Annona atemoya Leaf Extract Improves Scopolamine-Induced Memory Impairment by Preventing Hippocampal Cholinergic Dysfunction and Neuronal Cell Death


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
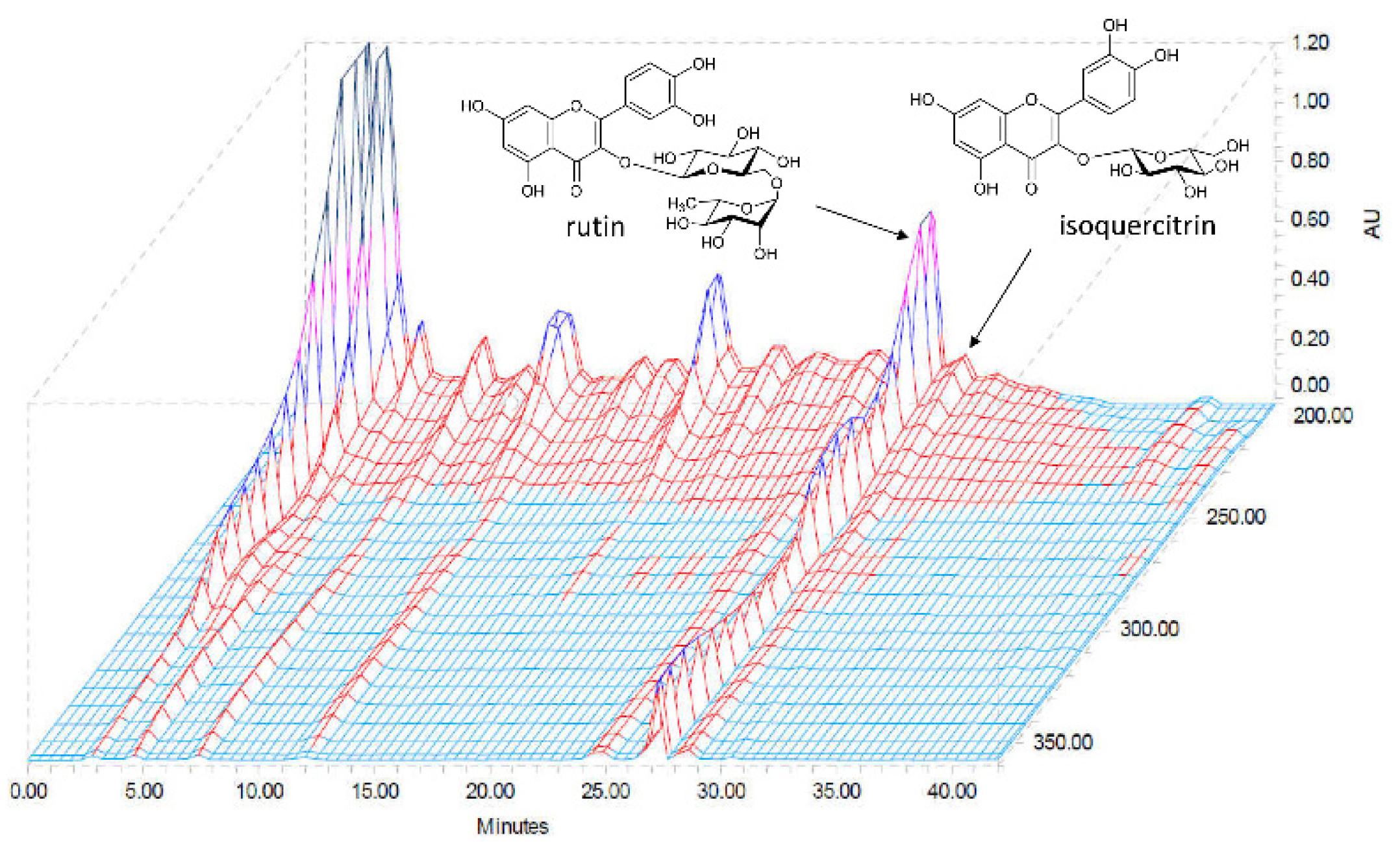
2.1. High-Performance Liquid Chromatography (HPLC) Determination of the Two Standard Compounds in AAL
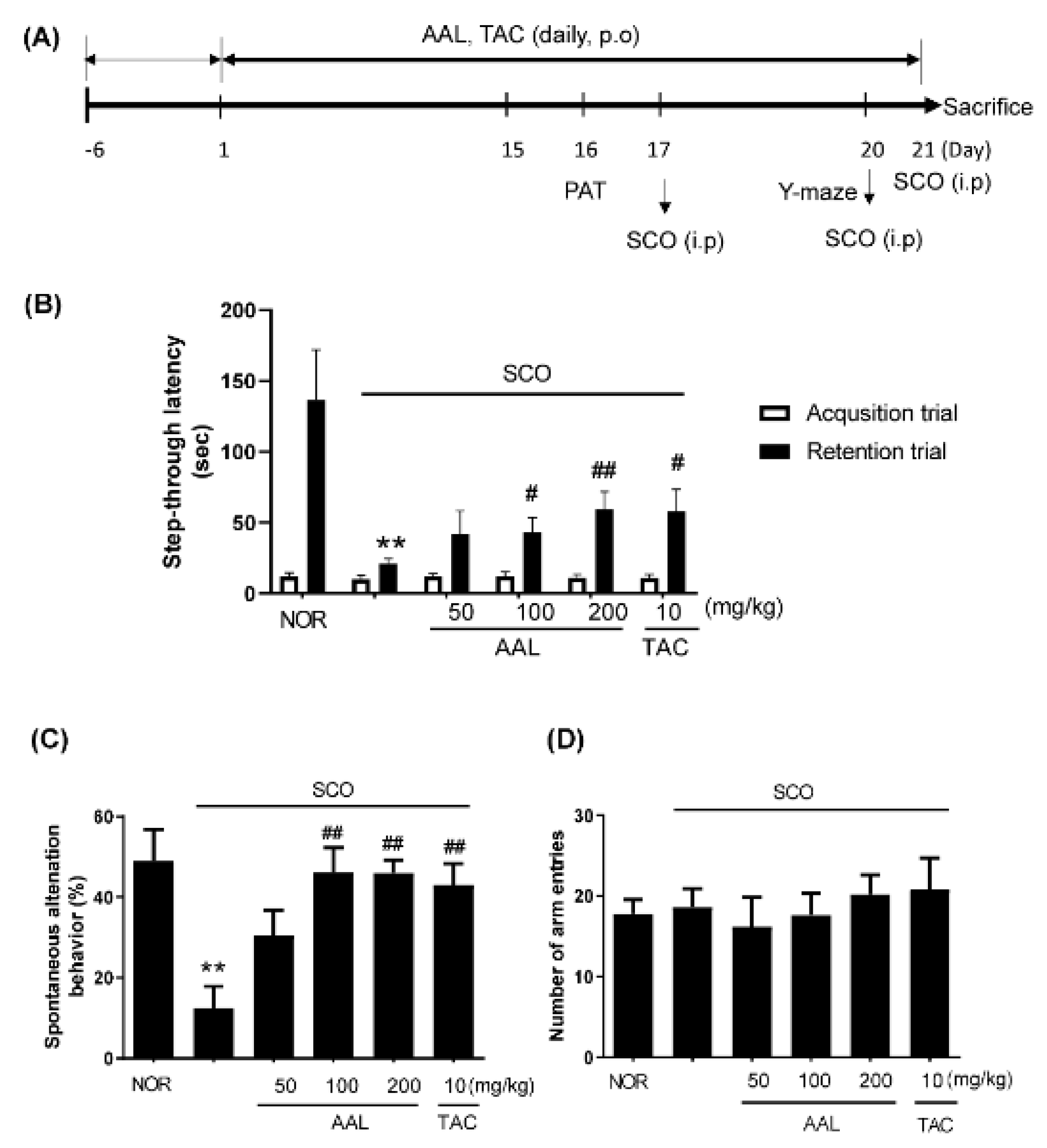
2.2. Effect of AAL Extract on Memory Deficit in SCO-Treated Mice
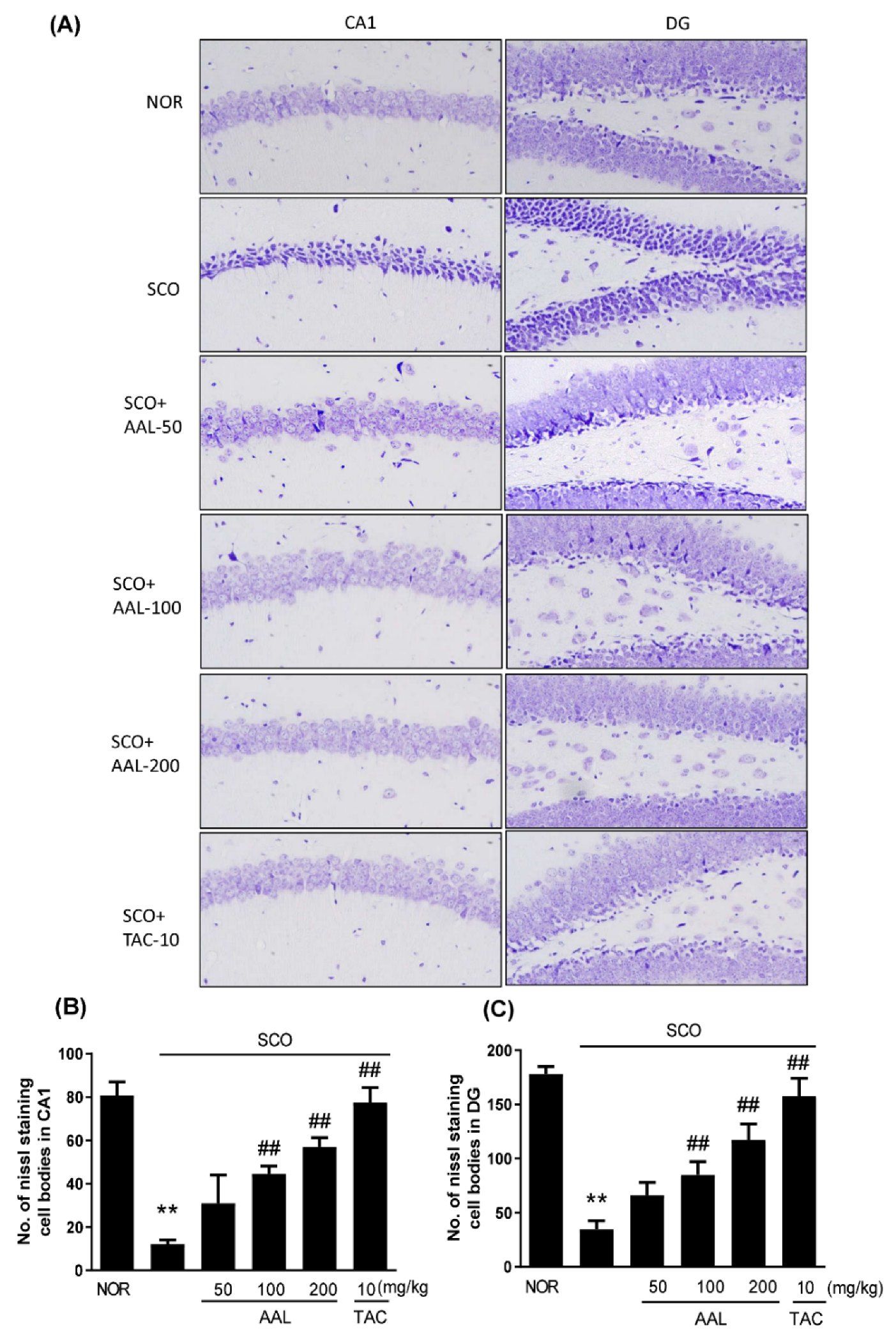
2.3. Effect of AAL Extract on Neuronal Cell Loss in SCO-Treated Mice
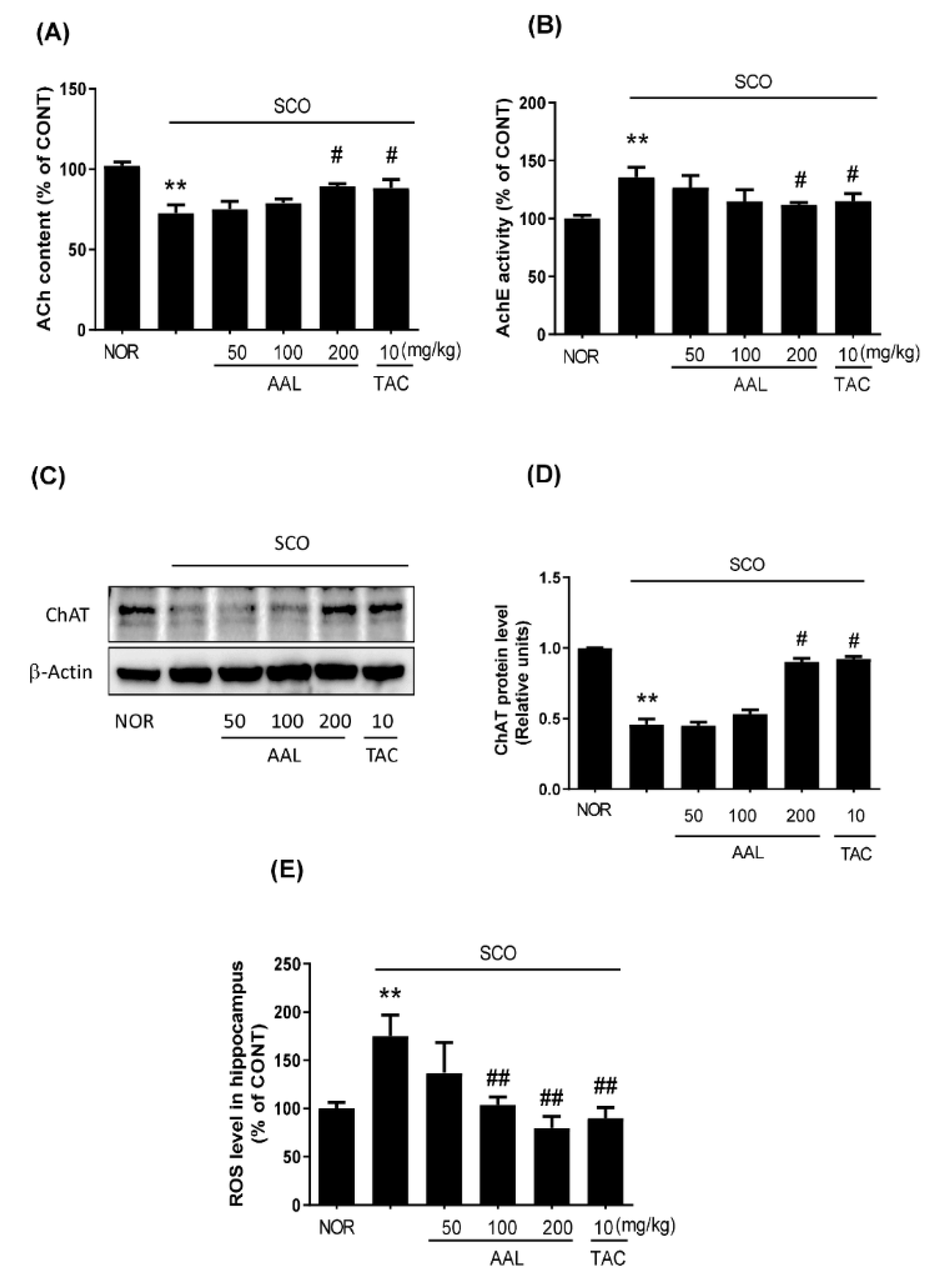
2.4. Effect of AAL Extract on Cholinergic System and Oxidative Stress in SCO-Treated Mice
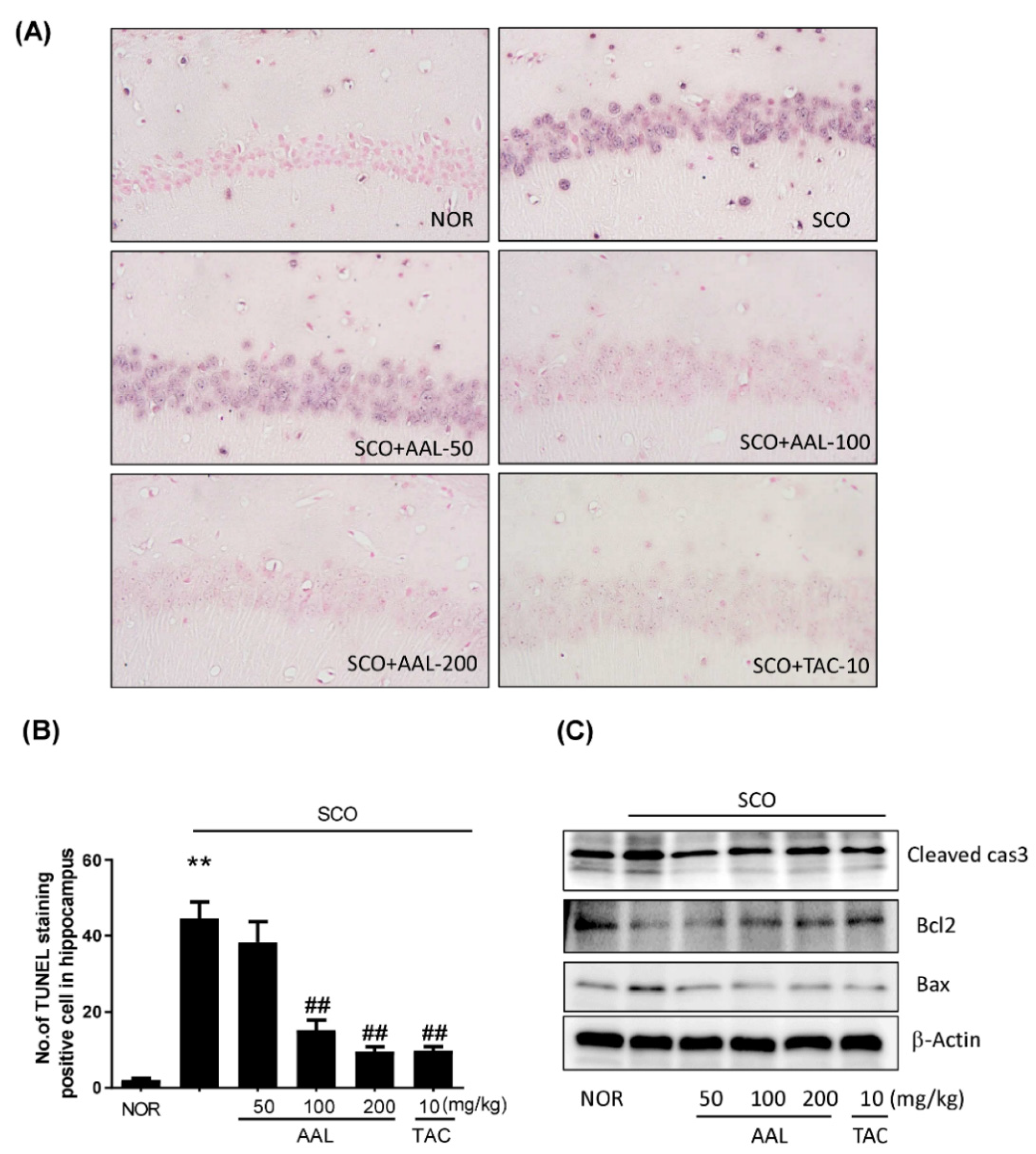
2.5. Effect of AAL Extract on Apoptosis in SCO-Treated Mice
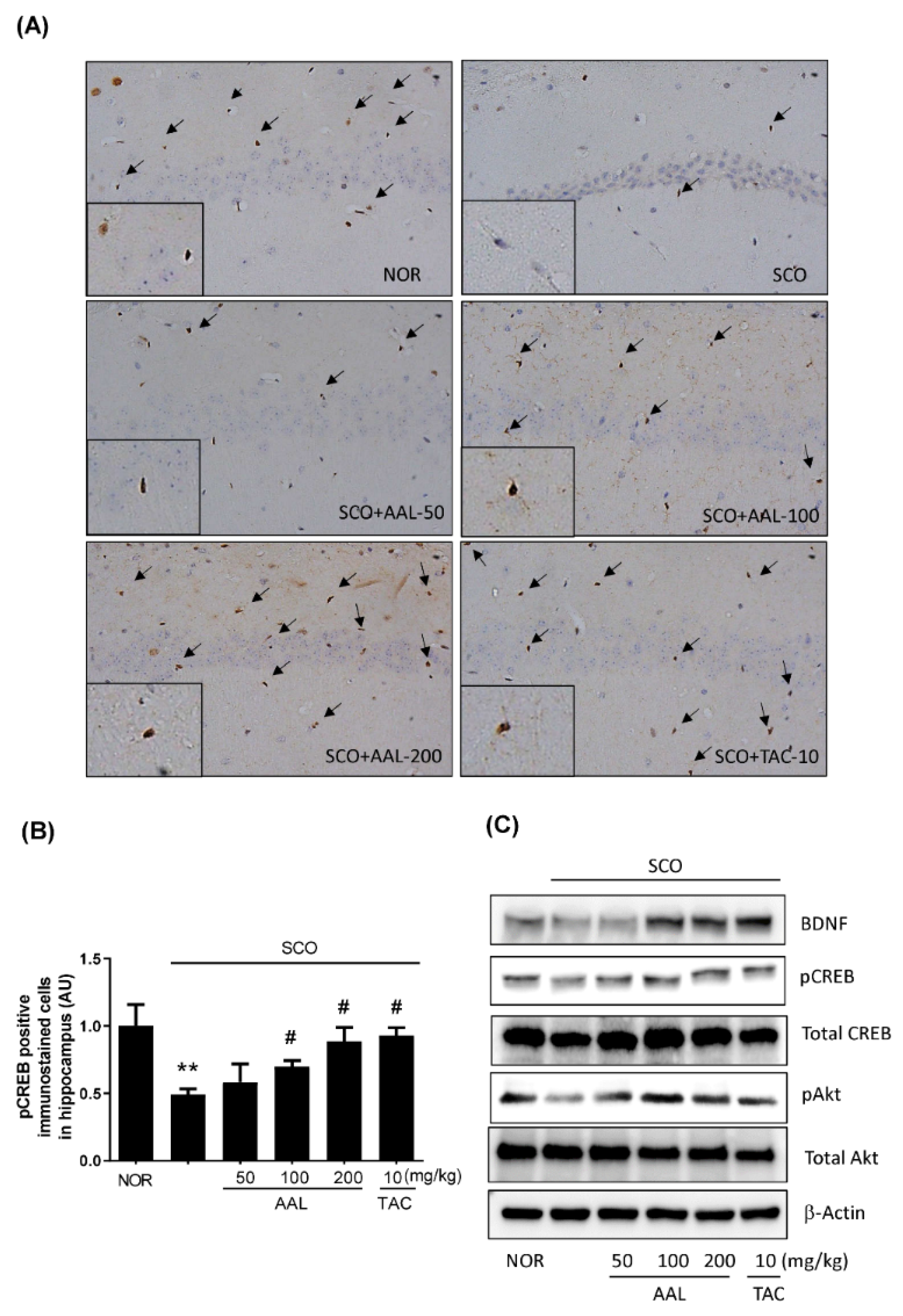
2.6. Effect of AAL Extract on SCO-Attenuated Expression of BDNF and Phosphorylation of CREB and Akt in the Hippocampus
3. Discussion
4. Materials and Methods
4.1. Preparation of Ethanol Extract from AAL
4.2. Reagents and Chemicals
4.3. Preparation of Sample and Standard Solutions for HPLC Analysis
4.4. Chromatographic Conditions
4.5. Animal and Drug Treatments
4.6. Brain Section and Tissue Preparation
4.7. Western Blot Analysis
4.8. Measurement of ACh Level and AChE Activity Assay
4.9. Nissl and TUNEL Staining
4.10. Passive Avoidance Test (PAT)
4.11. Y-Maze Test
4.12. Measurement of Reactive Oxygen Species (ROS) in Hippocampus
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACh | Acetylcholine |
| AChE | Acetylcholinesterase |
| pAkt | Phophorylated protein kinase B (Akt) |
| PBS | Phosphate-buffered saline |
| Bcl2 | B-cell lymphoma 2 |
| Bax | Bcl2-associated X protein |
| pCREB | Phosphorylated cAMP response elements binding protein |
| BDNF | Brain derived neurotrophic factor |
| ChAT | choline acetyltransferase |
| HPLC | High-performance liquid chromatography |
| TFA | Trifluoroacetic acid |
| ABTS | 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) |
| DPPH | 2,2diphenyl-1-picrylhydrazyl |
| DCFH | dichlorodihydrofluorescin |
| DCF | 2′, 7′-dichlorodihydrofluorescein |
| RIPA | Radioimmunoprecipitation assay buffer |
| ROS | reactive oxygen species |
| TUNEL | Terminal deoxynucleotidyl transferase dUTP nick end labeling |
References
- Bartus, R.T.; Dean, R.L., 3rd. Pharmaceutical treatment for cognitive deficits in Alzheimer’s disease and other neurodegenerative conditions: exploring new territory using traditional tools and established maps. Psychopharmacology 2009, 202, 15–36. [Google Scholar] [CrossRef] [PubMed]
- Lazarov, O.; Mattson, M.P.; Peterson, D.A.; Pimplikar, S.W.; van Praag, H. When neurogenesis encounters aging and disease. Trends Neurosci. 2010, 33, 569–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.S.; Kim, Y.J.; Sohn, E.; Yoon, J.; Kim, B.Y.; Jeong, S.J. Bojungikgi-Tang, a Traditional Herbal Formula, Exerts Neuroprotective Effects and Ameliorates Memory Impairments in Alzheimer’s Disease-Like Experimental Models. Nutrients 2018, 10, 1952. [Google Scholar] [CrossRef] [PubMed]
- Scheff, S.W.; Price, D.A.; Schmitt, F.A.; Mufson, E.J. Hippocampal synaptic loss in early Alzheimer’s disease and mild cognitive impairment. Neurobiol. Aging 2006, 27, 1372–1384. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Wang, Y.; Xu, T.; Li, Q.; Wang, D.; Zhang, L.; Fan, B.; Wang, F.; Liu, X. Genistein ameliorates scopolamine-Induced amnesia in mice through the regulation of the cholinergic neurotransmission, antioxidant system and the ERK/CREB/BDNF signaling. Front. Pharmacol. 2018, 9, 1153. [Google Scholar] [CrossRef] [PubMed]
- Araujo, J.A.; Studzinski, C.M.; Milgram, N.W. Further evidence for the cholinergic hypothesis of aging and dementia from the canine model of aging. Prog. Neuropsychopharmacol. Biol. Psychiatry 2005, 29, 411–422. [Google Scholar] [CrossRef]
- Niedzielska, E.; Smaga, I.; Gawlik, M.; Moniczewski, A.; Stankowicz, P.; Pera, J.; Filip, M. Oxidative stress in neurodegenerative diseases. Mol. Neurobiol. 2016, 53, 4094–4125. [Google Scholar] [CrossRef]
- Terry, A.V., Jr.; Buccafusco, J.J. The cholinergic hypothesis of age and Alzheimer’s disease-related cognitive deficits: recent challenges and their implications for novel drug development. J. Pharmacol. Exp. Ther. 2003, 306, 821–827. [Google Scholar] [CrossRef]
- Zhou, X.; Li, Y.; Shi, X.; Ma, C. An overview on therapeutics attenuating amyloid beta level in Alzheimer’s disease: Targeting neurotransmission, inflammation, oxidative stress and enhanced cholesterol levels. Am. J. Transl. Res. 2016, 8, 246–269. [Google Scholar]
- Lu, C.; Dong, L.; Lv, J.; Wang, Y.; Fan, B.; Wang, F.; Liu, X. 20(S)-protopanaxadiol (PPD) alleviates scopolamine-induced memory impairment via regulation of cholinergic and antioxidant systems, and expression of Egr-1, c-Fos and c-Jun in mice. Chem. Biol. Interact. 2018, 279, 64–72. [Google Scholar] [CrossRef]
- Jeon, S.J.; Kim, B.; Park, H.J.; Zhang, J.; Kwon, Y.; Kim, D.H.; Ryu, J.H. The ameliorating effect of 1-palmitoyl-2-linoleoyl-3-acetylglycerol on scopolamine-induced memory impairment via acetylcholinesterase inhibition and LTP activation. Behav. Brain Res. 2017, 324, 58–65. [Google Scholar] [CrossRef]
- De Cássia Seffrin, R.; Shikano, I.; Akhtar, Y.; Isman, M.B. Effects of crude seed extracts of Annona atemoya and Annona squamosa L. against the cabbage looper, Trichoplusia ni in the laboratory and greenhouse. Crop Prot. 2010, 29, 20–24. [Google Scholar] [CrossRef]
- Wongs-Aree, C.; Noichinda, S. Sugar apple (Annona squamosa L.) and atemoya (A. cherimola Mill. × A. squamosa L.). In Postharvest Biology and Technology of Tropical and Subtropical Fruits; Woodhead Publishing: Cambridge, UK, 2011; Volume 4, pp. 399–426. [Google Scholar]
- Dutra, L.M.; Costa, E.V.; Moraes, V.R.; Nogueira, P.C.; Vendramin, M.E.; Barison, A.; Prata, A.P. Chemical constituents from the leaves of Annona pickelii (Annonaceae). Biochem. Syst. Ecol. 2012, 41, 115–118. [Google Scholar] [CrossRef] [Green Version]
- Leboeuf, M.; Cavé, A.; Bhaumik, P.K.; Mukherjee, B.; Mukherjee, R. The phytochemistry of the annonaceae. Phytochemistry 1980, 21, 2783–2813. [Google Scholar] [CrossRef]
- Paull, R.E. Tropical Fruits. Crop Product. Sci. Hortic. 2011, 1, 148–150. [Google Scholar]
- Brandão, A.E. National Composition of Fruit Cultivars; Academic Press: Waltham, MA, USA, 2016. [Google Scholar]
- Chih, H.W.; Chiu, H.F.; Tang, K.S.; Chang, F.R.; Wu, Y.C. Bullatacin, a potent antitumor annonaceous acetogenin, inhibits proliferation of human hepatocarcinoma cell line 2.2.15 by apoptosis induction. Life Sci. 2001, 69, 1321–1331. [Google Scholar] [CrossRef]
- Chiu, H.F.; Chih, T.T.; Hsian, Y.M.; Tseng, C.H.; Wu, M.J.; Wu, Y.C. Bullatacin, a potent antitumor Annonaceous acetogenin, induces apoptosis through a reduction of intracellular cAMP and cGMP levels in human hepatoma 2.2.15 cells. Biochem. Pharmacol. 2003, 65, 319–327. [Google Scholar] [CrossRef]
- Yi, J.M.; Park, J.S.; Lee, J.; Hong, J.T.; Bang, O.S.; Kim, N.S. Anti-angiogenic potential of an ethanol extract of Annona atemoya seeds in vitro and in vivo. BMC Complement Altern. Med. 2014, 14, 353. [Google Scholar] [CrossRef]
- Rabêlo, S.V.; Costa, E.V.; Barison, A.; Dutra, L.M.; Nunes, X.P.; Tomaz, J.C.; Almeida, J.R. Antioxidant and antimicrobial activity of extracts from Atemoya. Rev. Bras. Frutic. 2014, 36, 265–271. [Google Scholar] [CrossRef]
- Rabêlo, S.V.; Costa, E.V.; Barison, A.; Dutra, L.M.; Nunes, X.P.; Tomaz, J.C.; Almeida, J.R. Alkaloids isolated from the leaves of atemoya (Annona cherimola Annona squamosa). Braz. J. Pharmagn. 2015, 25, 419–421. [Google Scholar] [CrossRef]
- Anand, P.; Singh, B. A review on cholinesterase inhibitors for Alzheimer’s disease. Arch. Pharm. Res. 2013, 36, 375–399. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Liu, Q.; Wang, Y.; Luo, G. Coadministration of huperzine A and ligustrazine phosphate effectively reverses scopolamine-induced amnesia in rats. Pharmacol. Biochem. Behav. 2010, 96, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.Q.; Wu, D.W.; Zhang, C.X.; Yan, R.; Yang, C.; Rong, C.P.; Zhang, L.; Chang, X.; Su, R.Y.; Zhang, S.J.; et al. BushenYizhi formula ameliorates cognition deficits and attenuates oxidative stressrelated neuronal apoptosis in scopolamineinduced senescence in mice. Int. J. Mol. Med. 2014, 34, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Scott Bitner, R. Cyclic AMP response element-binding protein (CREB) phosphorylation: a mechanistic marker in the development of memory enhancing Alzheimer’s disease therapeutics. Biochem. Pharmacol. 2012, 83, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Kouemou, N.E.; Taiwe, G.S.; Moto, F.C.O.; Pale, S.; Ngoupaye, G.T.; Njapdounke, J.S.K.; Nkantchoua, G.C.N.; Pahaye, D.B.; Bum, E.N. Nootropic and Neuroprotective Effects of Dichrocephala integrifolia on scopolamine mouse model of Alzheimer’s Disease. Front. Pharmacol. 2017, 8, 847. [Google Scholar] [CrossRef]
- Li, S.P.; Wang, Y.W.; Qi, S.L.; Zhang, Y.P.; Deng, G.; Ding, W.Z.; Ma, C.; Lin, Q.Y.; Guan, H.D.; Liu, W.; et al. Analogous beta-Carboline Alkaloids Harmaline and Harmine Ameliorate Scopolamine-Induced Cognition Dysfunction by Attenuating Acetylcholinesterase Activity, Oxidative Stress, and Inflammation in Mice. Front. Pharmacol. 2018, 9, 346. [Google Scholar] [CrossRef]
- Orta-Salazar, E.; Cuellar-Lemus, C.A.; Diaz-Cintra, S.; Feria-Velasco, A.I. Cholinergic markers in the cortex and hippocampus of some animal species and their correlation to Alzheimer’s disease. Neurologia 2014, 29, 497–503. [Google Scholar] [CrossRef]
- Rahnama, S.; Rabiei, Z.; Alibabaei, Z.; Mokhtari, S.; Rafieian-Kopaei, M.; Deris, F. Anti-amnesic activity of Citrus aurantium flowers extract against scopolamine-induced memory impairments in rats. Neurol Sci 2015, 36, 553–560. [Google Scholar] [CrossRef]
- Alvarez-Jimenez, R.; Groeneveld, G.J.; van Gerven, J.M.; Goulooze, S.C.; Baakman, A.C.; Hay, J.L.; Stevens, J. Model-based exposure-response analysis to quantify age related differences in the response to scopolamine in healthy subjects. Br. J. Clin. Pharmacol. 2016, 82, 1011–1021. [Google Scholar] [CrossRef] [Green Version]
- Balaban, H.; Naziroglu, M.; Demirci, K.; Ovey, I.S. The protective role of selenium on scopolamine-induced memory impairment, oxidative stress, and apoptosis in aged rats: The involvement of TRPM2 and TRPV1 channels. Mol. Neurobiol. 2017, 54, 2852–2868. [Google Scholar] [CrossRef]
- Cutuli, D.; Foti, F.; Mandolesi, L.; De Bartolo, P.; Gelfo, F.; Federico, F.; Petrosini, L. Cognitive performances of cholinergically depleted rats following chronic donepezil administration. J. Alzheimers Dis. 2009, 17, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Garibotto, V.; Tettamanti, M.; Marcone, A.; Florea, I.; Panzacchi, A.; Moresco, R.; Virta, J.R.; Rinne, J.; Cappa, S.F.; Perani, D. Cholinergic activity correlates with reserve proxies in Alzheimer’s disease. Neurobiol. Aging 2013, 34, 2613–2698. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, J.; Seo, S.G.; Choi, B.R.; Han, J.S.; Lee, K.W.; Kim, J. Sulforaphane alleviates scopolamine-induced memory impairment in mice. Pharmacol. Res. 2014, 85, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Song, S.H.; Choi, S.M.; Kim, J.E.; Sung, J.E.; Lee, H.A.; Choi, Y.H.; Bae, C.J.; Choi, Y.W.; Hwang, D.Y. Alpha-Isocubebenol alleviates scopolamine-induced cognitive impairment by repressing acetylcholinesterase activity. Neurosci. Lett. 2017, 638, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Hu, J.; Li, J.; Yang, Z.; Xin, X.; Wang, J.; Ding, J.; Geng, M. Effect of acidic oligosaccharide sugar chain on scopolamine-induced memory impairment in rats and its related mechanisms. Neurosci. Lett. 2005, 374, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Budzynska, B.; Boguszewska-Czubara, A.; Kruk-Slomka, M.; Skalicka-Wozniak, K.; Michalak, A.; Musik, I.; Biala, G. Effects of imperatorin on scopolamine-induced cognitive impairment and oxidative stress in mice. Psychopharmacology 2015, 232, 931–942. [Google Scholar] [CrossRef]
- Tuppo, E.E.; Forman, L.J. Free radical oxidative damage and Alzheimer’s disease. J. Am. Osteopath. Assoc. 2001, 101, S11–S15. [Google Scholar]
- Wang, G.; Fang, H.; Zhen, Y.; Xu, G.; Tian, J.; Zhang, Y.; Zhang, D.; Zhang, G.; Xu, J.; Zhang, Z.; et al. Sulforaphane prevents neuronal apoptosis and memory impairment in diabetic rats. Cell. Physiol. Biochem. 2016, 39, 901–907. [Google Scholar] [CrossRef]
- Rossi, C.; Angelucci, A.; Costantin, L.; Braschi, C.; Mazzantini, M.; Babbini, F.; Fabbri, M.E.; Tessarollo, L.; Maffei, L.; Berardi, N.; et al. Brain-derived neurotrophic factor (BDNF) is required for the enhancement of hippocampal neurogenesis following environmental enrichment. Eur. J. Neurosci. 2006, 24, 1850–1856. [Google Scholar] [CrossRef]
- Bartolotti, N.; Bennett, D.A.; Lazarov, O. Reduced pCREB in Alzheimer’s disease prefrontal cortex is reflected in peripheral blood mononuclear cells. Mol. Psychiatr. 2016, 21, 1158–1166. [Google Scholar] [CrossRef]
- Ambrogini, P.; Cuppini, R.; Cuppini, C.; Ciaroni, S.; Cecchini, T.; Ferri, P.; Sartini, S.; Del Grande, P. Spatial learning affects immature granule cell survival in adult rat dentate gyrus. Neurosci. Lett. 2000, 286, 21–24. [Google Scholar] [CrossRef]
- Jain, V.; Baitharu, I.; Prasad, D.; Ilavazhagan, G. Enriched environment prevents hypobaric hypoxia induced memory impairment and neurodegeneration: role of BDNF/PI3K/GSK3beta pathway coupled with CREB activation. PLoS ONE 2013, 8, e62235. [Google Scholar] [CrossRef]
- Wu, H.; Lu, D.; Jiang, H.; Xiong, Y.; Qu, C.; Li, B.; Mahmood, A.; Zhou, D.; Chopp, M. Simvastatin-mediated upregulation of VEGF and BDNF, activation of the PI3K/Akt pathway, and increase of neurogenesis are associated with therapeutic improvement after traumatic brain injury. J. Neurotrauma 2008, 25, 130–139. [Google Scholar] [CrossRef]
- Ramalingayya, G.V.; Nampoothiri, M.; Nayak, P.G.; Kishore, A.; Shenoy, R.R.; Mallikarjuna Rao, C.; Nandakumar, K. Naringin and rutin alleviates episodic memory deficits in two differentially challenged object recognition tasks. Pharmacogn. Mag. 2016, 12, S63–S70. [Google Scholar]
- Khan, M.M.; Ahmad, A.; Ishrat, T.; Khuwaja, G.; Srivastawa, P.; Khan, M.B.; Raza, S.S.; Javed, H.; Vaibhav, K.; Khan, A.; et al. Rutin protects the neural damage induced by transient focal ischemia in rats. Brain Res. 2009, 1292, 123–135. [Google Scholar] [CrossRef]
- Javed, H.; Khan, M.M.; Ahmad, A.; Vaibhav, K.; Ahmad, M.E.; Khan, A.; Ashafaq, M.; Islam, F.; Siddiqui, M.S.; Safhi MM et, a.l. Rutin prevents cognitive impairments by ameliorating oxidative stress and neuroinflammation in rat model of sporadic dementia of Alzheimer type. Neuroscience 2012, 210, 340–352. [Google Scholar] [CrossRef]
- Rogerio, A.P.; Kanashiro, A.; Fontanari, C.; da Silva, E.V.; Lucisano-Valim, Y.M.; Soares, E.G.; Faccioli, L.H. Anti-inflammatory activity of quercetin and isoquercitrin in experimental murine allergic asthma. Inflamm. Res. 2007, 56, 402–408. [Google Scholar] [CrossRef]
- Jung, S.H.; Kim, B.J.; Lee, E.H.; Osborne, N.N. Isoquercitrin is the most effective antioxidant in the plant Thuja orientalis and able to counteract oxidative-induced damage to a transformed cell line (RGC-5 cells). Neurochem. Int. 2010, 57, 713–721. [Google Scholar] [CrossRef]
- Hassan, W.; Rongyin, G.; Daoud, A.; Ding, L.; Wang, L.; Liu, J.; Shang, J. Reduced oxidative stress contributes to the lipid lowering effects of isoquercitrin in free fatty acids induced hepatocytes. Oxid. Med. Cell. Longev. 2014, 2014, 313602. [Google Scholar] [CrossRef]
- Valentova, K.; Vrba, J.; Bancirova, M.; Ulrichova, J.; Kren, V. Isoquercitrin: pharmacology, toxicology, and metabolism. Food Chem. Toxicol. 2014, 68, 267–282. [Google Scholar] [CrossRef]
- Ip, P.S.; Tsim, K.W.; Chan, K.; Bauer, R. Application of complementary and alternative medicine on neurodegenerative disorders: current status and future prospects. Evid. Based Complement. Alternat. Med. 2012, 2012, 930908. [Google Scholar] [CrossRef]
- Dai, Q.; Borenstein, A.R.; Wu, Y.; Jackson, J.C.; Larson, E.B. Fruit and vegetable juices and Alzheimer’s disease: The Kame Project. Am. J. Med. 2006, 119, 751–759. [Google Scholar] [CrossRef]
- Dayanc, M.; Kibar, Y.; Irkilata, H.C.; Gok, F.; Tahmaz, L.; Ors, O.; Akyol, I. A new modification of dismembered pyeloplasty for primary ureteropelvic junction obstruction. Eur. Surg. Res. 2008, 40, 225–229. [Google Scholar] [CrossRef]
- Zhang, S.J.; Luo, D.; Li, L.; Tan, R.R.; Xu, Q.Q.; Qin, J.; Zhu, L.; Luo, N.C.; Xu, T.T.; Zhang, R.; et al. Ethyl acetate extract components of bushen-yizhi formula provides neuroprotection against scopolamine-induced cognitive impairment. Sci. Rep. 2017, 7, 9824. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sohn, E.; Lim, H.-S.; Kim, Y.J.; Kim, B.-Y.; Jeong, S.-J. Annona atemoya Leaf Extract Improves Scopolamine-Induced Memory Impairment by Preventing Hippocampal Cholinergic Dysfunction and Neuronal Cell Death. Int. J. Mol. Sci. 2019, 20, 3538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143538
Sohn E, Lim H-S, Kim YJ, Kim B-Y, Jeong S-J. Annona atemoya Leaf Extract Improves Scopolamine-Induced Memory Impairment by Preventing Hippocampal Cholinergic Dysfunction and Neuronal Cell Death. International Journal of Molecular Sciences. 2019; 20(14):3538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143538
Chicago/Turabian StyleSohn, Eunjin, Hye-Sun Lim, Yu Jin Kim, Bu-Yeo Kim, and Soo-Jin Jeong. 2019. "Annona atemoya Leaf Extract Improves Scopolamine-Induced Memory Impairment by Preventing Hippocampal Cholinergic Dysfunction and Neuronal Cell Death" International Journal of Molecular Sciences 20, no. 14: 3538. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143538