Nitrogen Fertilizer Induced Alterations in The Root Proteome of Two Rice Cultivars

Abstract
:1. Introduction
2. Results
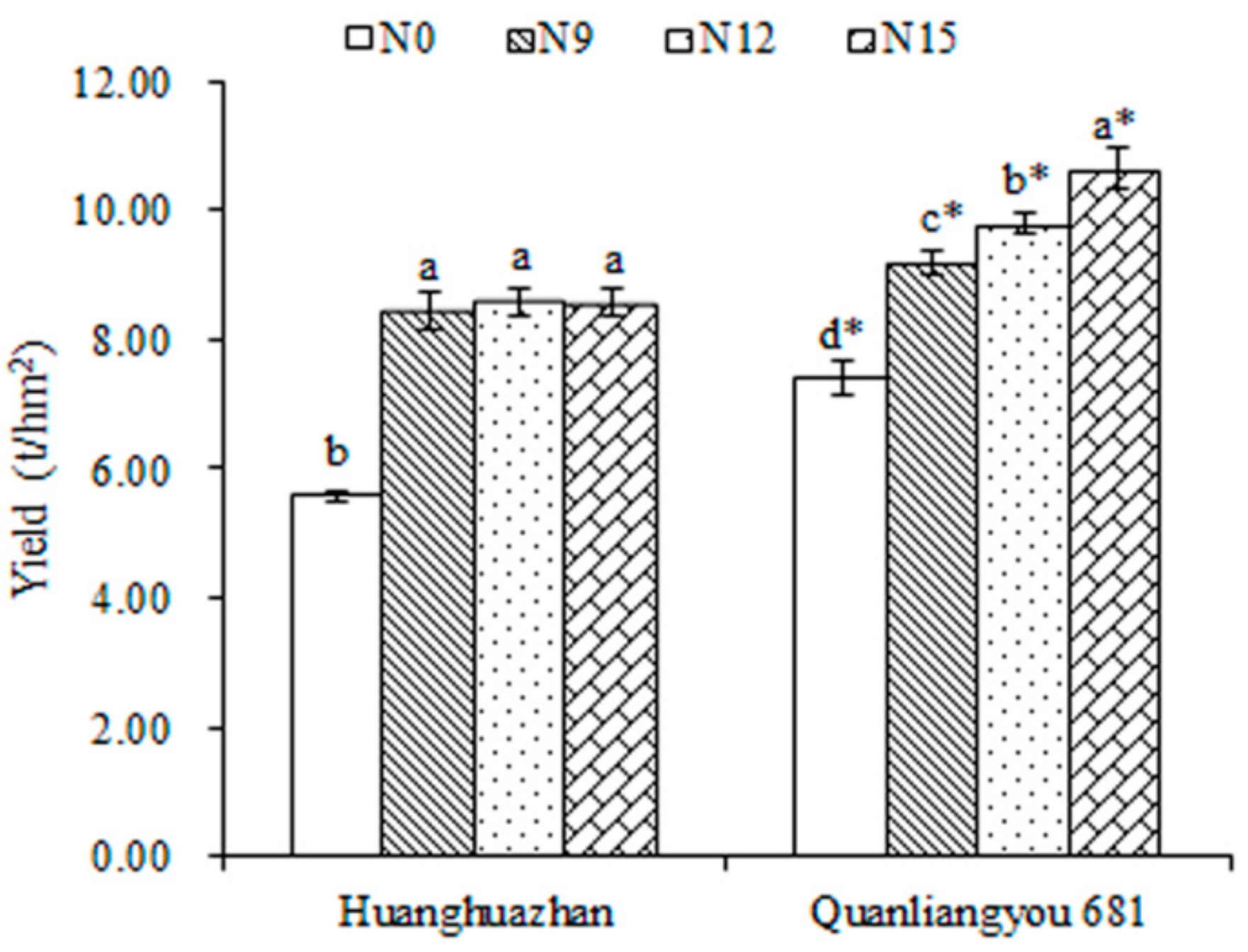
2.1. Determination of Agronomic Traits
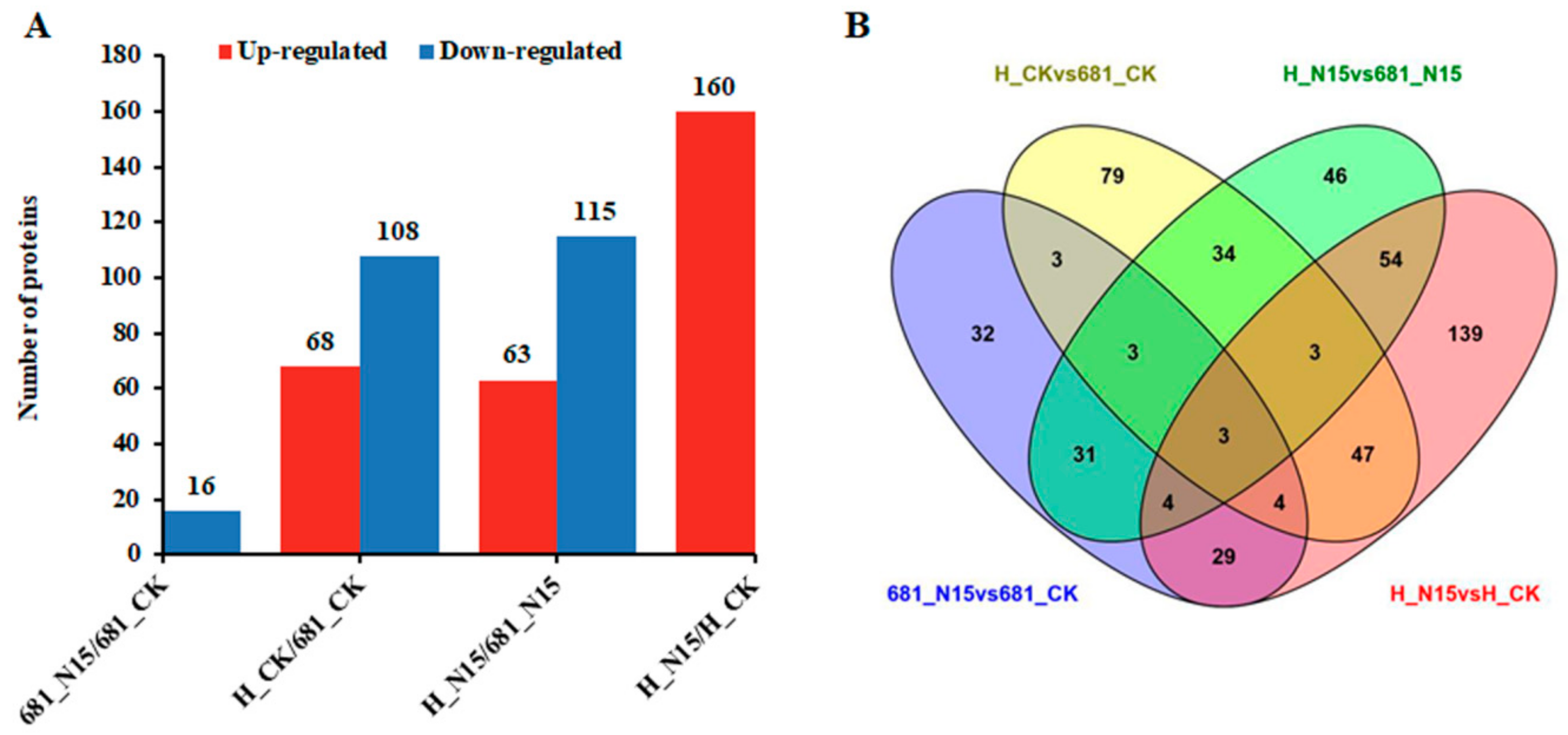
2.2. Proteome-Wide Analysis of Differentially Expressed Protein of The Two Rice Cultivars Treated With Nitrogen Fertilizer
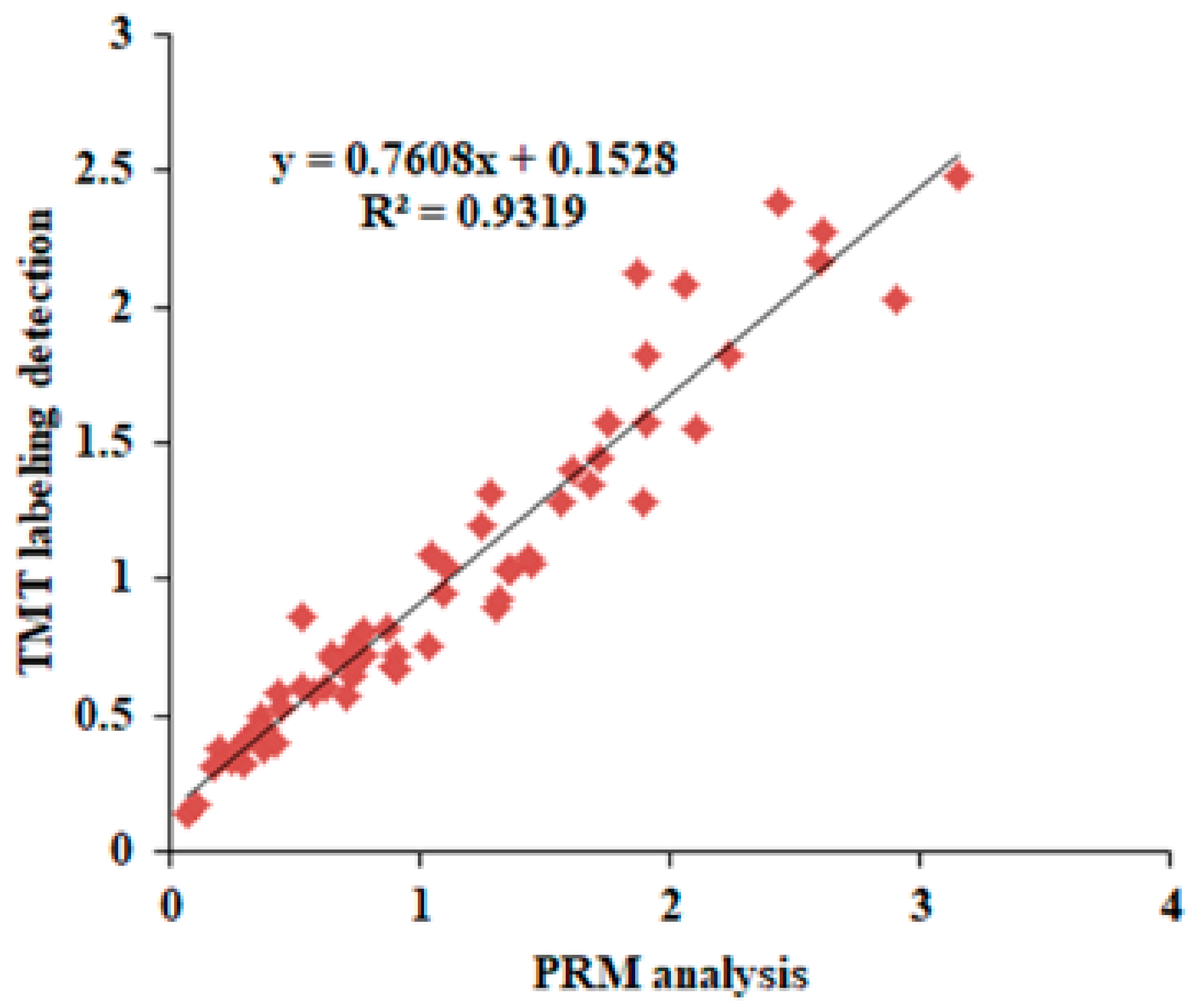
2.3. Validation of Differentially Expressed Proteins by Parallel Reaction Monitoring
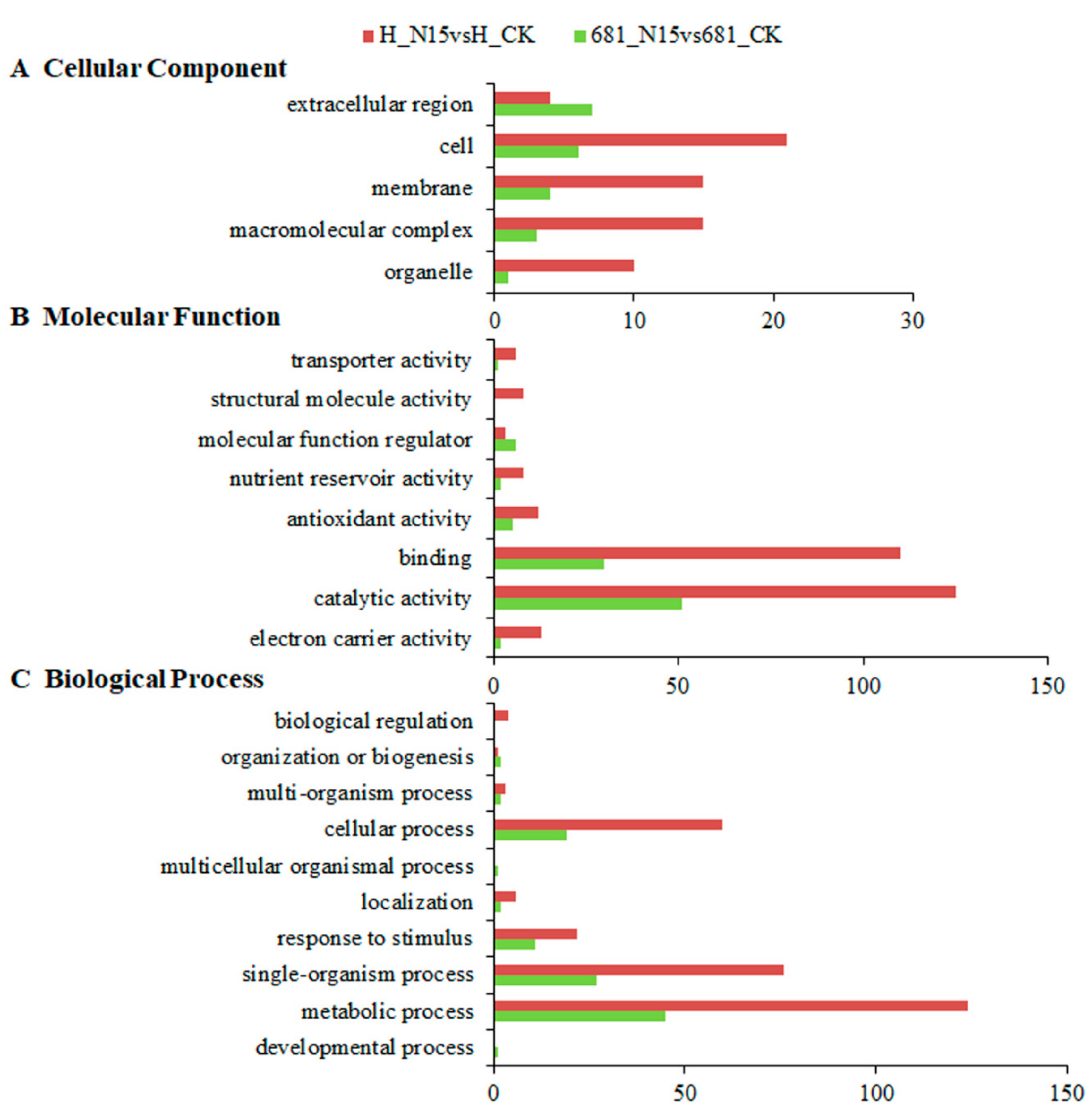
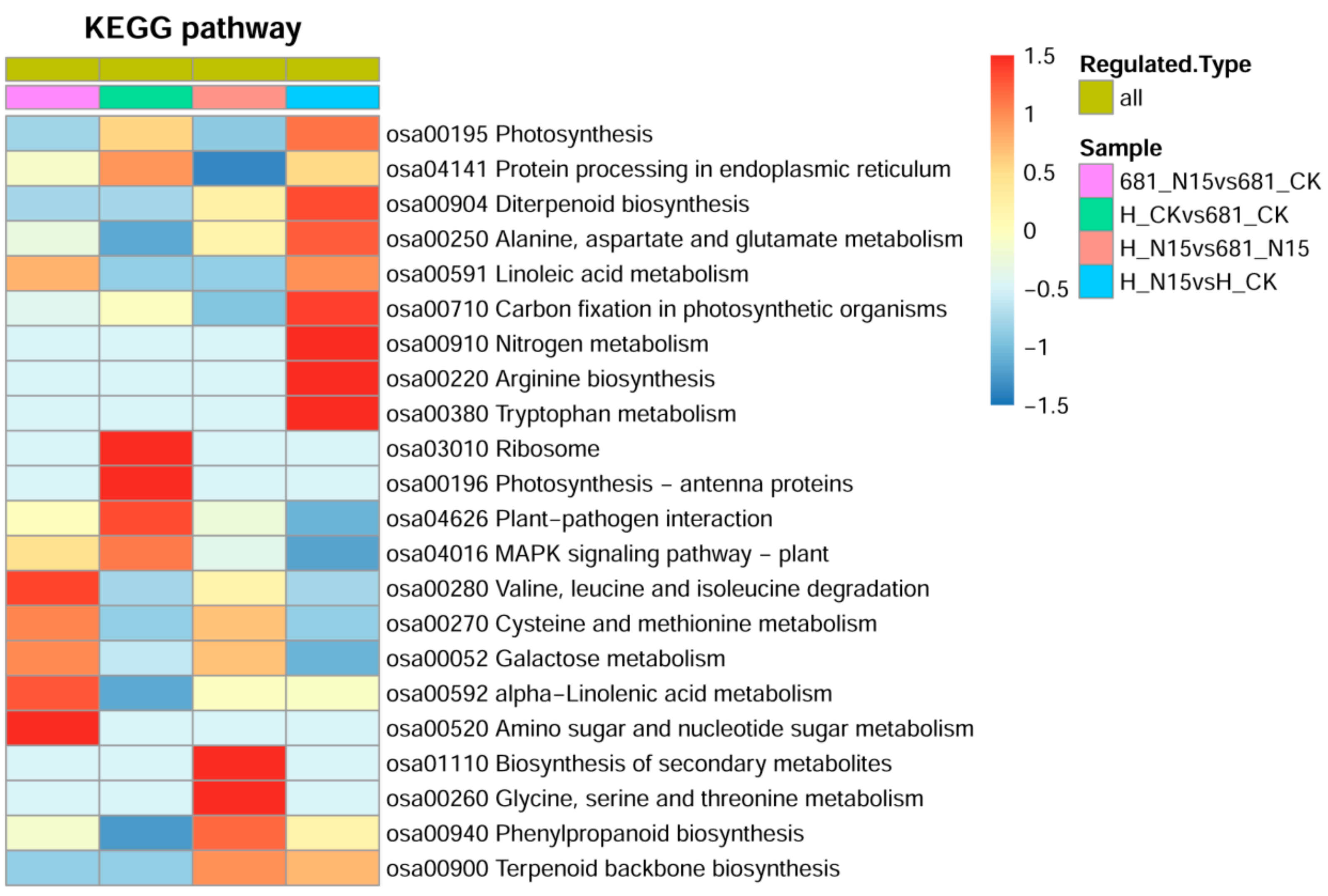
2.4. Gene Ontology Classification Analysis of Differentially Expressed Proteins
2.5. Clustering Analysis of The Differentially Expressed Proteins
3. Discussion
3.1. Ammonium Assimilation
3.2. Signal Transduction
3.3. Energy Metabolism and Material Transport
3.4. Substance Metabolism
3.5. Stress and Defense Response
4. Conclusions
5. Materials and Methods
5.1. Plant Materials and Growth Conditions
5.2. Measurement of Rice Dry Weight and Grain Yield
5.3. Protein Extraction
5.4. Trypsin Digestion
5.5. TMT Labeling
5.6. HPLC Fractionation
5.7. LC-MS/MS Analysis
5.8. Database Search
5.9. Bioinformatics Methods
5.10. Parallel Reaction Monitoring (PRM) Validation of Differentially Expressed Proteins
5.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hakeem, K.R.; Chandna, R.; Ahmad, A.; Qureshi, M.I.; Iqbal, M. Proteomic analysis for low and high nitrogen-responsive proteins in the leaves of rice genotypes grown at three nitrogen levels. Appl. Biochem. Biotechnol. 2012, 168, 834–850. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Peng, Y.; Ma, W.; Liu, R.; Li, C.; Li, X. Proteomic analysis revealed nitrogen-mediated metabolic, developmental, and hormonal regulation of maize (Zea mays L.) ear growth. J. Exp. Bot. 2012, 63, 5275–5288. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Zeng, F.; Feibo, W.; Ma, W.; Zhang, G. Proteomic analysis of nitrogen stress-responsive proteins in two rice cultivars differing in N utilization efficiency. J. Integr. Omics 2010, 1, 78–87. [Google Scholar]
- Frink, C.R.; Waggoner, P.E.; Ausubel, J.H. Nitrogen fertilizer: Retrospect and prospect. Proc. Natl. Acad. Sci. USA 1999, 96, 1175–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An overview of global rice production, supply, trade, and consumption. Ann. N Y Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H. Nitrate-specific and cytokinin-mediated nitrogen signaling pathways in plants. J. Plant Res. 2003, 116, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H.; Takei, K.; Hirose, N. Interactions between nitrogen and cytokinin in the regulation of metabolism and development. Trends Plant Sci. 2006, 11, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.M.; Arst, H.N., Jr. The molecular genetics of nitrate assimilation in fungi and plants. Annu. Rev. Genet. 1993, 27, 115–146. [Google Scholar] [CrossRef]
- Hoff, T.; Truong, H.N.; Caboche, M. The use of mutants and transgenic plants to study nitrate assimilation. Plant Cell Environ. 1994, 17, 489–506. [Google Scholar] [CrossRef]
- Lam, H.M.; Coschigano, K.T.; Oliveira, I.C.; Melo-Oliveira, R.; Coruzzi, G.M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu. Rev. Plant Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef]
- Miflin, B.J.; Lea, P.J. Ammonia assimilation. In Amino Acids and Derivatives; Academic Press: Cambridge, MA, USA, 1980; pp. 169–202. [Google Scholar]
- Pham, T.D.; Do, T.T.; Doan, T.H.Y.; Nguyen, T.A.H.; Mai, T.D.; Kobayashi, M.; Adachi, Y. Adsorptive removal of ammonium ion from aqueous solution using surfactant-modified alumina. Environ. Chem. 2017, 14, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Shrawat, A.K.; Good, A.G. Genetic Engineering Approaches to Improving Nitrogen use Efficiency. ISB News Report. May 2008. Available online: http://www.isb.vt.edu/news/2008/artspdf/may0801.pdf (accessed on 28 November 2011).
- Djennane, S.; Chauvin, J.E.; Meyer, C. Glasshouse behaviour of eight transgenic potato clones with a modified nitrate reductase expression under two fertilization regimes. J. Exp. Bot. 2002, 53, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Wang, S.; Zhang, J.; Feng, Q.; Zhang, L.; Fan, D.; Li, X.; Yuan, D.; Han, B.; Zhang, Q. Expression profiles of 10,422 genes at early stage of low nitrogen stress in rice assayed using a cDNA microarray. Plant Mol. Biol. 2006, 60, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Y.; Fang, G.; Zhao, Q.; Zeng, Q.; Li, X.; Gong, H.; Li, Y. Nitrite promotes the growth and decreases the lignin content of indica rice calli: A comprehensive transcriptome analysis of nitrite-responsive genes during in vitro culture of rice. PLoS ONE 2014, 9, e95105. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; You, J.; Liu, Z.; Rehmani, M.I.; Wang, S.; Li, G.; Wang, Q.; Ding, Y. Proteomic analysis of low nitrogen stress-responsive proteins in roots of rice. Plant Mol. Biol. Rep. 2011, 29, 618–625. [Google Scholar] [CrossRef]
- Chandna, R.; Ahmad, A. Nitrogen stress-induced alterations in the leaf proteome of two wheat varieties grown at different nitrogen levels. Physiol. Mol. Biol. Plants 2015, 21, 19–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Yang, L.; Yang, Y.; Ouyang, Z. Rice root growth and nutrient uptake as influenced by organic manure in continuously and alternately flooded paddy soils. Agric. Water Manag. 2004, 70, 67–81. [Google Scholar] [CrossRef]
- Yoshida, T.; Ancajas, R.R. Nitrogen Fixation by Bacteria in the Root Zone of Rice. Soil Sci. Soc. Am. J. 1971, 35, 156–158. [Google Scholar] [CrossRef]
- Forde, B.; Lorenzo, H. The nutritional control of root development. Plant Soil 2001, 232, 51–68. [Google Scholar] [CrossRef]
- Liu, K.; He, A.; Ye, C.; Liu, S.; Lu, J.; Gao, M.; Fan, Y.; Lu, B.; Tian, X.; Zhang, Y. Root morphological traits and spatial distribution under different nitrogen treatments and their relationship with grain yield in super hybrid rice. Sci. Rep. (UK) 2018, 8, 131. [Google Scholar] [CrossRef]
- Kim, D.Y.; Shibato, J.; Kim, D.W.; Oh, M.K.; Kim, M.K.; Shim, I.S.; Iwahashi, H.; Masuo, Y.; Rakwal, R. Gel-based proteomics approach for detecting low nitrogen-responsive proteins in cultivated rice species. Physiol. Mol. Biol. Plants 2009, 15, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.G.; Wang, Y.; Wu, J.; Kang, K.Y.; Kim, S.T. Physiological and proteomic analysis of young rice leaves grown under nitrogen-starvation conditions. Plant Biotechnol. Rep. 2011, 5, 309. [Google Scholar] [CrossRef]
- Ding, C.; You, J.; Wang, S.; Liu, Z.; Li, G.; Wang, Q.; Ding, Y. A proteomic approach to analyze nitrogen-and cytokinin-responsive proteins in rice roots. Mol. Biol. Rep. 2012, 39, 1617–1626. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Zhou, Y.; Xiao, J.; Li, X.; Zhang, Q.; Lian, X. Overexpressed glutamine synthetase gene modifies nitrogen metabolism and abiotic stress responses in rice. Plant Cell Rep. 2009, 28, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Gaufichon, L.; Reisdorf-Cren, M.; Rothstein, S.J.; Chardon, F.; Suzuki, A. Biological functions of asparagine synthetase in plants. Plant Sci. 2010, 179, 141–153. [Google Scholar] [CrossRef]
- Hirel, B.; Lea, P.J. Ammonia assimilation. In Plant Nitrogen; Springer: Berlin/Heidelberg, Germany, 2001; pp. 79–99. [Google Scholar]
- Coque, M.; Gallais, A. Genomic regions involved in response to grain yield selection at high and low nitrogen fertilization in maize. Theor. Appl. Genet. 2006, 112, 1205–1220. [Google Scholar] [CrossRef] [PubMed]
- Gallais, A.; Hirel, B. An approach to the genetics of nitrogen use efficiency in maize. J. Exp. Bot. 2004, 55, 295–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiu, S.H.; Bleecker, A.B. Receptor-like kinases from Arabidopsis form a monophyletic gene family related to animal receptor kinases. Proc. Natl. Acad. Sci. USA 2001, 98, 10763–10768. [Google Scholar] [CrossRef] [PubMed]
- Bleecker, A.B.; Kende, H. Ethylene: A gaseous signal molecule in plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 1–18. [Google Scholar] [CrossRef]
- Inoue, T.; Higuchi, M.; Hashimoto, Y.; Seki, M.; Kobayashi, M.; Kato, T.; Tabata, S.; Shinozaki, K.; Kakimoto, T. Identification of CRE1 as a cytokinin receptor from Arabidopsis. Nature 2001, 409, 1060. [Google Scholar] [CrossRef]
- Walker, J.C. Structure and function of the receptor-like protein kinases of higher plants. Plant Mol. Biol. 1994, 26, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Torii, K.U. Receptor kinase activation and signal transduction in plants: An emerging picture. Curr. Opin. Plant Biol. 2000, 3, 361–367. [Google Scholar] [CrossRef]
- Yang, S.F.; Hoffman, N.E. Ethylene biosynthesis and its regulation in higher plants. Annu. Rev. Plant Physiol. 1984, 35, 155–189. [Google Scholar] [CrossRef]
- Sehnke, P.C.; Chung, H.J.; Wu, K.; Ferl, R.J. Regulation of starch accumulation by granule-associated plant 14-3-3 proteins. Proc. Natl. Acad. Sci. USA 2001, 98, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Finnie, C.; Borch, J.; Collinge, D.B. 14-3-3 proteins: Eukaryotic regulatory proteins with many functions. Plant Mol. Biol. 1999, 40, 545–554. [Google Scholar] [CrossRef]
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. (Eds.) Biochemistry and Molecular Biology of Plants, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Mitchell, P. Chemiosmotic coupling in oxidative and photosynthetic phosphorylation. Biol. Rev. 1996, 41, 445–501. [Google Scholar] [CrossRef]
- Sarafian, V.; Kim, Y.; Poole, R.J.; Rea, P.A. Molecular cloning and sequence of cDNA encoding the pyrophosphate-energized vacuolar membrane proton pump of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1992, 89, 1775–1779. [Google Scholar] [CrossRef]
- Higgins, C.F. ABC transporters: From microorganisms to man. Annu. Rev. Cell Biol. 1992, 8, 67–113. [Google Scholar] [CrossRef]
- Dean, M.; Allikmets, R. Evolution of ATP-binding cassette transporter genes. Curr. Opin. Genet. Dev. 1995, 5, 779–785. [Google Scholar] [CrossRef]
- Dean, M.; Hamon, Y.; Chimini, G. The human ATP-binding cassette (ABC) transporter superfamily. J. Lipid Res. 2001, 42, 1007–1017. [Google Scholar] [CrossRef]
- Christen, P.; Metzler, D.E. Transaminases; John Wiley & Sons: Hoboken, NJ, USA, 1985. [Google Scholar]
- Calton, G.J.; Wood, L.L.; Updike, M.H.; Lantz, L.; Hamman, J.P. The Production of L–Phenylalanine by Polyazetidine Immobilized Microbes. Bio/technology 1986, 4, 317. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Reisdorf-Cren, M.; Pageau, K.; Lelandais, M.; Grandjean, O.; Kronenberger, J.; Valadier, M.H.; Feraud, M.; Jouglet, T.; Suzuki, A. Glutamine synthetase-glutamate synthase pathway and glutamate dehydrogenase play distinct roles in the sink-source nitrogen cycle in tobacco. Plant Physiol. 2006, 140, 444–456. [Google Scholar] [CrossRef] [PubMed]
- Purnell, M.P.; Botella, J.R. Tobacco isoenzyme 1 of NAD (H)-dependent glutamate dehydrogenase catabolizes glutamate in vivo. Plant Physiol. 2007, 143, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, Y.; Good, A.G. NAD (H)-dependent glutamate dehydrogenase is essential for the survival of Arabidopsis thaliana during dark-induced carbon starvation. J. Exp. Bot. 2008, 59, 667–680. [Google Scholar] [CrossRef] [PubMed]
- Larsen, T.M.; Boehlein, S.K.; Schuster, S.M.; Richards, N.G.; Thoden, J.B.; Holden, H.M.; Rayment, I. Three-dimensional structure of Escherichia coli asparagine synthetase B: A short journey from substrate to product. Biochemistry 1999, 38, 16146–16157. [Google Scholar] [CrossRef] [PubMed]
- Scofield, M.A.; Lewis, W.S.; Schuster, S.M. Nucleotide sequence of Escherichia coli asnB and deduced amino acid sequence of asparagine synthetase B. J. Biol. Chem. 1990, 265, 12895–12902. [Google Scholar] [PubMed]
- Lea, P.J.; Ireland, R.J. The Enzymes of Glutamine. Glutamate, Asparagine, and Aspartate Metabolism. Plant Amino Acids; CRC Press: Boca Raton, FL, USA, 1998; pp. 63–124. [Google Scholar]
- Babcock, G.D.; Esen, A. Substrate specificity of maize β-glucosidase. Plant Sci. 1994, 101, 31–39. [Google Scholar] [CrossRef]
- Campbell, J.A.; Davies, G.J.; Bulone, V.; Henrissat, B. A classification of nucleotide-diphospho-sugar glycosyltransferases based on amino acid sequence similarities. Biochem. J. 1997, 326, 929. [Google Scholar] [CrossRef] [PubMed]
- Radominsk-Pandya, A.; Czernik, P.J.; Little, J.M.; Battaglia, E.; Mackenzie, P.I. Structural and functional studies of UDP-glucuronosyltransferases. Drug Metab. Rev. 1999, 31, 817–899. [Google Scholar] [CrossRef]
- Ünligil, U.M.; Rini, J.M. Glycosyltransferase structure and mechanism. Curr. Opin. Struct. Biol. 2000, 10, 510–517. [Google Scholar]
- Murray, B.W.; Wittmann, V.; Burkart, M.D.; Hung, S.C.; Wong, C.H. Mechanism of human α-1, 3-fucosyltransferase V: Glycosidic cleavage occurs prior to nucleophilic attack. Biochemistry 1997, 36, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Singh, A.N.; Raushel, F.M. Analysis of the galactosyltransferase reaction by positional isotope exchange and secondary deuterium isotope effects. Arch. Biochem. Biophys. 1998, 267, 54–59. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, H.; Qin, R.; Jiang, W.; Liu, D. Cadmium induction of lipid peroxidation and effects on root tip cells and antioxidant enzyme activities in Vicia faba L. Ecotoxicology 2009, 18, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Dubey, R.S. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci. 2003, 164, 645–655. [Google Scholar] [CrossRef]
- Alvarez, M.E.; Lamb, C. Oxidative burst-mediated defense responses in plant disease resistance. Cold Spring Harbor Monogr. Arch. 1997, 34, 815–839. [Google Scholar]
- Tunnacliffe, A.; Wise, M.J. The continuing conundrum of the LEA proteins. Naturwissenschaften 2007, 94, 791–812. [Google Scholar] [CrossRef]
- Close, T.J. Dehydrins: Emergence of a biochemical role of a family of plant dehydration proteins. Physiol. Plantarum 1996, 97, 795–803. [Google Scholar] [CrossRef]
- Nylander, M.; Svensson, J.; Palva, E.T.; Welin, B.V. Stress-induced accumulation and tissue-specific localization of dehydrins in Arabidopsis thaliana. Plant Mol. Biol. 2001, 45, 263–279. [Google Scholar] [CrossRef]
- Hara, M.; Terashima, S.; Fukaya, T.; Kuboi, T. Enhancement of cold tolerance and inhibition of lipid peroxidation by citrus dehydrin in transgenic tobacco. Planta 2003, 217, 290–298. [Google Scholar]
- Sivamani, E.; Bahieldin, A.; Wraith, J.M.; Al-Niemi, T.; Dyer, W.E.; Ho, T.H.D.; Qu, R. Improved biomass productivity and water use efficiency under water deficit conditions in transgenic wheat constitutively expressing the barley HVA1 gene. Plant Sci. 2000, 155, 1–9. [Google Scholar] [CrossRef]
- Xu, D.; Duan, X.; Wang, B.; Hong, B.; Ho, T.H.D.; Wu, R. Expression of a late embryogenesis abundant protein gene, HVA1, from barley confers tolerance to water deficit and salt stress in transgenic rice. Plant Physiol. 1996, 110, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Hawkins, T.; Hussey, P.J.; Edwards, R. Enzyme activities and subcellular localization of members of the Arabidopsis glutathione transferase superfamily. J. Exp. Bot. 2009, 60, 1207–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.A.O.; Li, Y.S.; Chen, X.F.; Hu, J.; Chang, X.U.N.; Zhu, Y.G. Transgenic tobacco plants overexpressing cotton glutathione S-transferase (GST) show enhanced resistance to methyl viologen. J. Plant Physiol. 2003, 160, 1305–1311. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, G.K.; Rakwal, R.; Tamogami, S.; Yonekura, M.; Kubo, A.; Saji, H. Chitosan activates defense/stress response (s) in the leaves of Oryza sativa seedlings. Plant Physiol. Biochem. 2002, 40, 1061–1069. [Google Scholar] [CrossRef]
- Jung, Y.H.; Lee, J.H.; Agrawal, G.K.; Rakwal, R.; Kim, J.A.; Shim, J.K.; Lee, S.K.; Jeon, J.S.; Koh, H.J.; Lee, Y.H.; et al. The rice (Oryza sativa) blast lesion mimic mutant, blm, may confer resistance to blast pathogens by triggering multiple defense-associated signaling pathways. Plant Physiol. Biochem. 2005, 43, 397–496. [Google Scholar] [CrossRef] [PubMed]
- Nojiri, H.; Sugimori, M.; Yamane, H.; Nishimura, Y.; Yamada, A.; Shibuya, N.; Kodama, O.; Murofushi, N.; Omori, T. Involvement of jasmonic acid in elicitor-induced phytoalexin production in suspension-cultured rice cells. Plant Physiol. 1996, 110, 387–392. [Google Scholar] [CrossRef]
- Sharma, N.; Sharma, K.P.; Gaur, R.K.; Gupta, V.K. Role of chitinase in plant defense. Asian J. Biochem. 2011, 6, 29–37. [Google Scholar] [CrossRef]
- Gupta, V.; Misra, A.; Gupta, A.; Pandey, B.; Gaur, R. Rapd-Pcr of Trichoderma Isolates and In Vitro Antagonism Against Fusarium Wilt Pathogens of Psidium Guajaval. J. Plant Prot. Res. 2010, 50, 256–262. [Google Scholar] [CrossRef]
- Punja, Z.K.; Zhang, Y.Y. Plant chitinases and their roles in resistance to fungal diseases. J. Nematol. 1993, 25, 526. [Google Scholar]
- Hu, Z.; Liu, A.; Bi, A.; Amombo, E.; Gitau, M.M.; Huang, X.; Liang, C.; Fu, J. Identification of differentially expressed proteins in bermudagrass response to cold stress in the presence of ethylene. Environ. Exp. Bot. 2017, 139, 67–78. [Google Scholar] [CrossRef]
- Zhou, H.; Finkemeier, I.; Guan, W.; Tossounian, M.A.; Wei, B.; Young, D.; Huang, J.; Messens, J.; Yang, X.; Zhu, J.; et al. Oxidative stress-triggered interactions between the succinyl-and acetyl-proteomes of rice leaves. Plant Cell Environ. 2018, 41, 1139–1153. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.M. The bicinchoninic acid (BCA) assay for protein quantitation. In The Protein Protocols Handbook; Humana Press: Totowa, NJ, USA, 2009; pp. 11–15. [Google Scholar]
- Peterson, A.C.; Russell, J.D.; Bailey, D.J.; Westphall, M.S.; Coon, J.J. Parallel reaction monitoring for high resolution and high mass accuracy quantitative, targeted proteomics. Mol. Cell Proteomics 2012, 11, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Rauniyar, N. Parallel reaction monitoring: a targeted experiment performed using high resolution and high mass accuracy mass spectrometry. Int. J. Mol. Sci. 2015, 16, 28566–28581. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Level | Tillering Stage | Jointing Stage | Full Heading Stage | Mature Stage |
|---|---|---|---|---|---|
| Huanghuazhan | N0 | 0.57 ± 0.06c | 2.34 ± 0.17c | 8.70 ± 0.37c | 11.86 ± 0.19b |
| N9 | 1.21 ± 0.12b | 3.63 ± 0.04b | 10.29 ± 0.29c | 16.68 ± 0.26a | |
| N12 | 1.57 ± 0.15a | 4.38 ± 0.17a | 12.58 ± 0.45b | 17.06 ± 0.35a | |
| N15 | 1.78 ± 0.15a | 4.61 ± 0.20a | 14.94 ± 0.54a | 16.86 ± 0.27a | |
| Quanliangyou 681 | N0 | 0.72 ± 0.07b | 3.07 ± 0.14d * | 9.94 ± 0.46c * | 14.77 ± 0.16b * |
| N9 | 2.15 ± 0.02a * | 4.25 ± 0.26c * | 14.76 ± 0.16b * | 18.21 ± 0.29a * | |
| N12 | 2.39 ± 0.19a * | 4.91 ± 0.07b * | 14.59 ± 0.062b * | 18.10 ± 0.23a * | |
| N15 | 2.48 ± 0.25a * | 5.76 ± 0.17a * | 16.26 ± 0.13a * | 19.23 ± 1.15a * |
| Protein Accession | Protein Description | Gene Name | H_N15/H_CK | 681_N15/681_CK |
|---|---|---|---|---|
| B8AQE3 | Glutamine synthetase | OsI_13264 | 1.359 | 3.216 |
| B8AVN6 | Glutamine synthetase | OsI_17756 | 0.486 | 0.782 |
| B8AJN3 | Glutamine synthetase | OsI_10575 | 0.259 | 0.605 |
| B8AZE9 | Glutathione synthetase | OsI_18978 | 2.226 | 1.734 |
| B8AWG6 | glutamate synthase 2 [NADH], chloroplastic | OsI_20922 | 0.308 | 0.605 |
| B8A9D2 | probable glutamate carboxypeptidase 2 isoform X1 | OsI_03680 | 2.028 | 1.108 |
| B8AGA6 | glutathionyl-hydroquinone reductase YqjG isoform X2 | OsI_06963 | 2.115 | 1.618 |
| Protein Accession | Protein Description | Gene Name | H_N15/H_CK | 681_N15/681_CK |
|---|---|---|---|---|
| A2Z8U1 | putative receptor-like protein kinase | OsI_34134 | 3.803 | 1.294 |
| A2Y361 | receptor-like protein kinase FERONIA | OsI_19449 | 2.090 | 1.301 |
| B8B6Z1 | cysteine-rich receptor-like protein kinase 25 | OsI_26338 | 1.814 | 2.017 |
| A2ZC94 | 1-aminocyclopropane-1-carboxylate oxidase | OsI_35404 | 2.959 | 3.840 |
| B8AY41 | 1-aminocyclopropane-1-carboxylate oxidase | OsI_18467 | 2.102 | 2.676 |
| B8AGJ3 | serine/threonine-protein kinase STY46 | OsI_05618 | 1.331 | 2.621 |
| A2XEP1 | phosphoenolpyruvate carboxykinase [ATP] | OsI_10803 | 1.220 | 2.310 |
| B8B2K2 | serine/threonine-protein kinase STY46 | OsI_24506 | 0.447 | 0.745 |
| B8ANP3 | aspartokinase 1, chloroplastic isoform X1 | OsI_14341 | 0.402 | 0.359 |
| Protein Accession | Protein Description | Gene Name | H_N15/H_CK | 681_N15/681_CK |
|---|---|---|---|---|
| Energy Metabolism | ||||
| A2Z4G9 | probable NAD(P)H-dependent oxidoreductase 1 | OsI_32550 | 2.723 | 1.387 |
| B8BFL3 | probable NAD(P)H-dependent oxidoreductase 2 | OsI_32552 | 2.242 | 2.204 |
| A2XNS0 | Ferredoxin | OsI_15810 | 2.574 | 0.622 |
| A2YI62 | Ferredoxin--NADP reductase | OsI_24896 | 2.283 | 1.372 |
| A2ZCK1 | Alcohol dehydrogenase family-3 | OSI9Ba083O10 | 4.092 | 1.227 |
| B8ADR5 | cytokinin dehydrogenase 2 | OsI_00771 | 3.210 | 1.205 |
| A2YBK1 | aldehyde dehydrogenase family 2 member B7, mitochondrial | OsI_22484 | 2.081 | 1.288 |
| A2XW22 | Glutamate dehydrogenase 2, mitochondrial | GDH2 | 1.416 | 2.212 |
| B8AF09 | Glyceraldehyde-3-phosphate dehydrogenase | OsI_07948 | 0.402 | 0.643 |
| A2Y8B2 | L-lactate dehydrogenase | OsI_21291 | 0.371 | 0.569 |
| A2XU83 | Glyceraldehyde-3-phosphate dehydrogenase | OsI_16160 | 0.361 | 1.098 |
| Material Transport | ||||
| B8BLE1 | ABC transporter G family member 48 | OsI_36727 | 2.274 | 1.518 |
| B8B6Q3 | ABC transporter G family member 43 | OsI_26239 | 2.152 | 1.900 |
| A2WPG7 | pyrophosphate-energized vacuolar membrane proton pump | OsI_01741 | 2.025 | 1.262 |
| A2XAP0 | pyrophosphate-energized vacuolar membrane proton pump | OsI_09320 | 0.390 | 0.685 |
| B8ACQ3 | potassium channel KAT2 isoform X1 | OsI_00861 | 0.486 | 0.783 |
| B8BBQ9 | dipeptide transport ATP-binding protein | OsI_28206 | 0.445 | 0.747 |
| A2YP92 | "transmembrane protein, putative | OsI_27085 | 0.432 | 1.146 |
| A2YVV8 | putative copper transporter 5.2 | OsI_29464 | 0.474 | 0.892 |
| A2YML7 | sugar carrier protein C | OsI_26468 | 1.572 | 2.925 |
| B8ACJ8 | calmodulin-like protein | OsI_01318 | 1.446 | 5.881 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, J.; Sun, Z.; Chen, Q.; Damaris, R.N.; Lu, B.; Hu, Z. Nitrogen Fertilizer Induced Alterations in The Root Proteome of Two Rice Cultivars. Int. J. Mol. Sci. 2019, 20, 3674. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20153674
Tang J, Sun Z, Chen Q, Damaris RN, Lu B, Hu Z. Nitrogen Fertilizer Induced Alterations in The Root Proteome of Two Rice Cultivars. International Journal of Molecular Sciences. 2019; 20(15):3674. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20153674
Chicago/Turabian StyleTang, Jichao, Zhigui Sun, Qinghua Chen, Rebecca Njeri Damaris, Bilin Lu, and Zhengrong Hu. 2019. "Nitrogen Fertilizer Induced Alterations in The Root Proteome of Two Rice Cultivars" International Journal of Molecular Sciences 20, no. 15: 3674. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20153674