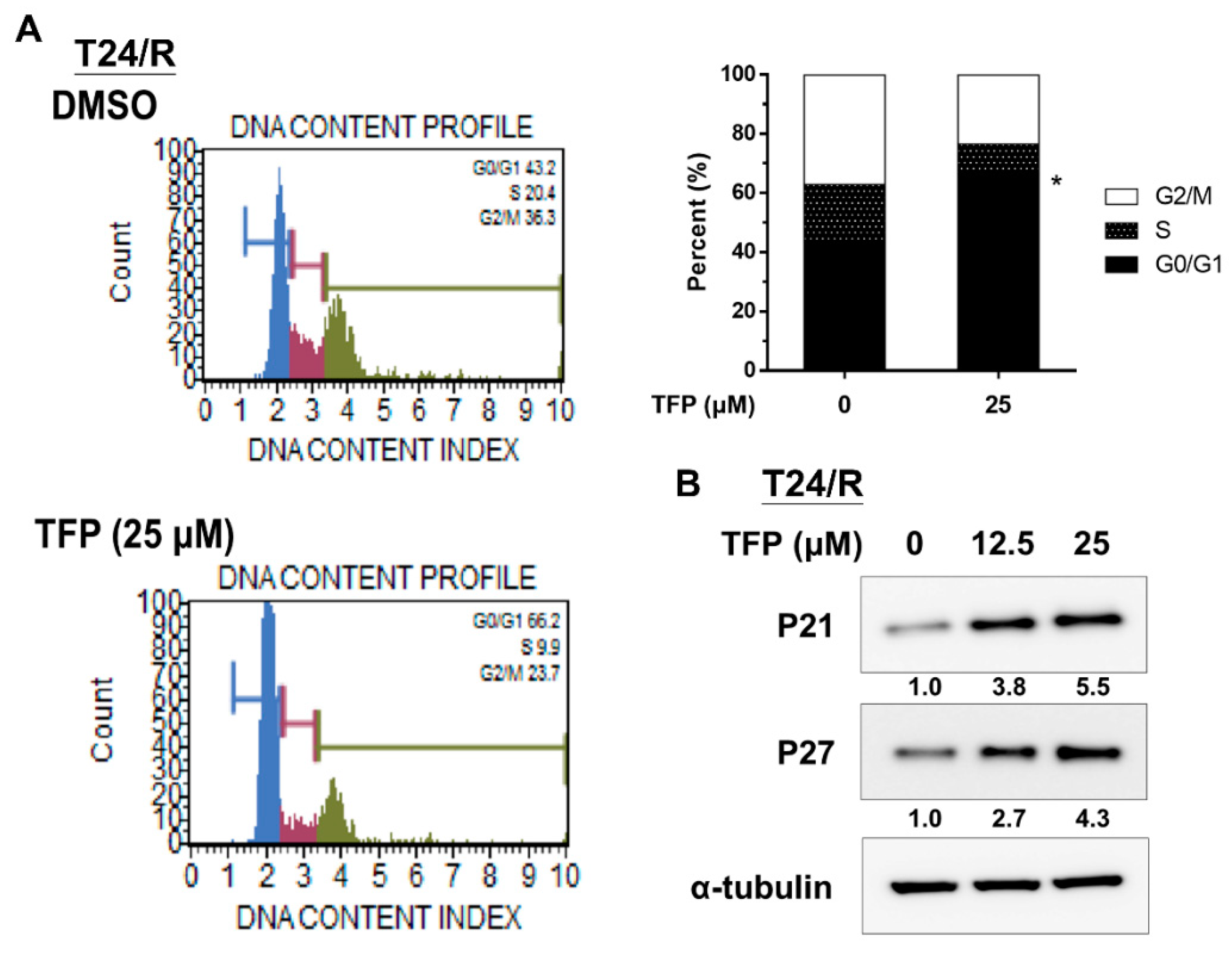
2.3. TFP Induced G0/G1 Arrest in Cisplatin-Resistant UC Cells (T24/R)
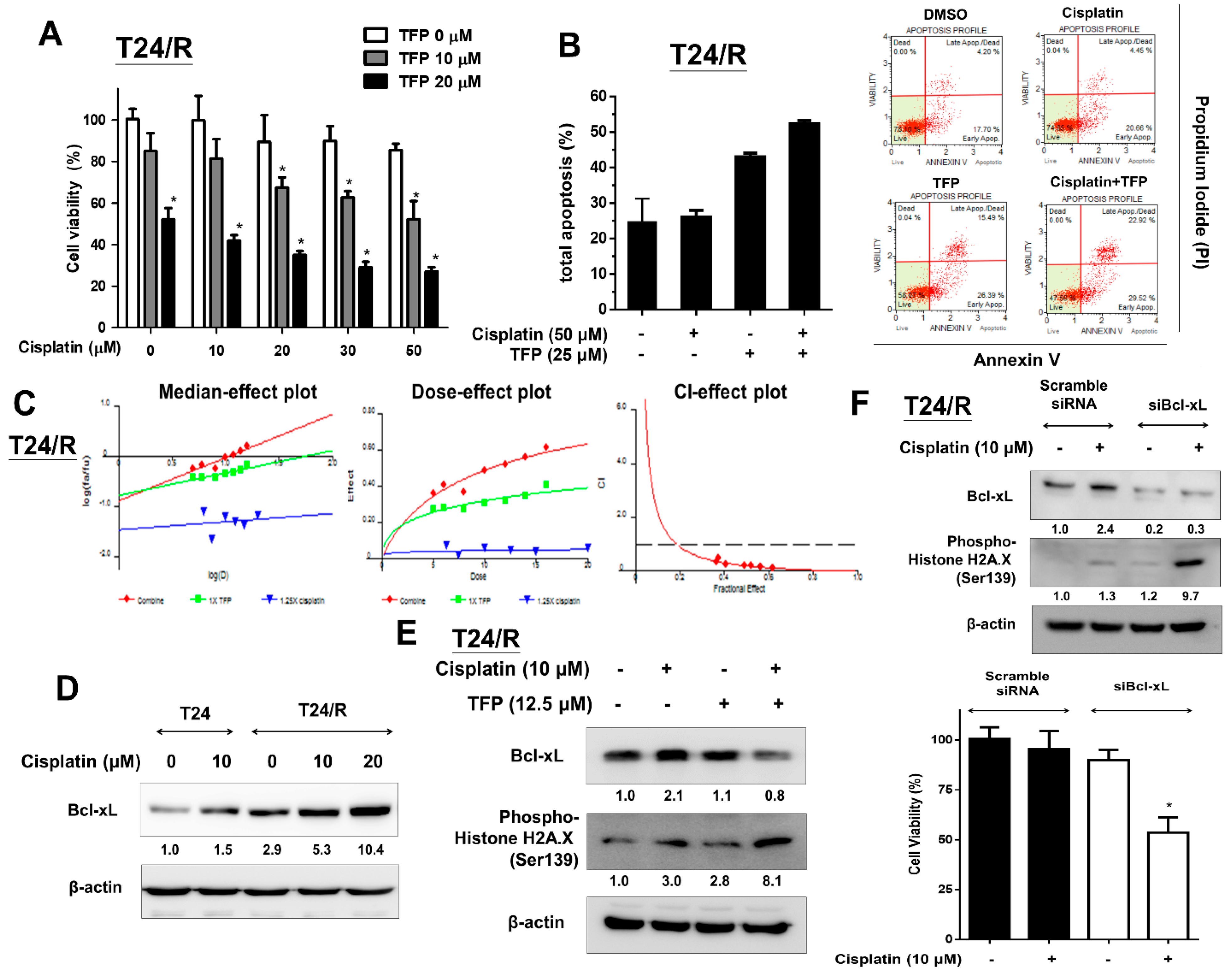
2.4. TFP Enhanced the Cisplatin Antitumor Effects and Alleviated Cisplatin Resistance with Concurrent Bcl-xL Suppression in T24/R Cells
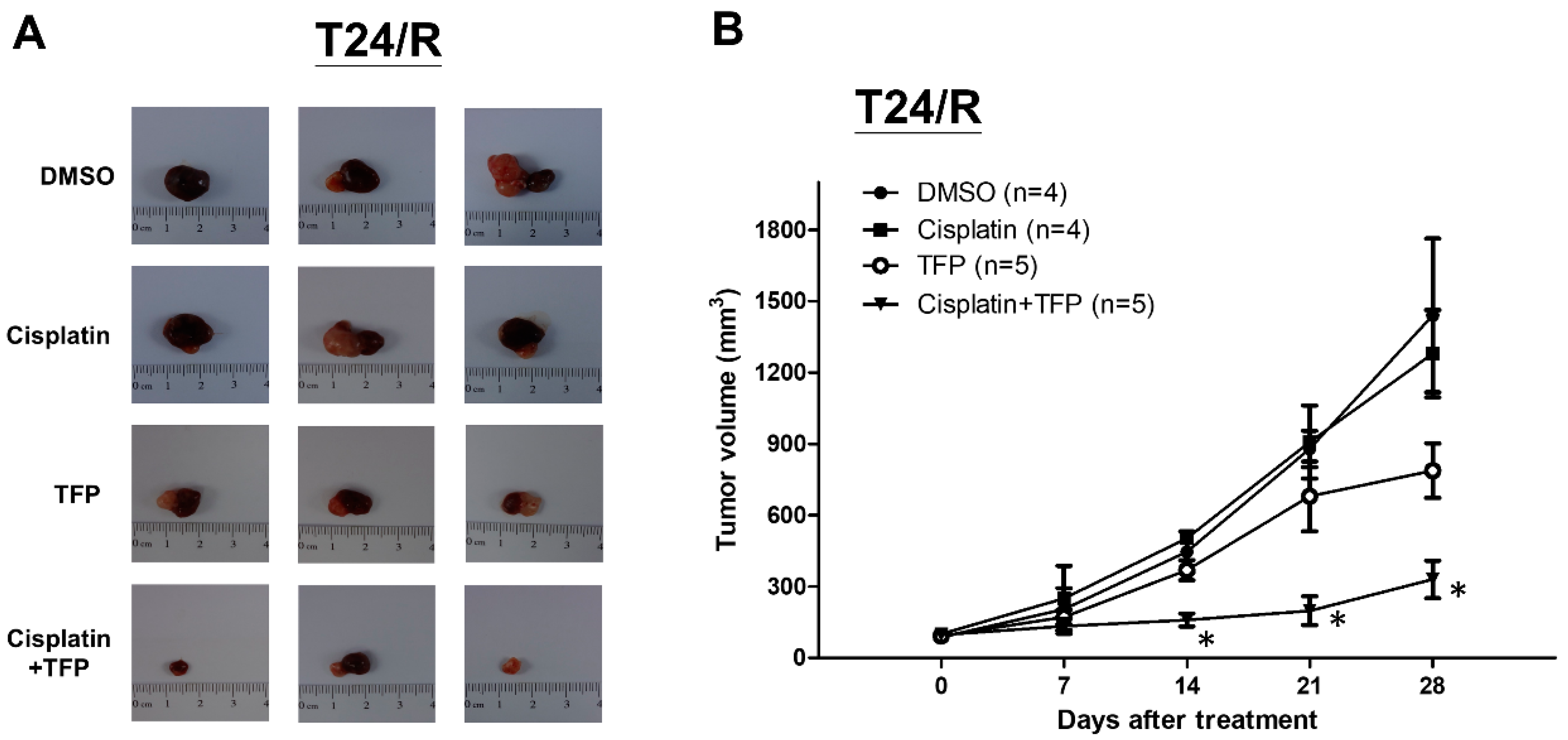
2.5. TFP Enhanced Antitumor Effect of Cisplatin in a Xenograft Mouse Model of Cisplatin-Resistant UC Cells (T24/R)
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Reagents and Antibodies
4.3. Measurement of Cell Viability
4.4. Knockdown of BcL-xL Using siRNA
4.5. Western Blot Analysis
4.6. Apoptosis Assay
4.7. Cell Cycle Analysis by Flow Cytometry
4.8. Combinative Drug Effects
4.9. In Vivo Xenograft Experiments
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Bcl-xL | B-cell lymphoma-extra large |
| UC | urothelial carcinoma |
| TFP | Trifluoperazine |
| IC50 | half maximal inhibitory concentration |
References
- American Cancer Society. Cancer Facts & Figures 2015. 2015. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2015.html (accessed on 29 June 2019).
- Bellmunt, J.; Albiol, S.; Suarez, C.; Albanell, J. Optimizing therapeutic strategies in advanced bladder cancer: Update on chemotherapy and the role of targeted agents. Crit. Rev. Oncol. Hematol. 2009, 69, 211–222. [Google Scholar] [CrossRef] [PubMed]
- von der Maase, H.; Hansen, S.W.; Roberts, J.T.; Dogliotti, L.; Oliver, T.; Moore, M.J.; Bodrogi, I.; Albers, P.; Knuth, A.; Lippert, C.M.; et al. Gemcitabine and cisplatin versus methotrexate, vinblastine, doxorubicin, and cisplatin in advanced or metastatic bladder cancer: Results of a large, randomized, multinational, multicenter, phase III study. J. Clin. Oncol. 2000, 18, 3068–3077. [Google Scholar] [CrossRef] [PubMed]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharm. 2014, 740, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulos, K. Targeting the Bcl-2 family in cancer therapy. Semin. Oncol. 2006, 33, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Minn, A.J.; Rudin, C.M.; Boise, L.H.; Thompson, C.B. Expression of bcl-xL can confer a multidrug resistance phenotype. Blood 1995, 86, 1903–1910. [Google Scholar] [PubMed]
- Lei, X.; Huang, Z.; Zhong, M.; Zhu, B.; Tang, S.; Liao, D. Bcl-XL small interfering RNA sensitizes cisplatin-resistant human lung adenocarcinoma cells. Acta Biochim. Biophys. Sin. (Shanghai) 2007, 39, 344–350. [Google Scholar] [CrossRef] [PubMed]
- Villedieu, M.; Louis, M.H.; Dutoit, S.; Brotin, E.; Lincet, H.; Duigou, F.; Staedel, C.; Gauduchon, P.; Poulain, L. Absence of Bcl-xL down-regulation in response to cisplatin is associated with chemoresistance in ovarian carcinoma cells. Gynecol. Oncol. 2007, 105, 31–44. [Google Scholar] [CrossRef]
- Yu, L.; Wang, Z. Difference in expression of Bcl-2 and Bcl-xl genes in cisplatin-sensitive and cisplatin-resistant human in ovarian cancer cell lines. J. Huazhong Univ. Sci. Technol. Med. Sci. 2004, 24, 151–153. [Google Scholar]
- Thomas, S.; Quinn, B.A.; Das, S.K.; Dash, R.; Emdad, L.; Dasgupta, S.; Wang, X.Y.; Dent, P.; Reed, J.C.; Pellecchia, M.; et al. Targeting the Bcl-2 family for cancer therapy. Expert. Opin. Ther. Targets 2013, 17, 61–75. [Google Scholar] [CrossRef]
- Yamaguchi, R.; Lartigue, L.; Perkins, G. Targeting Mcl-1 and other Bcl-2 family member proteins in cancer therapy. Pharmacol. Ther. 2019, 195, 13–20. [Google Scholar] [CrossRef]
- Dai, Y.; Jin, S.; Li, X.; Wang, D. The involvement of Bcl-2 family proteins in AKT-regulated cell survival in cisplatin resistant epithelial ovarian cancer. Oncotarget 2017, 8, 1354–1368. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Xia, Y.; Gao, T.; Xu, F.; Lei, Q.; Peng, C.; Yang, Y.; Xue, Q.; Hu, X.; Wang, Q.; et al. The antipsychotic agent trifluoperazine hydrochloride suppresses triple-negative breast cancer tumor growth and brain metastasis by inducing G0/G1 arrest and apoptosis. Cell Death Dis. 2018, 9, 1006. [Google Scholar] [CrossRef]
- Shin, S.Y.; Kim, C.G.; Hong, D.D.; Kim, J.H.; Lee, Y.H. Implication of Egr-1 in trifluoperazine-induced growth inhibition in human U87MG glioma cells. Exp. Mol. Med. 2004, 36, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.W.; Hickie, R.A.; Klaassen, D.J. Inhibition of human breast cancer colony formation by anticalmodulin agents: Trifluoperazine, W-7, and W-13. Cancer Chemother. Pharm. 1983, 11, 86–90. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Wu, L.J.; Wu, Y.Q.; Lu, G.H.; Jiang, Z.Y.; Zhan, J.W.; Jie, Y.; Zhou, J.Y. Molecular mechanism of trifluoperazine induces apoptosis in human A549 lung adenocarcinoma cell lines. Mol. Med. Rep. 2009, 2, 811–817. [Google Scholar] [CrossRef]
- Qian, K.; Sun, L.; Zhou, G.; Ge, H.; Meng, Y.; Li, J.; Li, X.; Fang, X. Trifluoperazine as an alternative strategy for the inhibition of tumor growth of colorectal cancer. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, S.; Shiraishi, N.; Kuratomi, Y.; Nakagawa, M.; Kuwano, M. Circumvention of multiple-drug resistance in human cancer cells by thioridazine, trifluoperazine, and chlorpromazine. J. Natl. Cancer Inst. 1986, 76, 839–844. [Google Scholar]
- Shin, S.Y.; Choi, B.H.; Kim, J.R.; Kim, J.H.; Lee, Y.H. Suppression of P-glycoprotein expression by antipsychotics trifluoperazine in adriamycin-resistant L1210 mouse leukemia cells. Eur. J. Pharm. Sci 2006, 28, 300–306. [Google Scholar] [CrossRef]
- Murren, J.R.; Durivage, H.J.; Buzaid, A.C.; Reiss, M.; Flynn, S.D.; Carter, D.; Hait, W.N. Trifluoperazine as a modulator of multidrug resistance in refractory breast cancer. Cancer Chemother. Pharm. 1996, 38, 65–70. [Google Scholar] [CrossRef]
- Wang, B.; Luo, Y.; Zhou, X.; Li, R. Trifluoperazine induces apoptosis through the upregulation of Bax/Bcl2 and downregulated phosphorylation of AKT in mesangial cells and improves renal function in lupus nephritis mice. Int. J. Mol. Med. 2018, 41, 3278–3286. [Google Scholar]
- Chen, X.; Luo, X.; Cheng, Y. Trifluoperazine prevents FOXO1 nuclear excretion and reverses doxorubicin-resistance in the SHG44/DOX drug-resistant glioma cell line. Int. J. Mol. Med. 2018, 42, 3300–3308. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.T.; Wu, A.T.; Chang, P.M.; Chen, K.Y.; Yang, C.N.; Yang, S.C.; Ho, C.C.; Chen, C.C.; Kuo, Y.L.; Lee, P.Y.; et al. Trifluoperazine, an antipsychotic agent, inhibits cancer stem cell growth and overcomes drug resistance of lung cancer. Am. J. Respir. Crit. Care Med. 2012, 186, 1180–1188. [Google Scholar] [CrossRef] [PubMed]
- Burlacu, A. Regulation of apoptosis by Bcl-2 family proteins. J. Cell. Mol. Med. 2003, 7, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Delbridge, A.R.; Grabow, S.; Strasser, A.; Vaux, D.L. Thirty years of BCL-2: Translating cell death discoveries into novel cancer therapies. Nat. Rev. Cancer 2016, 16, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Delbridge, A.R.; Strasser, A. The BCL-2 protein family, BH3-mimetics and cancer therapy. Cell Death Differ. 2015, 22, 1071–1080. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Guo, W.; Zhang, L.; Davis, J.J.; Wu, S.; Teraishi, F.; Cao, X.; Smythe, W.R.; Fang, B. Enhancing TRAIL-induced apoptosis by Bcl-X(L) siRNA. Cancer Biol. Ther. 2005, 4, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Mu, P.; Nagahara, S.; Makita, N.; Tarumi, Y.; Kadomatsu, K.; Takei, Y. Systemic delivery of siRNA specific to tumor mediated by atelocollagen: Combined therapy using siRNA targeting Bcl-xL and cisplatin against prostate cancer. Int. J. Cancer 2009, 125, 2978–2990. [Google Scholar] [CrossRef]
- Basu, A.; Krishnamurthy, S. Cellular responses to Cisplatin-induced DNA damage. J. Nucleic. Acids 2010. [Google Scholar] [CrossRef]
- Sun, Q.; Yogosawa, S.; Iizumi, Y.; Sakai, T.; Sowa, Y. The alkaloid emetine sensitizes ovarian carcinoma cells to cisplatin through downregulation of bcl-xL. Int. J. Oncol. 2015, 46, 389–394. [Google Scholar] [CrossRef]
- Simonian, P.L.; Grillot, D.A.; Merino, R.; Nunez, G. Bax can antagonize Bcl-XL during etoposide and cisplatin-induced cell death independently of its heterodimerization with Bcl-XL. J. Biol. Chem. 1996, 271, 22764–22772. [Google Scholar] [CrossRef]
- Polischouk, A.G.; Holgersson, A.; Zong, D.; Stenerlow, B.; Karlsson, H.L.; Moller, L.; Viktorsson, K.; Lewensohn, R. The antipsychotic drug trifluoperazine inhibits DNA repair and sensitizes non small cell lung carcinoma cells to DNA double-strand break induced cell death. Mol. Cancer Ther. 2007, 6, 2303–2309. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.C.; Hsu, F.S.; Kuo, K.L.; Liu, S.H.; Shun, C.T.; Shi, C.S.; Chang, H.C.; Tsai, Y.C.; Lin, M.C.; Wu, J.T.; et al. Trichostatin A, a histone deacetylase inhibitor, induces synergistic cytotoxicity with chemotherapy via suppression of Raf/MEK/ERK pathway in urothelial carcinoma. J. Mol. Med. (Berl) 2018, 96, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.S.; Wu, J.T.; Lin, J.Y.; Yang, S.P.; Kuo, K.L.; Lin, W.C.; Shi, C.S.; Chow, P.M.; Liao, S.M.; Pan, C.I.; et al. Histone Deacetylase Inhibitor, Trichostatin A, Synergistically Enhances Paclitaxel-Induced Cytotoxicity in Urothelial Carcinoma Cells by Suppressing the ERK Pathway. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Ho, I.L.; Kuo, K.L.; Liu, S.H.; Chang, H.C.; Hsieh, J.T.; Wu, J.T.; Chiang, C.K.; Lin, W.C.; Tsai, Y.C.; Chou, C.T.; et al. MLN4924 Synergistically Enhances Cisplatin-induced Cytotoxicity via JNK and Bcl-xL Pathways in Human Urothelial Carcinoma. Sci. Rep. 2015, 5, 16948. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuo, K.-L.; Liu, S.-H.; Lin, W.-C.; Hsu, F.-S.; Chow, P.-M.; Chang, Y.-W.; Yang, S.-P.; Shi, C.-S.; Hsu, C.-H.; Liao, S.-M.; et al. Trifluoperazine, an Antipsychotic Drug, Effectively Reduces Drug Resistance in Cisplatin-Resistant Urothelial Carcinoma Cells via Suppressing Bcl-xL: An In Vitro and In Vivo Study. Int. J. Mol. Sci. 2019, 20, 3218. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133218
Kuo K-L, Liu S-H, Lin W-C, Hsu F-S, Chow P-M, Chang Y-W, Yang S-P, Shi C-S, Hsu C-H, Liao S-M, et al. Trifluoperazine, an Antipsychotic Drug, Effectively Reduces Drug Resistance in Cisplatin-Resistant Urothelial Carcinoma Cells via Suppressing Bcl-xL: An In Vitro and In Vivo Study. International Journal of Molecular Sciences. 2019; 20(13):3218. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133218
Chicago/Turabian StyleKuo, Kuan-Lin, Shing-Hwa Liu, Wei-Chou Lin, Fu-Shun Hsu, Po-Ming Chow, Yu-Wei Chang, Shao-Ping Yang, Chung-Sheng Shi, Chen-Hsun Hsu, Shih-Ming Liao, and et al. 2019. "Trifluoperazine, an Antipsychotic Drug, Effectively Reduces Drug Resistance in Cisplatin-Resistant Urothelial Carcinoma Cells via Suppressing Bcl-xL: An In Vitro and In Vivo Study" International Journal of Molecular Sciences 20, no. 13: 3218. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133218