Acute and Chronic Sleep Deprivation-Related Changes in N-methyl-D-aspartate Receptor—Nitric Oxide Signalling in the Rat Cerebral Cortex with Reference to Aging and Brain Lateralization

Abstract
:1. Introduction
2. Results
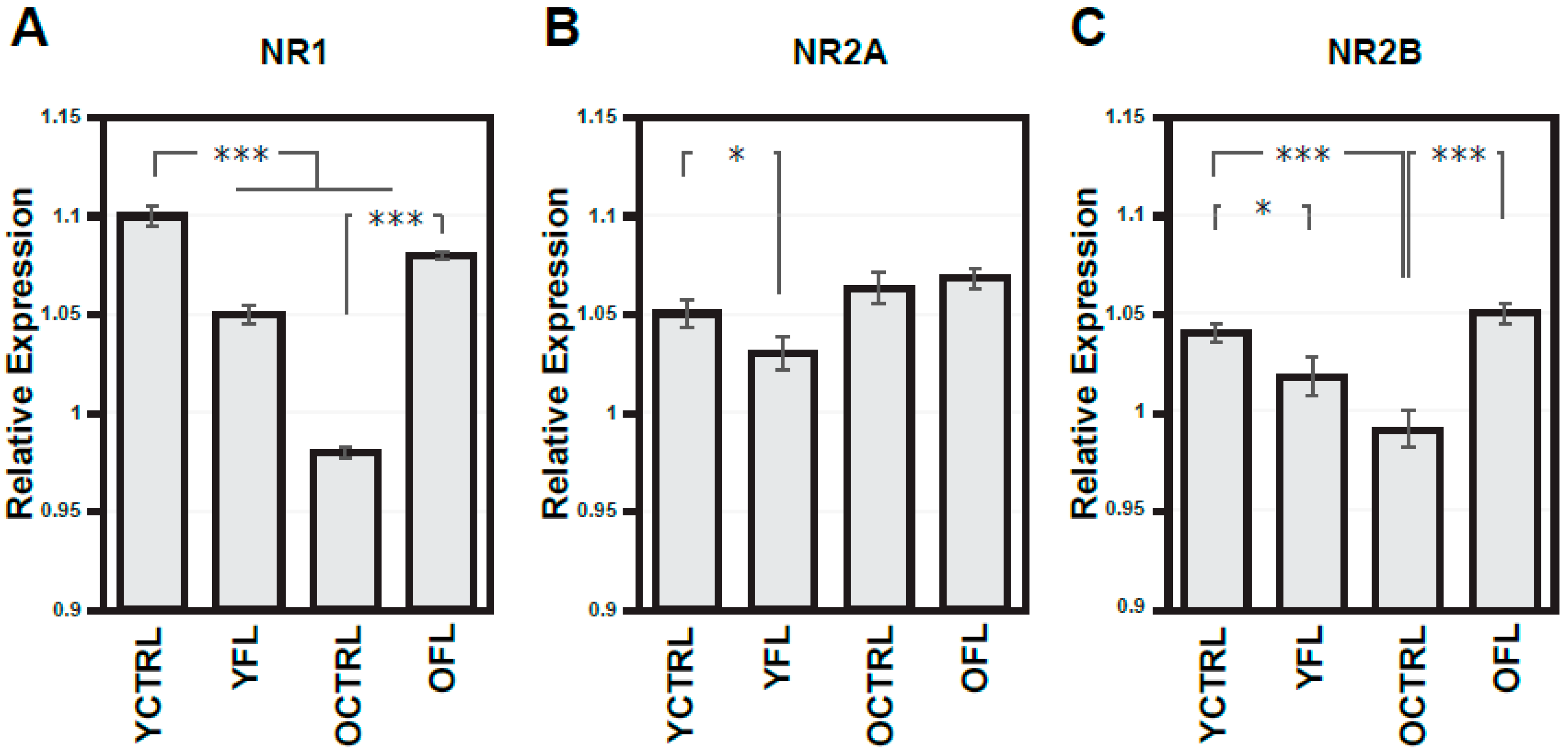
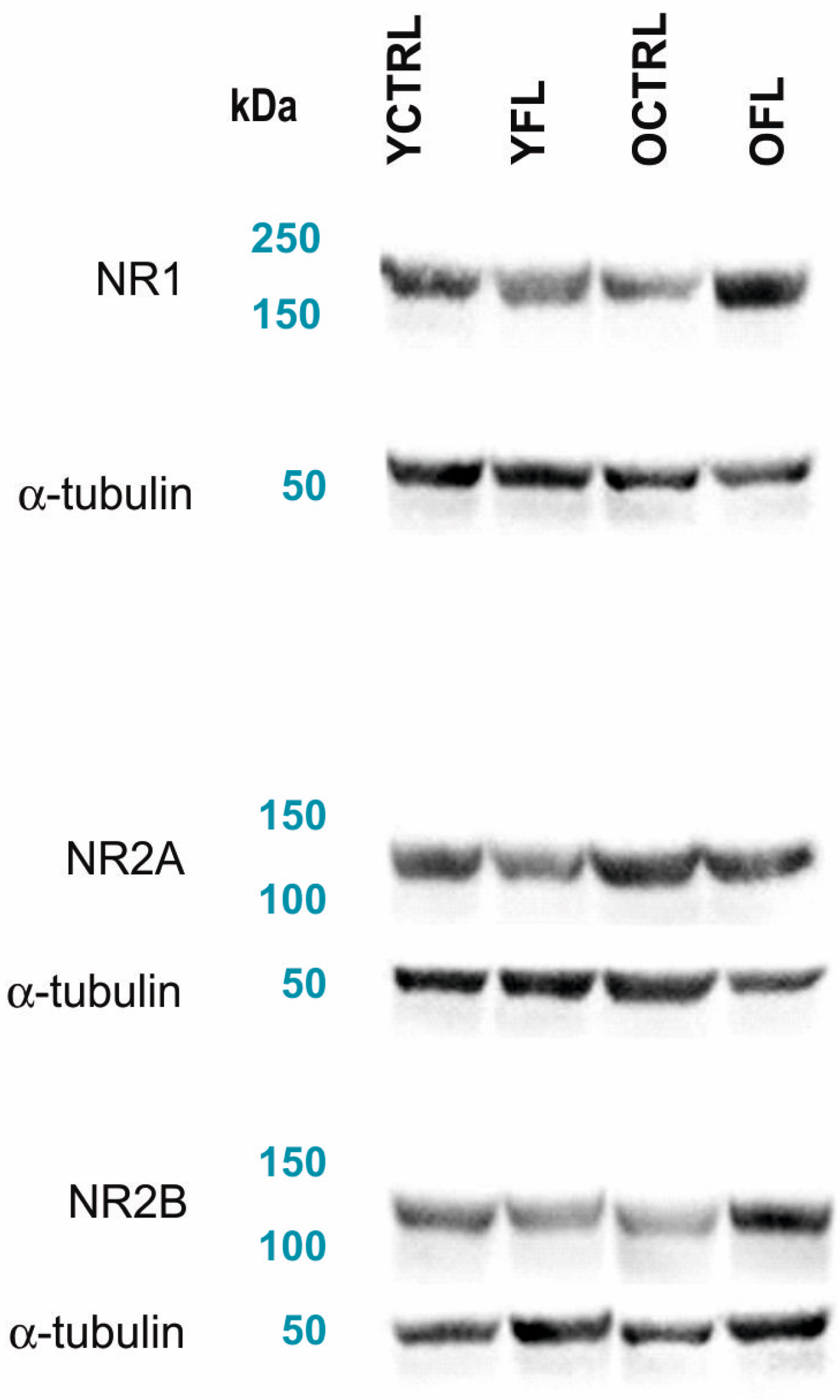
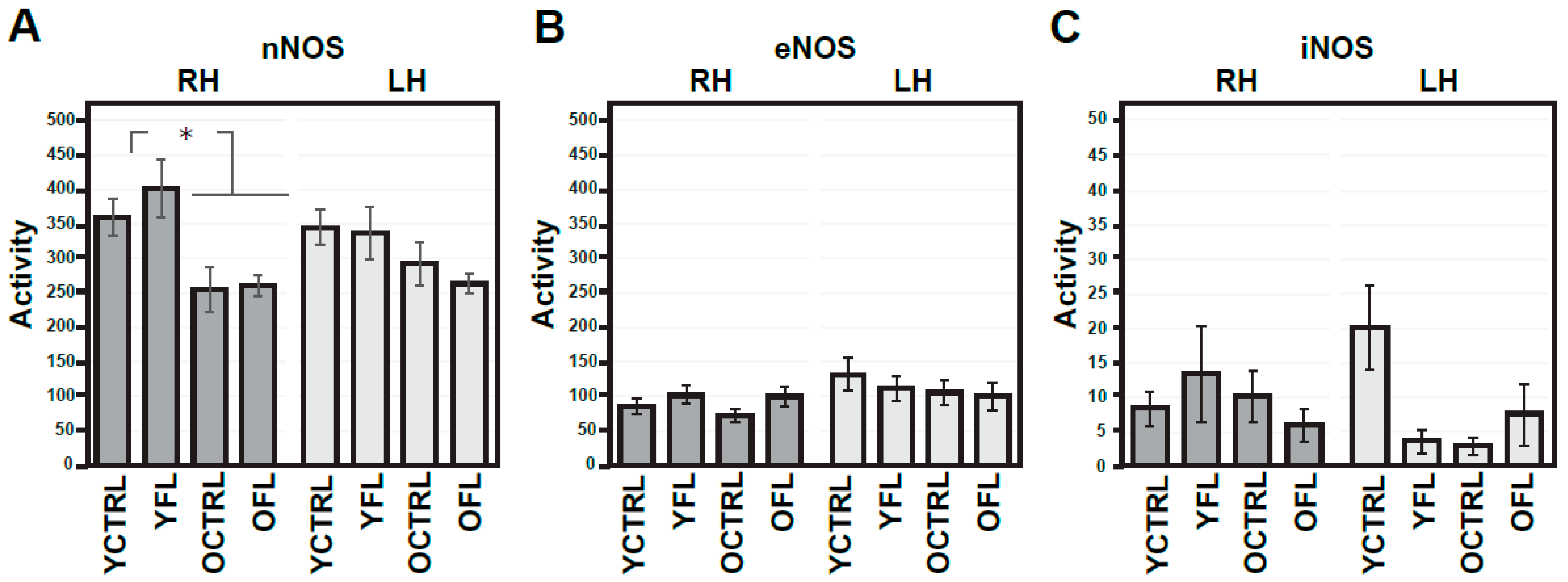
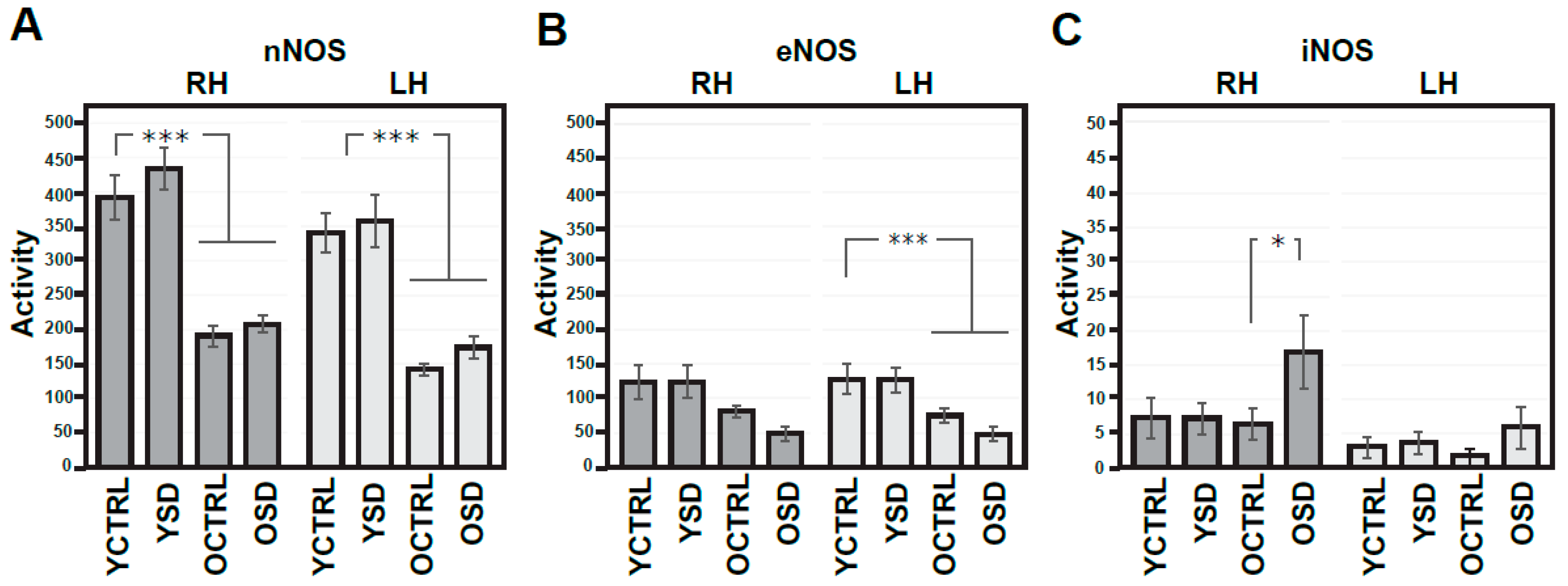
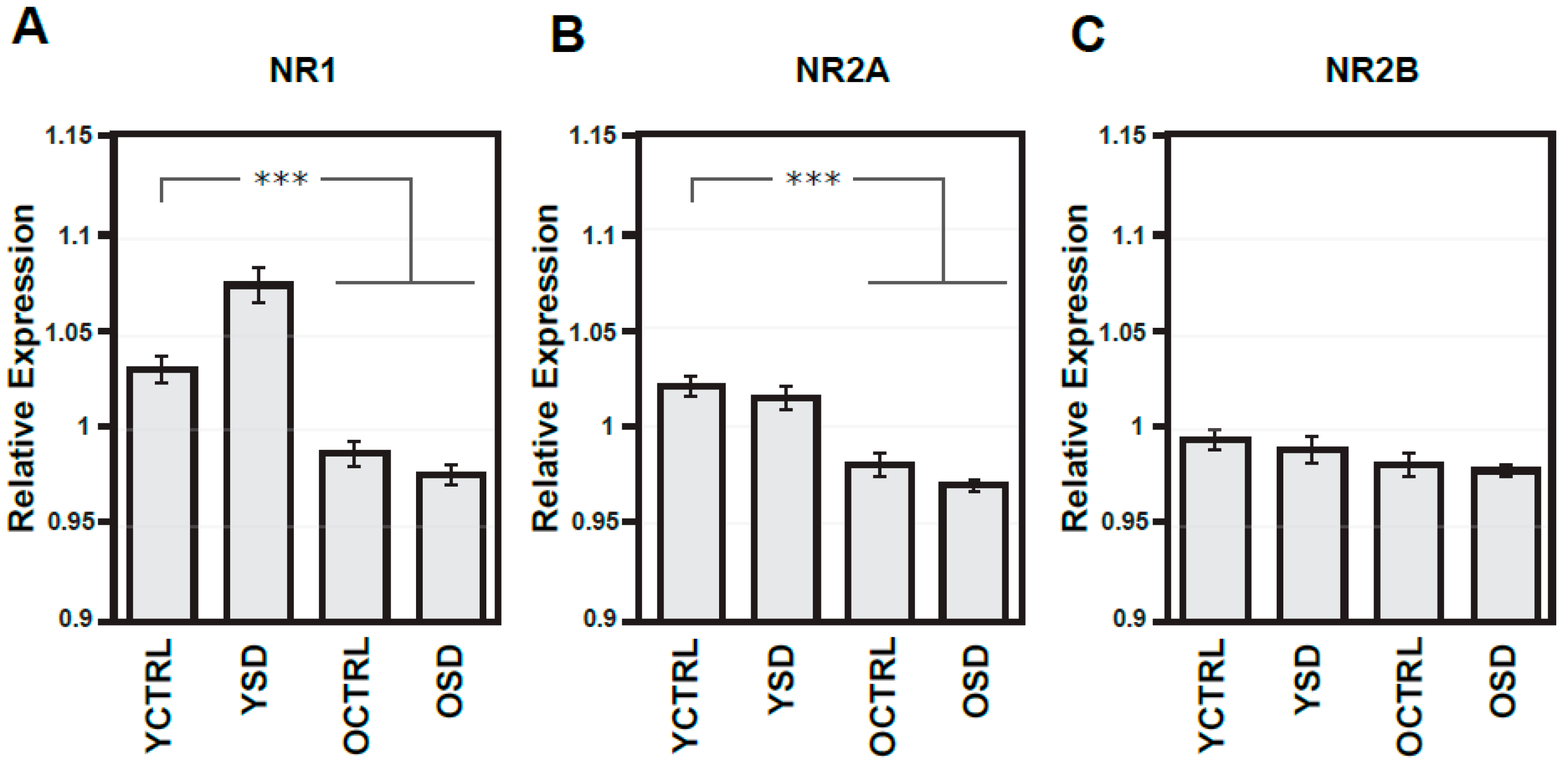
2.1. Changes in NMDA Receptors Subunits and NOS Activities Related to FL (Experiment I)
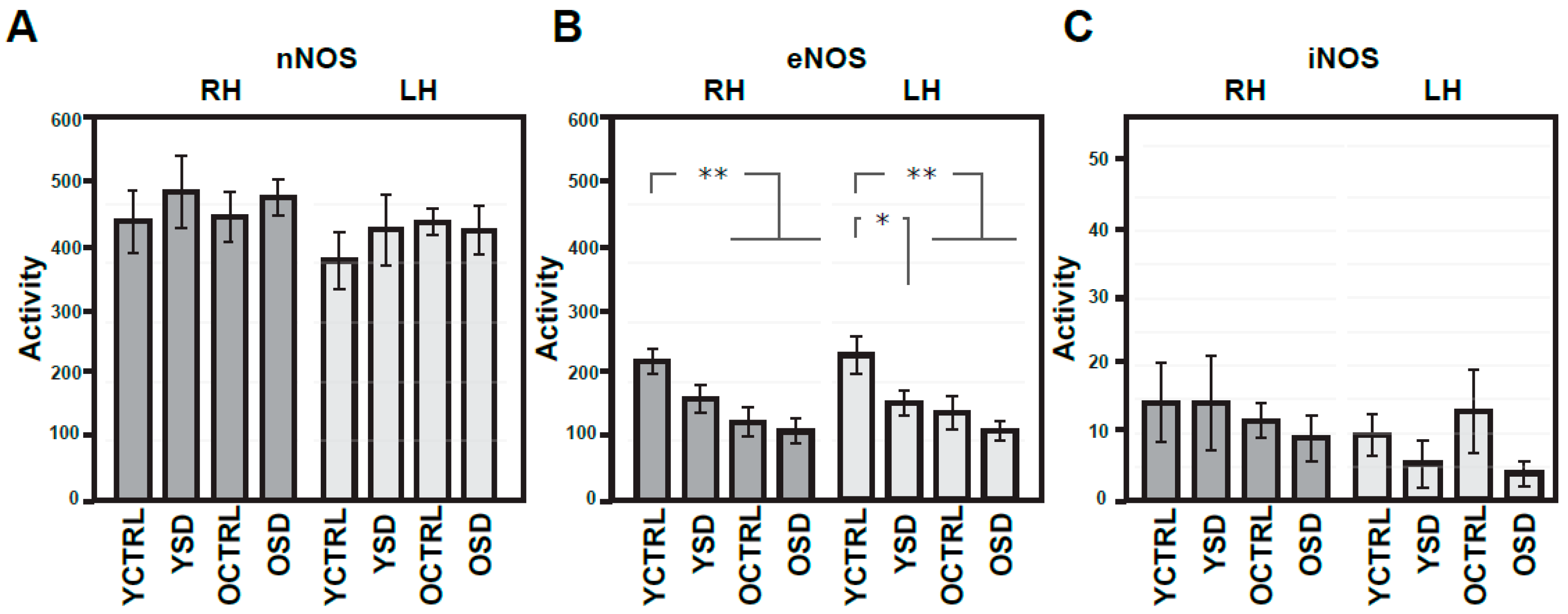
2.2. Changes in NMDA Receptors Subunits and NOS Activities Related to Acute SD (Experiment II)
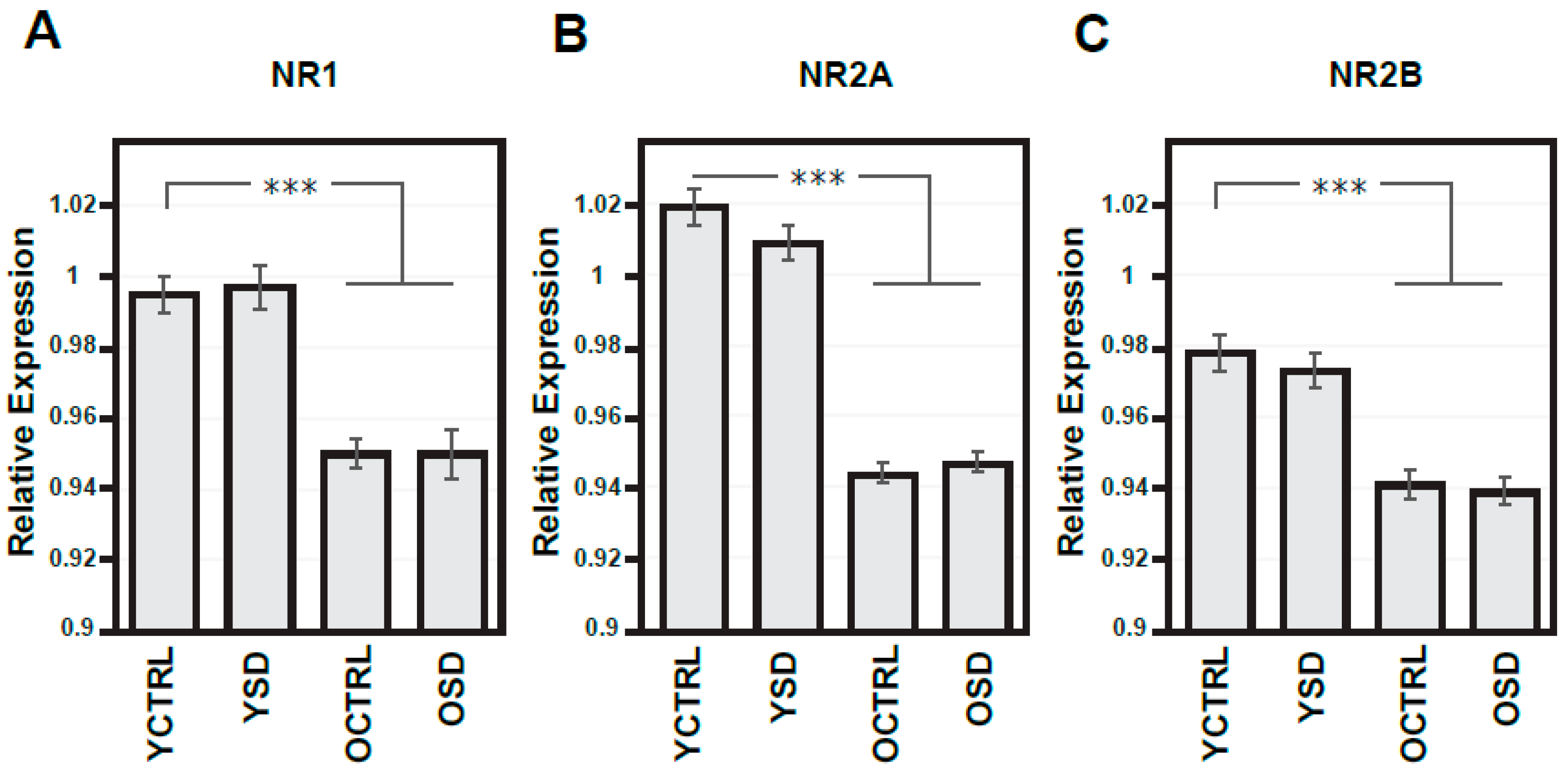
2.3. Changes in NMDA Receptors Subunits and NOS Activities Related to Chronic SD (Experiment III)
2.4. Age-Dependent Changes in NMDA Receptors Subunits and NOS Activities (Experiments I–III)
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Sleep Deprivation Experiments
4.3. Analytical Biochemistry Experiments
4.4. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| SD | sleep deprivation |
| AD | Alzheimer’s disease |
| NMDA | N-methyl-D-aspartate |
| NO | nitric oxide |
| REM | rapid eye movement |
| FL | forced locomotion |
| nNOS | neural nitric oxide synthase |
| eNOS | endothelial nitric oxide synthase |
| iNOS | inducible nitric oxide synthase |
| Aβ | amyloid β peptide |
| PBS buffer | phosphate-buffered saline |
| TBS buffer | Tris-buffered saline |
| R | right |
| L | left |
References
- Costandi, M. Amyloid awakenings. Nature 2013, 497, 19–20. [Google Scholar] [CrossRef] [PubMed]
- Huitron-Resendiz, S.; Sanchez-Alavez, M.; Gallegos, R.; Berg, G.; Crawford, E.; Giacchino, J.L.; Games, D.; Henriksen, S.J.; Criado, J.R. Age-independent and age-related deficits in visuospatial learning, sleep-wake states, thermoregulation and motor activity in PDAPP mice. Brain Res. 2002, 928, 126–137. [Google Scholar] [CrossRef]
- Petrasek, T.; Vojtechova, I.; Lobellova, V.; Popelikova, A.; Janikova, M.; Broyka, H.; Houdek, P.; Sladek, M.; Sumova, A.; Kristofikova, Z.; et al. The McGill transgenic rat model of Alzheimer’s disease displays cognitive and motor impairments, changes in anxiety and social behavior, and altered circadian activity. Front. Aging Neurosci. 2018, 10, 250. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.H.; Abilio, V.C.; Takatsu, A.L.; Kameda, S.R.; Grassl, C.; Chehin, A.B.; Medrano, W.A.; Calzavara, M.B.; Registro, S.; Andersen, M.L.; et al. Role of hippocampal oxidative stress in memory deficits induced by sleep deprivation in mice. Neuropharmacology 2004, 46, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Drake, J.; Pocernich, C.; Castegna, A. Evidence of oxidative damage in Alzheimer’s brain: Central role for amyloid beta-peptide. Trends Mol. Med. 2001, 7, 548–554. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Howard, B.J.; LaFontaine, M.A. Brain oxidative stress in animal models of accelerated aging and the age-related neurodegenerative disorders, Alzheimer’s disease and Huntington’s disease. Curr. Med. Chem. 2001, 8, 815–858. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.E.; Lim, M.M.; Bateman, R.J.; Lee, J.J.; Smyth, L.P.; Cirrito, J.R.; Fujiki, N.; Nishino, S.; Holtzman, D.M. Amyloid-β dynamics are regulated by orexin and the sleep-wake cycle. Science 2009, 326, 1005–1007. [Google Scholar] [CrossRef] [PubMed]
- Spira, A.P.; Gamaldo, A.A.; An, Y.; Wu, M.N.; Simonsick, E.M.; Bilgel, M.; Zhou, Y.; Wong, D.F.; Ferucci, L.; Resnick, S.M. Self-reported sleep and β-amyloid deposition in community-dwelling older adults. JAMA Neurol. 2013. [Google Scholar] [CrossRef]
- Shen, Y.X.; Xu, S.Y.; Wei, W.; Wang, X.L.; Wang, H.; Sun, X. Melatonin blocks rat hippocampal neuronal apoptosis induced by amyloid beta-peptide 25-35. J. Pineal Res. 2002, 32, 163–167. [Google Scholar] [CrossRef]
- Matsubara, E.; Bryant-Thomas, T.; Pacheco Quinto, J.; Henry, T.L.; Poeggeler, B.; Herbert, D.; Crus-Sanchez, F.; Chyan, Y.J.; Smith, M.A.; Perry, G.; et al. Melatonin increases survival and inhibits oxidative and amyloid pathology in a transgenic model of Alzheimer’s disease. J. Neurochem. 2003, 85, 1101–1108. [Google Scholar] [CrossRef]
- Lahiri, D.K.; Cen, D.; Ge, Y.W.; Bondy, S.C.; Sharman, E.H. Dietary supplementation with melatonin reduces levels of amyloid beta-peptides in the murine cerebral cortex. J. Pineal Res. 2004, 36, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Quin, J.; Kulhanek, D.; Nowlin, J.; Jones, R.; Practico, D.; Rokach, J.; Stackman, R. Chronic melatonin therapy fails to alter amyloid burden or oxidative damage in old Tg2576 mice: Implications for clinical trials. Brain Res. 2005, 1037, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Masilamoni, J.G.; Jesudason, E.P.; Dhandayuthapani, S.; Ashok, B.S.; Vignesh, S.; Jerabaj, C.E.; Paul, S.F.D.; Jayakumar, R. The neuroprotective role of melatonin against amyloid β peptide injected mice. Free Radic. Res. 2008, 42, 661–673. [Google Scholar] [CrossRef] [PubMed]
- Roh, J.H.; Jiang, H.; Finn, M.B.; Stewart, F.R.; Mahan, T.E.; Cirrito, J.R.; Heda, A.; Snider, B.J.; Li, M.; Yanagisawa, M.; et al. Potential role of orexin and sleep modulation in the pathogenesis of Alzheimer’s disease. J. Exp. Med. 2014, 211, 2487–2496. [Google Scholar] [CrossRef] [PubMed]
- Rothman, S.M.; Herdener, N.; Frankola, K.A.; Mughal, M.R.; Mattson, M.P. Chronic mild sleep restriction accentuates contextual memory impairments and accumulations of cortical Aβ and pTau in a mouse model of Alzheimer’s disease. Brain Res. 2013, 1529, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Di Meco, A.; Joshi, Y.B.; Pratico, D. Sleep deprivation impairs memory, tau metabolism, and synaptic integrity of a mouse model of Alzheimer’s disease with plaques and tangles. Neurobiol. Aging 2014, 35, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Wang, J.Z. Alzheimer-like tau phosphorylation induced by wortmannin in vivo and its attenuation by melatonin. Acta Pharmacol. Sin. 2002, 23, 183–187. [Google Scholar]
- Zhu, L.Q.; Wang, S.H.; Ling, Z.Q.; Wang, D.L.; Wang, J.Z. Effect of inhibiting melatonin biosynthesis on spatial memory retention and tau phosphorylation in the rat. J. Pineal Res. 2004, 37, 71–77. [Google Scholar] [CrossRef]
- Stenberg, D. Neuroanatomy and neurochemistry of sleep. Cell. Mol. Life Sci. 2007, 64, 1187–1204. [Google Scholar] [CrossRef]
- Watson, C.J.; Baghdoyan, H.A.; Lydic, R. Neuropharmacology of sleep and wakefulness. Sleep Med. Clin. 2010, 5, 513–528. [Google Scholar] [CrossRef]
- Cortese, B.M.; Mitchell, T.R.; Galloway, M.P.; Prevost, K.E.; Fang, J.; Moore, G.J.; Uhde, T.W. Region-specific alteration in brain glutamate: Possible relationship to risk-taking behavior. Physiol. Behav. 2010, 99, 445–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dash, M.B.; Douglas, C.L.; Vyazovskiy, V.V.; Cirelli, C.; Tononi, G. Long-term homeostasis of extracellular glutamate in the rat cerebral cortex across sleep and waking states. J. Neurosci. 2009, 29, 620–629. [Google Scholar] [CrossRef] [PubMed]
- Prince, T.M.; Abel, T. The impact of sleep loss on hippocampal function. Learn. Mem. 2013, 20, 558–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, M.; Yan, J.; He, C.; Yang, L.; Tan, G.; Li, C.; Hu, Z.; Wang, J. Short/term sleep deprivation impairs spatial working memory and modulates expression levels of ionotropic glutamate receptor subunits in the hippocampus. Behav. Brain Res. 2015, 286, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Li, C.; He, C.; Yang, L.; Tan, G.; Yan, J.; Wang, J.; Hu, Z. Short-term sleep deprivation disrupts the molecular composition of ionotropic glutamate receptors in the entorhinal cortex and impairs the rat spatial reference memory. Behav. Brain Res. 2016, 300, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Cirelli, C.; Tononi, G. Gene expression in the brain across the sleep-waking cycle. Brain Res. 2000, 885, 303–321. [Google Scholar] [CrossRef]
- Magnusson, K.R.; Nelson, S.E.; Young, A.B. Age-related changes in the protein expression of subunits of the NMDA receptor. Brain Res. Mol. Brain Res. 2002, 99, 40–45. [Google Scholar] [CrossRef]
- Liu, P.; Smith, P.F.; Darlington, C.L. Glutamate receptor subunits expression in memory-associated brain structures: Regional variations and effects of aging. Synapse 2008, 62, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Kristofikova, Z.; Vrajova, M.; Sirova, J.; Vales, K.; Petrasek, T.; Schonig, K.; Tews, B.; Schwab, M.; Bartsch, D.; Stuchlik, A.; et al. N-methyl-d-aspartate receptor—Nitric oxide synthase pathway in the cortex of NOGO-A-deficient rats in relation to brain laterality and schizophrenia. Front. Behav. Neurosci. 2013, 7, 90. [Google Scholar] [CrossRef]
- Bi, H.; Sze, C.I. N-methyl-d-aspartate receptor subunit NR2A and NR2B messenger RNA levels are altered in the hippocampus and entorhinal cortex in Alzheimer’s disease. J. Neurol. Sci. 2002, 200, 11–18. [Google Scholar] [CrossRef]
- Mishizen-Eberz, A.J.; Rissman, R.A.; Carter, T.L.; Ikonomovic, M.D.; Wolfe, B.B.; Armstrong, D.M. Biochemical and molecular studies of NMDA receptor subunits NR1/2A/2B in hippocampal subregions throughout the progression of Alzheimer disease pathology. Neurobiol. Dis. 2004, 15, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Hynd, M.R.; Scott, H.L.; Dodd, P.R. Differential expression of N-methyl-d-aspartate receptor NR2 isoforms in Alzheimer’s disease. J. Neurochem. 2004, 90, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Texido, L.; Martin-Satue, M.; Alberdi, E.; Solsona, C.; Matute, C. Amyloid β peptide oligomers directly activate NMDA receptors. Cell Calcium 2011, 49, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Ronicke, R.; Mikhaylova, M.; Ronicke, S.; Meinhardt, J.; Schroder, U.H.; Fandrich, M.; Reiser, G.; Kreutz, M.R.; Reymann, K.G. Early neuronal dysfunction by amyloid β oligomers depends on activation of NR2B-containing NMDA receptor. Neurobiol. Aging 2011, 32, 2219–2228. [Google Scholar] [CrossRef] [PubMed]
- Allyson, J.; Dontigny, E.; Auberson, Y.; Cyr, M.; Massicotte, G. Blockade of NR2A-containing NMDA receptors induces tau phosphorylation in rat hippocampal slices. Neural Plast. 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- Petrasek, T.; Skurlova, M.; Maleninska, K.; Vojtechova, I.; Kristofikova, Z.; Matuskova, H.; Sirova, J.; Vales, K.; Ripova, D.; Stuchlik, A. A rat model of Alzheimer’s disease based on Abeta42 and pro-oxidative substances exhibits cognitive deficit and alterations in glutamatergic and cholinergic neurotransmitter systems. Front. Aging Neurosci. 2016, 8, 83. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, Y.; Hirase, H.; Watanabe, M.; Itakura, M.; Takahashi, M.; Shigemoto, R. Left-right asymmetry of the hippocampal synapses with differential subunit allocation of glutamate receptors. Proc. Natl. Acad. Sci. USA 2008, 105, 19498–19503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristofikova, Z.; Ricny, J.; Ort, M. Aging and lateralization of the rat brain on a biochemical level. Neurochem. Res. 2010, 35, 1138–1146. [Google Scholar] [CrossRef]
- Kalinchuk, A.V.; Stenberg, D.; Rosenberg, P.A.; Porkka-Heiskanen, T. Inducible, and neuronal nitric oxide synthases (NOS) have complementary roles in recovery sleep induction. Eur. J. Neurosci. 2006, 24, 1443–1456. [Google Scholar] [CrossRef]
- Kalinchuk, A.V.; McCarley, R.W.; Porkka-Heiskanen, T.; Basheer, R. Sleep deprivation triggers inducible nitric oxide-dependent nitric oxide production in wake-active basal forebrain neurons. J. Neurosci. 2010, 30, 13254–13264. [Google Scholar] [CrossRef]
- Kalinchuk, A.V.; Porkka-Heiskanen, T.; McCarley, R.W.; Basheer, R. Cholinergic neurons of the basal forebrain mediate biochemical and electrophysiological mechanisms underlaying sleep homeostasis. Eur. J. Neurosci. 2015, 41, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Cespuglio, R.; Amrouni, D.; Meiller, A.; Buguet, A.; Gautier-Sauvigne, S. Nitric oxide in the regulation of the sleep-wake states. Sleep Med. Rev. 2012, 16, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Sauvet, F.; Florence, G.; Van Beers, P.; Drogou, C.; Lagrume, C.; Chaumes, C.; Ciret, S.; Leftheriotis, G.; Chennaoui, M. Total sleep deprivation alters endothelial function in rats: A nonsympathetic mechanism. Sleep 2014, 37, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Rytkonen, K.M.; Wigren, H.K.; Kostin, A.; Porkka-Heiskanen, T.; Kalinchuk, A.V. Nitric oxide-mediated recovery sleep is attenuated with aging. Neurobiol. Aging 2010, 31, 2011–2019. [Google Scholar] [CrossRef] [PubMed]
- Kristofikova, Z.; Kozmikova, I.; Hovorkova, P.; Ricny, J.; Zach, P.; Majer, E.; Klaschka, J.; Ripova, D. Lateralization of hippocampal nitric oxide mediator system in people with Alzheimer disease, multi-infarct dementia and schizophrenia. Neurochem. Int. 2008, 53, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Kristofikova, Z.; Stastny, F.; Bubenikova, V.; Druga, R.; Klaschka, J.; Spaniel, F. Age- and sex-dependent laterality of rat hippocampal cholinergic system in relation to animal models of neurodevelopmental and neurodegenerative disorders. Neurochem. Res. 2004, 29, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Perusquia, M.; Greenway, C.D.; Perkins, L.M.; Stallone, J.N. Systemic hypotensive effects of testosterone are androgen structure-specific and neuronal nitric oxide synthase-dependent. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, 189–195. [Google Scholar] [CrossRef]
- Moreau, K.L. Modulatory influence of sex hormones on vascular aging. Am. J. Physiol. Heart Circ. Physiol. 2019. [Google Scholar] [CrossRef]
- Ganguly, P.; Holland, F.H.; Brenhouse, H.C. Functional uncoupling NMDAR NR2A subunit from PSD-95 in the prefrontal cortex: Effects on behavioral dysfunction and parvalbumin loss after early-life stress. Neuropsychopharmacology 2015, 40, 2666–2675. [Google Scholar] [CrossRef]
- Martisova, E.; Solas, M.; Horrillo, I.; Ortega, J.E.; Meana, J.J.; Tordera, R.M.; Ramirez, M.J. Long-lasting effects of early-life stress on glutamatergic/GABAergic circuitry in the rat hippocampus. Neuropharmacology 2012, 62, 1944–1953. [Google Scholar] [CrossRef]
- Chauhan, P.S.; Misra, U.K.; Kalita, J. A study of glutamate levels, NR1, NR2A, NR2B receptors and oxidative stress in a rat model of Japanese encephalitis. Physiol. Behav. 2017, 171, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Costa-Nunes, J.; Zubareva, O.; Araujo-Correia, M.; Valenca, A.; Schroeter, C.A.; Pawluski, J.L.; Vignisse, J.; Steinbusch, H.; Hermes, D.; Phillipines, M.; et al. Altered emotionality, hippocampus-dependent performance and expression of NMDA receptor subunit mRNAs in chronically stressed mice. Stress 2014, 17, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Christie, M.A.; McKenna, J.T.; Connolly, N.P.; McCarley, R.W.; Strecker, R.E. 24 h of sleep deprivation in the rat increases sleepiness and decreases vigilance: Introduction of the rat-psychomotor vigilance task. J. Sleep Res. 2008, 17, 376–384. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Index of Laterality for nNOS | Index of Laterality for eNOS | Index of Laterality for iNOS |
|---|---|---|---|
| Experiment I | |||
| young adult controls (n = 8) | −0.034 ± 0.033 | +0.188 ± 0.077 | +0.196 ± 0.235 |
| young adult exposed to FL (n = 10) | −0.072 ± 0.082 | +0.020 ± 0.094 | −0.128 ± 0.232 |
| old controls (n = 8) | +0.067 ± 0.065 | +0.156 ± 0.064 | −0.239 ± 0.226 |
| old exposed to FL (n = 9) | −0.007 ± 0.040 | −0.029 ± 0.102 | +0.084 ± 0.306 |
| ANOVA: | p = 0.448 | p = 0.266 | p = 0.637 |
| Experiment II | |||
| young adult controls (n = 12) | −0.064 ± 0.033 | +0.035 ± 0.088 | −0.163 ± 0.270 |
| young adult exposed to acute SD (n = 12) | −0.113 ± 0.055 | +0.093 ± 0.098 | −0.282 ± 0.249 |
| old controls (n = 12) | −0.136 ± 0.058 | −0.054 ± 0.061 | −0.385 ± 0.137 |
| old exposed to acute SD (n = 12) | −0.077 ± 0.092 | −0.007 ± 0.161 | −0.396 ± 0.184 |
| ANOVA: | p = 0.846 | p = 0.800 | p = 0.860 |
| Experiment III | |||
| young adult controls (n = 12) | −0.113 ± 0.046 | +0.005 ± 0.052 | −0.180 ± 0.229 |
| young adult exposed to chronic SD (n = 12) | −0.128 ± 0.038 | −0.011 ± 0.072 | −0.263 ± 0.229 |
| old controls (n = 12) | +0.007 ± 0.034 | +0.044 ± 0.074 | −0.131 ± 0.179 |
| old exposed to chronic SD (n = 12) | −0.073 ± 0.046 | +0.057 ± 0.071 | −0.137 ± 0.231 |
| ANOVA: | p = 0.109 | p = 0.879 | p = 0.971 |
| Components of NMDA—NO Pathway | Young Adult Controls (n = 32) | Old Controls (n = 32) | ANOVA: p |
|---|---|---|---|
| NR1 expression | 1.034 ± 0.008 | 0.971 ± 0.004 | <0.001 *** |
| NR2A expression | 1.026 ± 0.004 | 0.988 ± 0.009 | <0.001 *** |
| NR2B expression | 1.000 ± 0.005 | 0.969 ± 0.005 | <0.001 *** |
| nNOS activity in the R side | 423.3 ± 26.2 | 313.6 ± 29.3 | =0.007 ** |
| nNOS activity in the L side | 361.6 ± 22.0 | 294.9 ± 28.8 | =0.071 |
| eNOS activity in the R side | 155.9 ± 17.0 | 97.2 ± 11.1 | =0.005 ** |
| eNOS activity in the L side | 172.8 ± 18.1 | 109.5 ± 13.4 | =0.007 ** |
| iNOS activity in the R side | 9.2 ± 2.2 | 8.5 ± 1.5 | =0.800 |
| iNOS activity in the L side | 9.5 ± 2.2 | 6.0 ± 2.3 | =0.272 |
- nNOS: aging—F(1,62) = 6.23, p = 0.015, laterality—F(1,62) = 8.87, p = 0.004, interaction—F(1,62) = 2.54, p = 0.116.
- eNOS: aging—F(1,62) = 9.23, p = 0.004, laterality—F(1,62) = 3.79, p = 0.056, interaction—F(1,62) = 0.09, p = 0.761.
- iNOS: aging—F(1,62) = 0.99, p = 0.325, laterality—F(1,62) = 0.30, p = 0.589, interaction—F(1,62) = 0.51, p = 0.477.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kristofikova, Z.; Sirova, J.; Klaschka, J.; Ovsepian, S.V. Acute and Chronic Sleep Deprivation-Related Changes in N-methyl-D-aspartate Receptor—Nitric Oxide Signalling in the Rat Cerebral Cortex with Reference to Aging and Brain Lateralization. Int. J. Mol. Sci. 2019, 20, 3273. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133273
Kristofikova Z, Sirova J, Klaschka J, Ovsepian SV. Acute and Chronic Sleep Deprivation-Related Changes in N-methyl-D-aspartate Receptor—Nitric Oxide Signalling in the Rat Cerebral Cortex with Reference to Aging and Brain Lateralization. International Journal of Molecular Sciences. 2019; 20(13):3273. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133273
Chicago/Turabian StyleKristofikova, Zdenka, Jana Sirova, Jan Klaschka, and Saak V. Ovsepian. 2019. "Acute and Chronic Sleep Deprivation-Related Changes in N-methyl-D-aspartate Receptor—Nitric Oxide Signalling in the Rat Cerebral Cortex with Reference to Aging and Brain Lateralization" International Journal of Molecular Sciences 20, no. 13: 3273. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133273