Improving the Clinical Application of Natural Killer Cells by Modulating Signals Signal from Target Cells

 ,
,
Abstract
:1. Introduction
2. Results
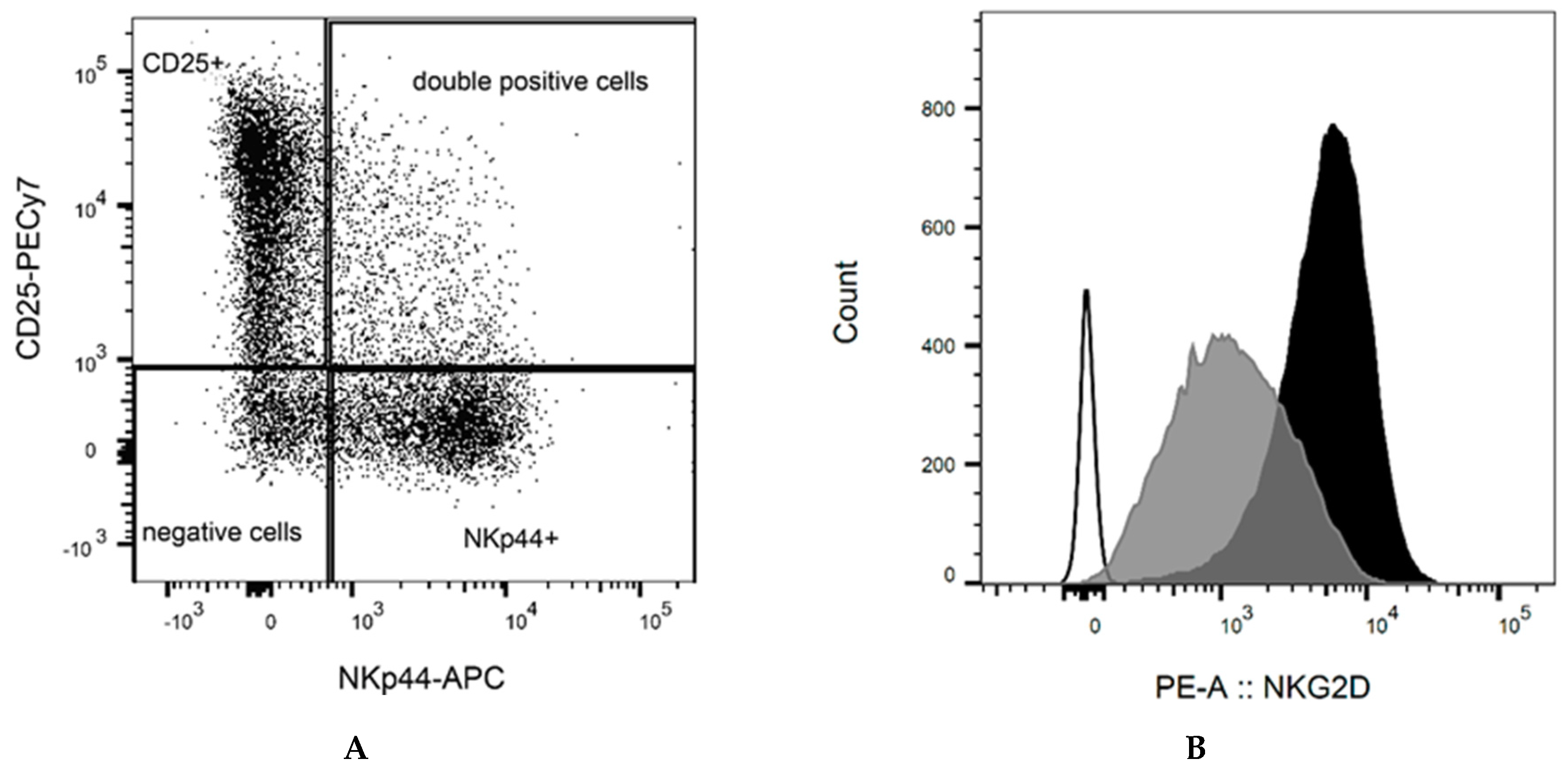
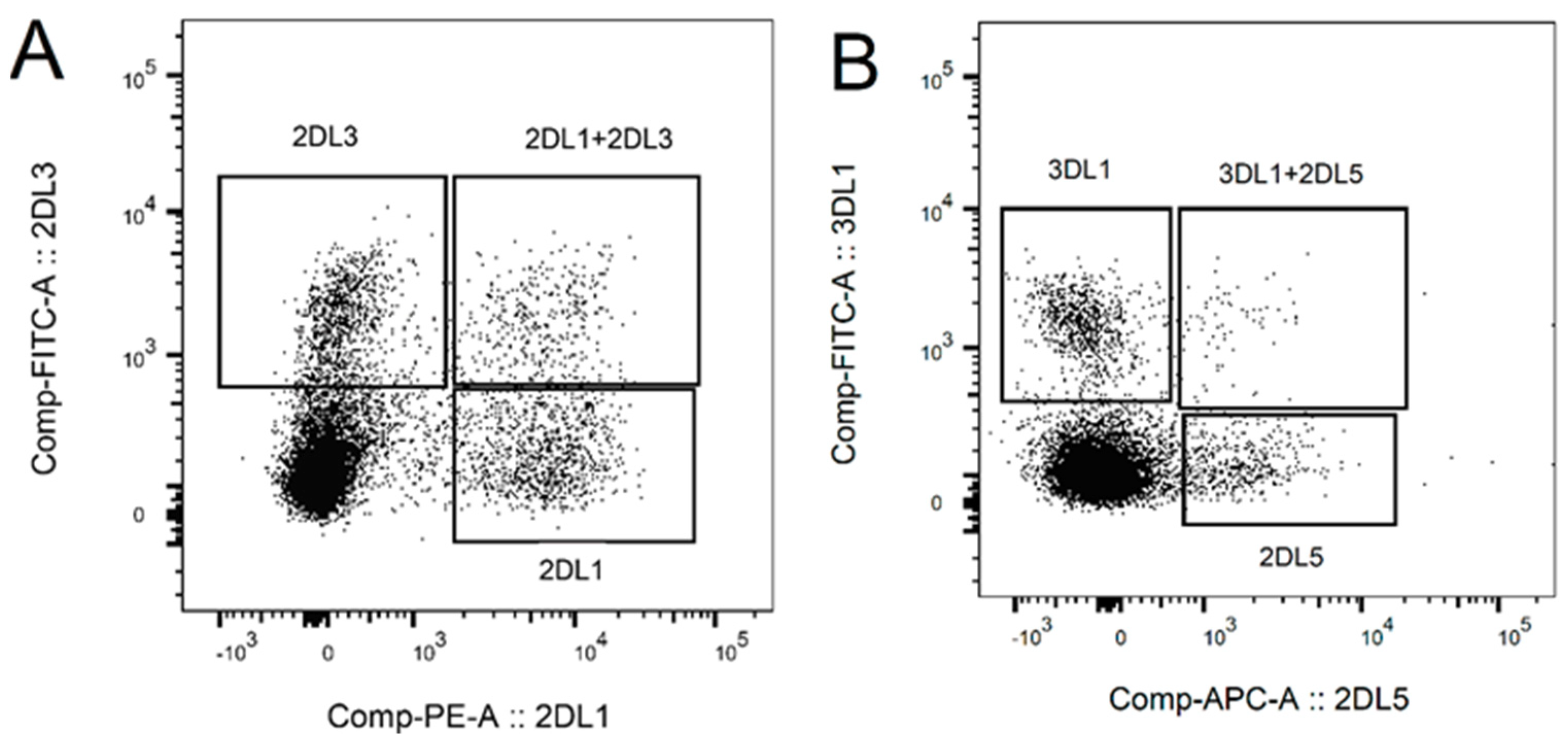
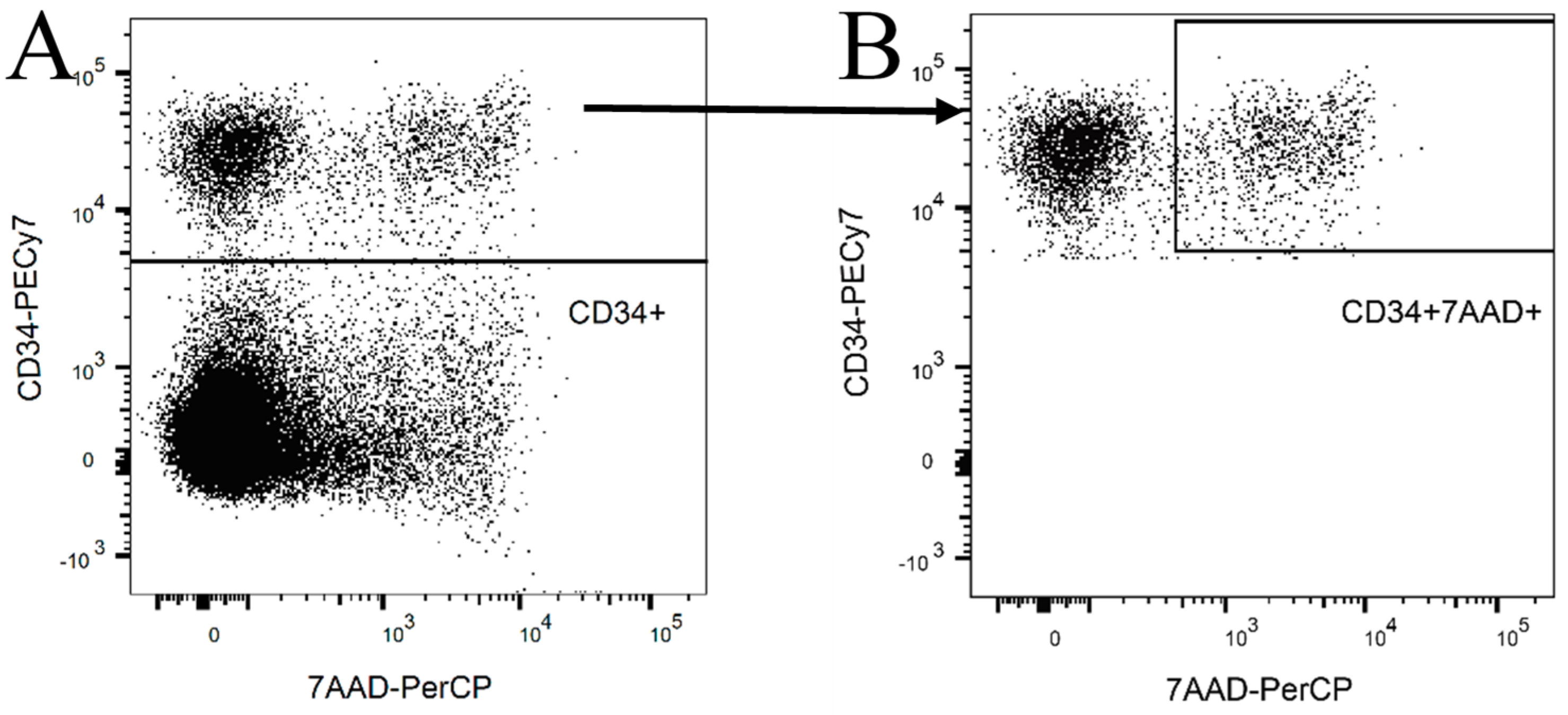
2.1. NK Cell Preparation
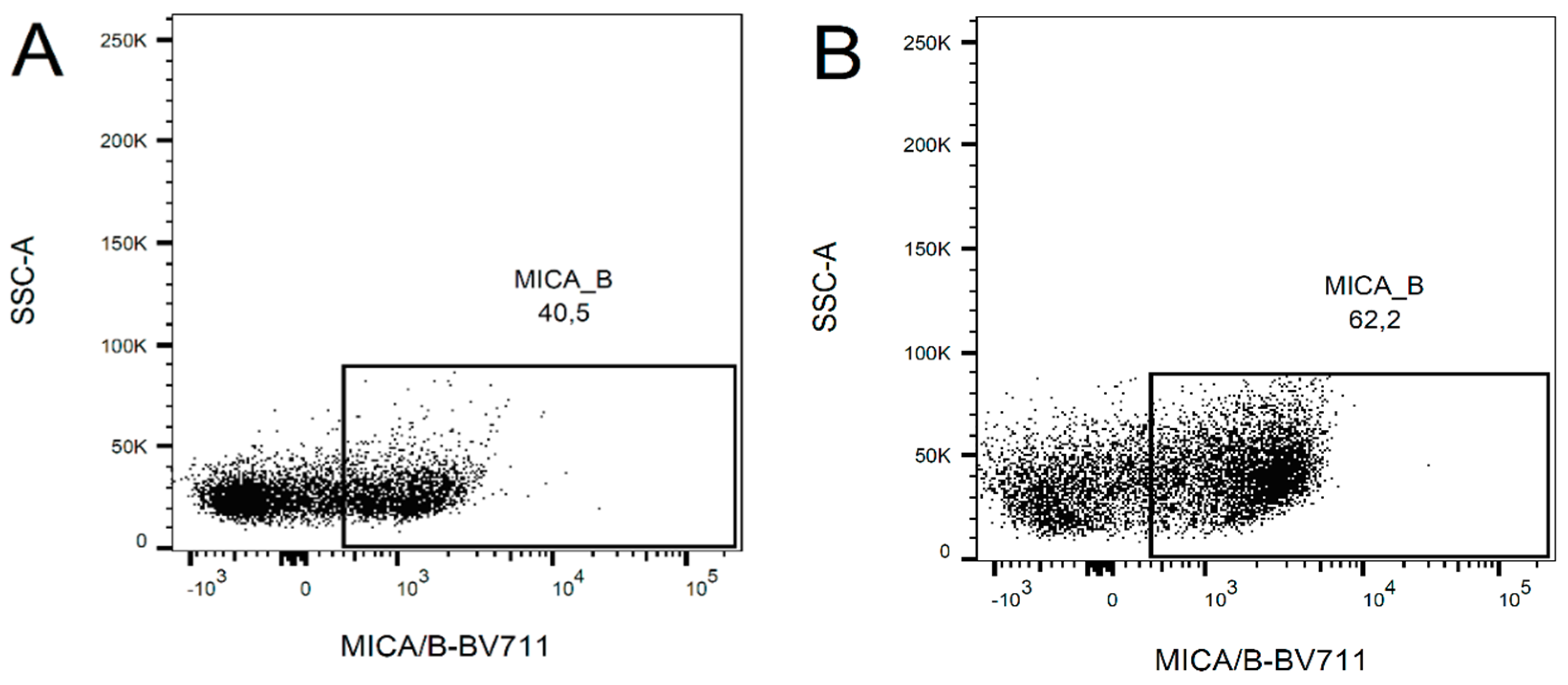
2.2. Change in the Expression of NKG2D Ligands after Ara-C Application
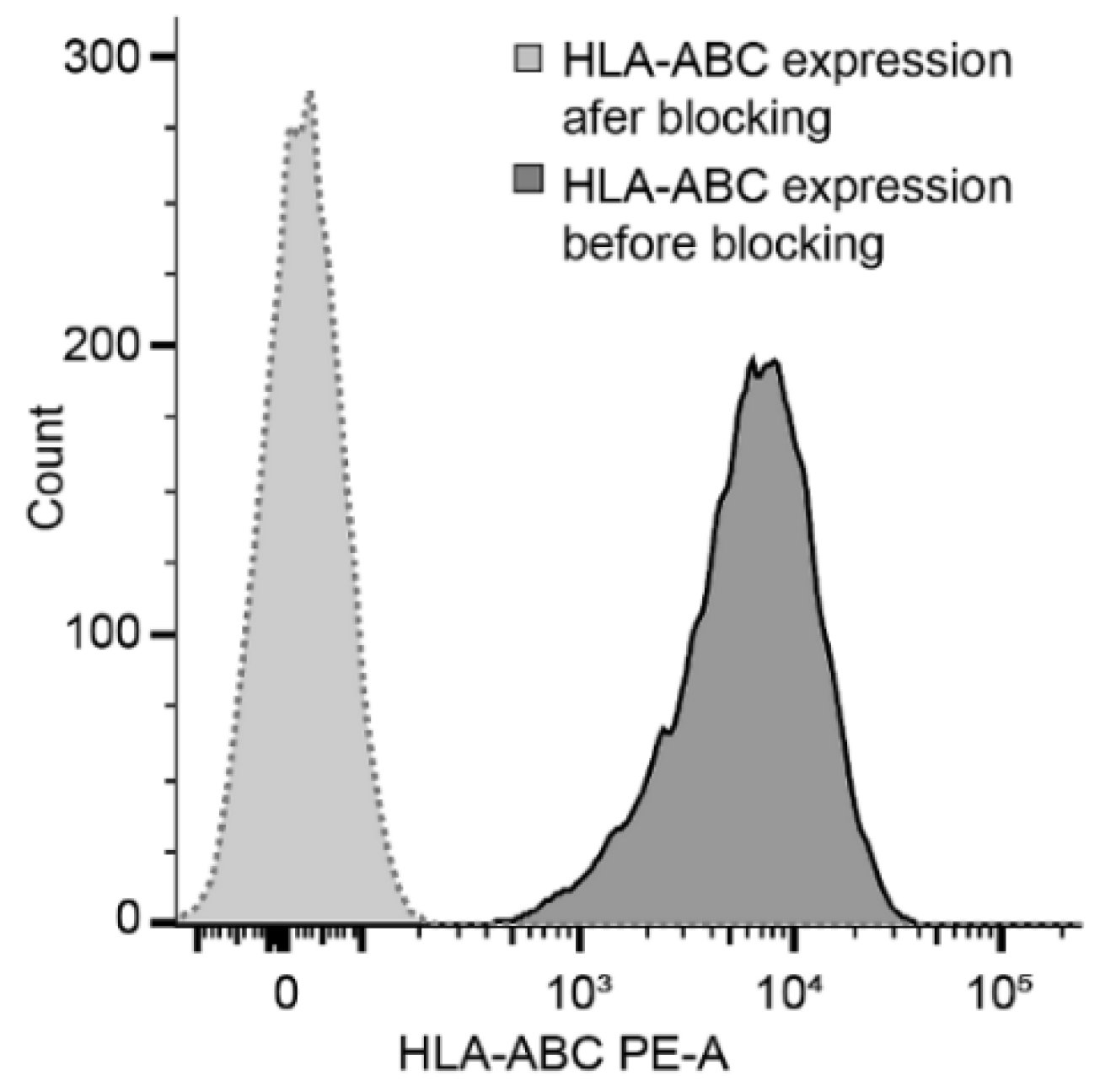
2.3. Target Cell Preparation and Cytotoxic Potential of NK Cells
3. Discussion
4. Methods
4.1. NK Cell Preparation
4.2. Target Cells Preparation
4.3. Flow Cytometry Detection of Surface Markers and Cytotoxicity
4.4. Evaluation of NKG2D Ligand Expression—qRT PCR
4.5. Co-Culture Cytotoxic Assay
4.6. Data Evaluation
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fathi, A.T.; Chen, Y.B. Treatment of relapse of acute myeloid leukemia after allogeneic hematopoietic stem cell transplantation. Curr. Hematol. Malig Rep. 2014, 9, 186–192. [Google Scholar] [CrossRef]
- Bejanyan, N.; Weisdorf, D.J.; Logan, B.R.; Wang, H.L.; Devine, S.M.; de Lima, M.; Bunjes, D.W.; Zhang, M.J. Survival of patients with acute myeloid leukemia relapsing after allogeneic hematopoietic cell transplantation: A center for international blood and marrow transplant research study. Biol. Blood Marrow Transplant. 2015, 21, 454–459. [Google Scholar] [CrossRef]
- Tasian, S.K. Acute myeloid leukemia chimeric antigen receptor T-cell immunotherapy: How far up the road have we traveled? Ther. Adv. Hematol. 2018, 9, 135–148. [Google Scholar] [CrossRef]
- Biassoni, R.; Cantoni, C.; Pende, D.; Sivori, S.; Parolini, S.; Vitale, M.; Bottino, C.; Moretta, A. Human natural killer cell receptors and co-receptors. Immunol. Rev. 2001, 181, 203–214. [Google Scholar] [CrossRef]
- Stebbins, C.C.; Watzl, C.; Billadeau, D.D.; Leibson, P.J.; Burshtyn, D.N.; Long, E.O. Vav1 dephosphorylation by the tyrosine phosphatase SHP-1 as a mechanism for inhibition of cellular cytotoxicity. Mol. Cell Biol. 2003, 23, 6291–6299. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Cherwinski, H.; Spies, T.; Phillips, J.H.; Lanier, L.L. DAP10 and DAP12 form distinct, but functionally cooperative, receptor complexes in natural killer cells. J. Exp. Med. 2000, 192, 1059–1068. [Google Scholar] [CrossRef]
- Garrido, F.; Romero, I.; Aptsiauri, N.; Garcia-Lora, A.M. Generation of MHC class I diversity in primary tumors and selection of the malignant phenotype. Int. J. Cancer 2016, 138, 271–280. [Google Scholar] [CrossRef]
- Vey, N.; Karlin, L.; Sadot-Lebouvier, S.; Broussais, F.; Berton-Rigaud, D.; Rey, J.; Charbonnier, A.; Marie, D.; Andre, P.; Paturel, C.; et al. A phase 1 study of lirilumab (antibody against killer immunoglobulin-like receptor antibody KIR2D; IPH2102) in patients with solid tumors and hematologic malignancies. Oncotarget 2018, 9, 17675–17688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, T.; Hartl, F.U.; Wieland, F. CD40, an extracellular receptor for binding and uptake of Hsp70-peptide complexes. J. Cell Biol. 2002, 158, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Binder, R.J.; Han, D.K.; Srivastava, P.K. CD91: A receptor for heat shock protein gp96. Nat. Immunol. 2000, 1, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Cerboni, C.; Fionda, C.; Soriani, A.; Zingoni, A.; Doria, M.; Cippitelli, M.; Santoni, A. The DNA Damage Response: A Common Pathway in the Regulation of NKG2D and DNAM-1 Ligand Expression in Normal, Infected, and Cancer Cells. Front. Immunol. 2014, 4, 508. [Google Scholar] [CrossRef] [PubMed]
- Poggi, A.; Catellani, S.; Garuti, A.; Pierri, I.; Gobbi, M.; Zocchi, M.R. Effective in vivo induction of NKG2D ligands in acute myeloid leukaemias by all-trans-retinoic acid or sodium valproate. Leukemia 2009, 23, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Son, Y.O.; Park, S.W.; Bae, J.H.; Chung, J.S.; Kim, H.H.; Chung, B.S.; Kim, S.H.; Kang, C.D. Increase of NKG2D ligands and sensitivity to NK cell-mediated cytotoxicity of tumor cells by heat shock and ionizing radiation. Exp. Mol. Med. 2006, 38, 474–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morisaki, T.; Onishi, H.; Katano, M. Cancer immunotherapy using NKG2D and DNAM-1 systems. Anticancer Res. 2012, 32, 2241–2247. [Google Scholar] [PubMed]
- Liu, Y.; Zeng, G. Cancer and innate immune system interactions: Translational potentials for cancer immunotherapy. J. Immunother. 2012, 35, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guo, J.R.; Chen, Q.Q.; Wang, C.Y.; Zhang, W.J.; Yao, M.C.; Zhang, W. Exploring the Antitumor Mechanism of High-Dose Cytarabine through the Metabolic Perturbations of Ribonucleotide and Deoxyribonucleotide in Human Promyelocytic Leukemia HL-60 Cells. Molecules 2017, 22, 499. [Google Scholar] [CrossRef] [PubMed]
- Laverdiere, I.; Boileau, M.; Neumann, A.L.; Frison, H.; Mitchell, A.; Ng, S.W.K.; Wang, J.C.Y.; Minden, M.D.; Eppert, K. Leukemic stem cell signatures identify novel therapeutics targeting acute myeloid leukemia. Blood Cancer J. 2018, 8, 52. [Google Scholar] [CrossRef]
- Chavez-Blanco, A.; De la Cruz-Hernandez, E.; Dominguez, G.I.; Rodriguez-Cortez, O.; Alatorre, B.; Perez-Cardenas, E.; Chacon-Salinas, R.; Trejo-Becerril, C.; Taja-Chayeb, L.; Trujillo, J.E.; et al. Upregulation of NKG2D ligands and enhanced natural killer cell cytotoxicity by hydralazine and valproate. Int. J. Oncol. 2011, 39, 1491–1499. [Google Scholar]
- Nanbakhsh, A.; Pochon, C.; Mallavialle, A.; Amsellem, S.; Bourhis, J.H.; Chouaib, S. c-Myc regulates expression of NKG2D ligands ULBP1/2/3 in AML and modulates their susceptibility to NK-mediated lysis. Blood 2014, 123, 3585–3595. [Google Scholar] [CrossRef] [Green Version]
- Tajima, F.; Kawatani, T.; Endo, A.; Kawasaki, H. Natural killer cell activity and cytokine production as prognostic factors in adult acute leukemia. Leukemia 1996, 10, 478–482. [Google Scholar]
- Farag, S.S.; Caligiuri, M.A. Immunologic approaches to acute leukemia in the elderly. Semin. Hematol. 2006, 43, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Parisi, S.; Lecciso, M.; Ocadlikova, D.; Salvestrini, V.; Ciciarello, M.; Forte, D.; Corradi, G.; Cavo, M.; Curti, A. The More, The Better: “Do the Right Thing” For Natural Killer Immunotherapy in Acute Myeloid Leukemia. Front. Immunol. 2017, 8, 1330. [Google Scholar] [CrossRef]
- Holubova, M.; Miklikova, M.; Leba, M.; Georgiev, D.; Jindra, P.; Caprnda, M.; Ciccocioppo, R.; Kruzliak, P.; Lysak, D. Cryopreserved NK cells in the treatment of hematological malignancies: Preclinical study. J. Cancer Res. Clin. Oncol. 2016, 142, 2561–2567. [Google Scholar] [CrossRef] [PubMed]
- Clausen, J.; Vergeiner, B.; Enk, M.; Petzer, A.L.; Gastl, G.; Gunsilius, E. Functional significance of the activation-associated receptors CD25 and CD69 on human NK-cells and NK-like T-cells. Immunobiology 2003, 207, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Berg, M.; Lundqvist, A.; McCoy, P., Jr.; Samsel, L.; Fan, Y.; Tawab, A.; Childs, R. Clinical-grade ex vivo-expanded human natural killer cells up-regulate activating receptors and death receptor ligands and have enhanced cytolytic activity against tumor cells. Cytotherapy 2009, 11, 341–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastaglio, S.; Wong, E.; Perera, T.; Ripley, J.; Blombery, P.; Smyth, M.J.; Koldej, R.; Ritchie, D. Natural killer receptor ligand expression on acute myeloid leukemia impacts survival and relapse after chemotherapy. Blood Adv. 2018, 2, 335–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diermayr, S.; Himmelreich, H.; Durovic, B.; Mathys-Schneeberger, A.; Siegler, U.; Langenkamp, U.; Hofsteenge, J.; Gratwohl, A.; Tichelli, A.; Paluszewska, M.; et al. NKG2D ligand expression in AML increases in response to HDAC inhibitor valproic acid and contributes to allorecognition by NK-cell lines with single KIR-HLA class I specificities. Blood 2008, 111, 1428–1436. [Google Scholar] [CrossRef] [PubMed]
- Ogbomo, H.; Michaelis, M.; Klassert, D.; Doerr, H.W.; Cinatl, J., Jr. Resistance to cytarabine induces the up-regulation of NKG2D ligands and enhances natural killer cell lysis of leukemic cells. Neoplasia 2008, 10, 1402–1410. [Google Scholar] [CrossRef]
- Niu, C.; Jin, H.; Li, M.; Zhu, S.; Zhou, L.; Jin, F.; Zhou, Y.; Xu, D.; Xu, J.; Zhao, L.; et al. Low-dose bortezomib increases the expression of NKG2D and DNAM-1 ligands and enhances induced NK and gammadelta T cell-mediated lysis in multiple myeloma. Oncotarget 2017, 8, 5954–5964. [Google Scholar] [CrossRef]
- He, S.; Yin, T.; Li, D.; Gao, X.; Wan, Y.; Ma, X.; Ye, T.; Guo, F.; Sun, J.; Lin, Z.; et al. Enhanced interaction between natural killer cells and lung cancer cells: Involvement in gefitinib-mediated immunoregulation. J. Transl. Med. 2013, 11, 186. [Google Scholar] [CrossRef]
- Gao, M.; Gao, L.; Yang, G.; Tao, Y.; Hou, J.; Xu, H.; Hu, X.; Han, Y.; Zhang, Q.; Zhan, F.; et al. Myeloma cells resistance to NK cell lysis mainly involves an HLA class I-dependent mechanism. Acta Biochim. Biophys. Sin. (Shanghai) 2014, 46, 597–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leone, P.; De Re, V.; Vacca, A.; Dammacco, F.; Racanelli, V. Cancer treatment and the KIR-HLA system: An overview. Clin. Exp. Med. 2017, 17, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Godal, R.; Bachanova, V.; Gleason, M.; McCullar, V.; Yun, G.H.; Cooley, S.; Verneris, M.R.; McGlave, P.B.; Miller, J.S. Natural killer cell killing of acute myelogenous leukemia and acute lymphoblastic leukemia blasts by killer cell immunoglobulin-like receptor-negative natural killer cells after NKG2A and LIR-1 blockade. Biol. Blood Marrow Transplant. 2010, 16, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Vey, N.; Bourhis, J.H.; Boissel, N.; Bordessoule, D.; Prebet, T.; Charbonnier, A.; Etienne, A.; Andre, P.; Romagne, F.; Benson, D.; et al. A phase 1 trial of the anti-inhibitory KIR mAb IPH2101 for AML in complete remission. Blood 2012, 120, 4317–4323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohrt, H.E.; Thielens, A.; Marabelle, A.; Sagiv-Barfi, I.; Sola, C.; Chanuc, F.; Fuseri, N.; Bonnafous, C.; Czerwinski, D.; Rajapaksa, A.; et al. Anti-KIR antibody enhancement of anti-lymphoma activity of natural killer cells as monotherapy and in combination with anti-CD20 antibodies. Blood 2014, 123, 678–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holubova, M.; Houdova, L.; Jindra, P.; Lysak, D. Expression of inhibitory KIRs after hematopoietic stem cells transplantation. In Proceedings of the Analytical Cytometry IX, Prague, Czech Republic, 14–17 October 2017; Tomáš, K., Ed.; AMCA: Prague, Czech Republic, 2017. Abstract number pCT-7. p. 111. [Google Scholar]
- Bachanova, V.; Cooley, S.; Defor, T.E.; Verneris, M.R.; Zhang, B.; McKenna, D.H.; Curtsinger, J.; Panoskaltsis-Mortari, A.; Lewis, D.; Hippen, K.; et al. Clearance of acute myeloid leukemia by haploidentical natural killer cells is improved using IL-2 diphtheria toxin fusion protein. Blood 2014, 123, 3855–3863. [Google Scholar] [CrossRef]
- Mavers, M.; Bertaina, A. High-Risk Leukemia: Past, Present, and Future Role of NK Cells. J. Immunol Res. 2018, 2018, 1586905. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Name | Sequence 5′→3′ |
|---|---|---|
| MICA | MICA-F | CTTGGCCATGAACGTCAGG |
| MICA-R | CCTCTGAGGCCTCGCTGCG | |
| MICB | MICB-F | ACCTTGGCTATGAACGTCACA |
| MICB-R | CCCTCTGAGACCTCGCTGCA | |
| ULBP1 | ULBP1-F | GTACTGGGAACAAATGCTGGAT |
| ULBP1-R | AACTCTCCTCATCTGCCAGCT | |
| ULBP2 | ULBP2-F | TTACTTCTCAATGGGAGACTGT |
| ULBP2-R | TGTGCCTGAGGACATGGCGA | |
| ULBP3 | ULBP3-F | CTGATGCACAGGAAGAAGAG |
| ULBP3-R | TATGGCTTTGGGTTGAGCTAAG | |
| B2M | B2M-F | CTATCCAGCGTACTCCAAAG |
| B2M-R | GAAAGACCAGTCCTTGCTGA |
| Ara-C before Addition to NK Cells | HLA-Blocking before Addition to NK Cells | Co-Culture with NK Cells | |
|---|---|---|---|
| T1 | 24 h | 24 h | 8 h |
| T2 | 24 h | 24 h | 24 h |
| T3 | 48 h | 24 h | 8 h |
| T4 | 48 h | 24 h | 24 h |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holubova, M.; Leba, M.; Gmucova, H.; Caputo, V.S.; Jindra, P.; Lysak, D. Improving the Clinical Application of Natural Killer Cells by Modulating Signals Signal from Target Cells. Int. J. Mol. Sci. 2019, 20, 3472. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143472
Holubova M, Leba M, Gmucova H, Caputo VS, Jindra P, Lysak D. Improving the Clinical Application of Natural Killer Cells by Modulating Signals Signal from Target Cells. International Journal of Molecular Sciences. 2019; 20(14):3472. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143472
Chicago/Turabian StyleHolubova, Monika, Martin Leba, Hana Gmucova, Valentina S. Caputo, Pavel Jindra, and Daniel Lysak. 2019. "Improving the Clinical Application of Natural Killer Cells by Modulating Signals Signal from Target Cells" International Journal of Molecular Sciences 20, no. 14: 3472. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143472