Deciphering GRINA/Lifeguard1: Nuclear Location, Ca2+ Homeostasis and Vesicle Transport

Abstract
:1. Introduction
2. The Controversial Discovery of GRINA and Its Alternative Names
3. The Gene Coding for GRINA
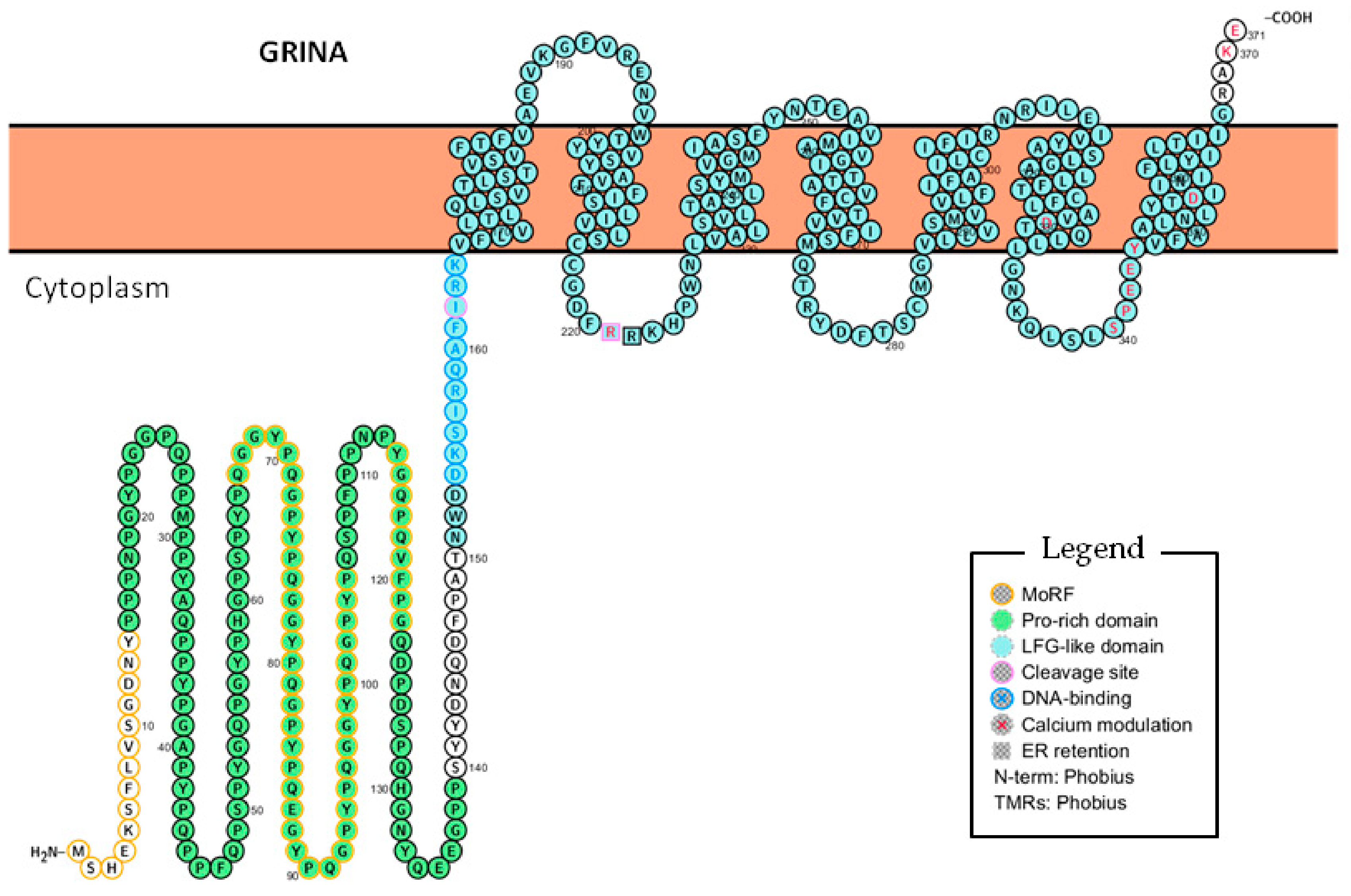
4. Deciphering the Functions of GRINA Through Its Domains and Motifs
4.1. The Lifeguard Domain of GRINA
4.2. The Pro-Rich Domain of GRINA
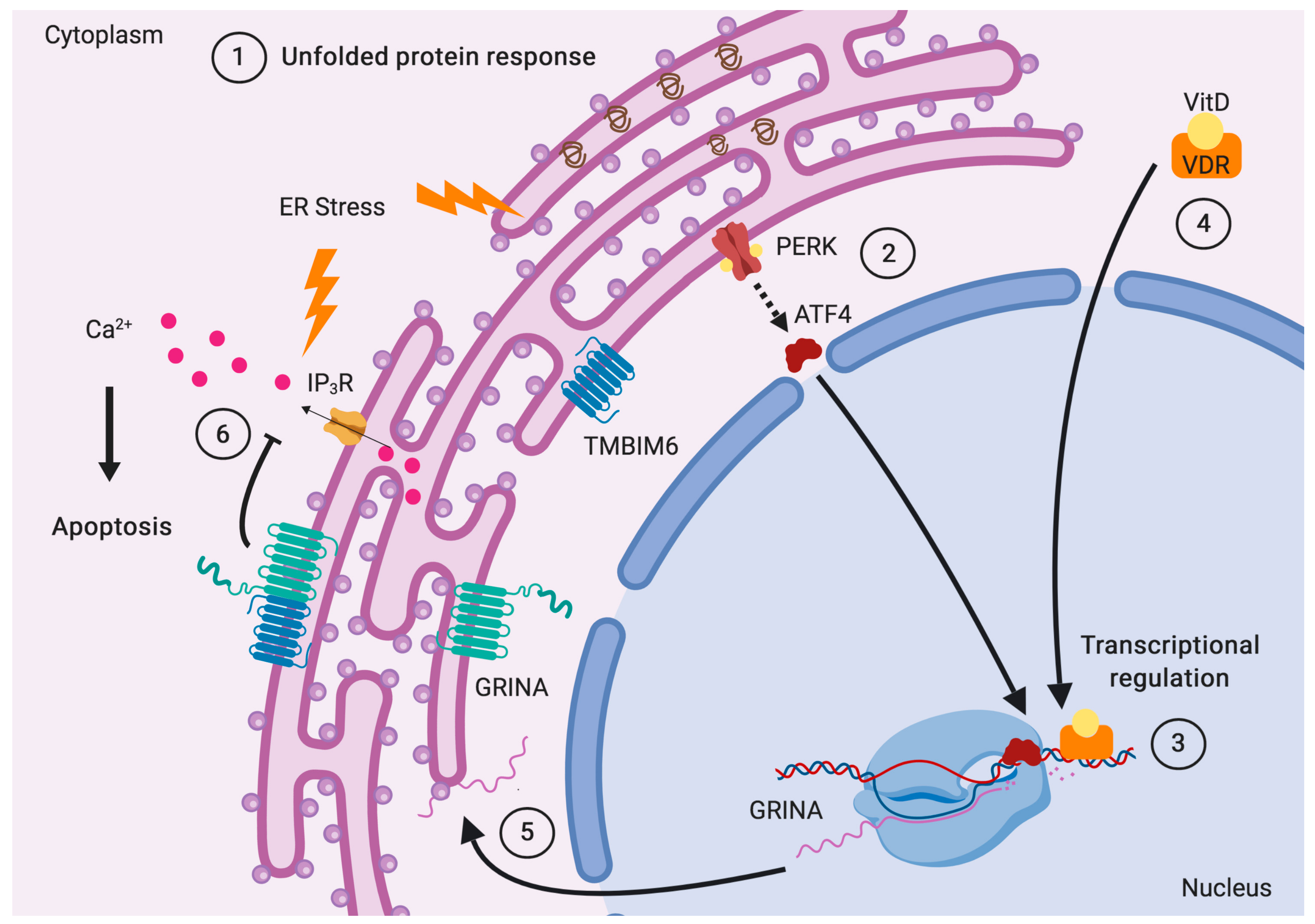
5. The Subcellular Locations of GRINA
5.1. The Unexplained Nuclear Staining of GRINA
5.2. The N-Terminal Half of GRINA Contains a Potential DNA-binding Sequence
5.3. The N-Terminal Half of GRINA May Be Cleaved or Alternatively Spliced to Enter the Nucleus
5.4. GRINA Affects Lipid and Cholesterol Metabolism
6. Transcriptional Regulation of GRINA
- Aryl-hydrocarbon receptor nuclear translocator 2 (ARNT2) is almost exclusively expressed in the brain and prevents cell death [90].
- RORA participates in the induction of ER stress response [91]. It is a key factor in calcium homeostasis pathways and plays a role in spinocerebellar ataxias [92]. It governs postnatal cerebellar development [93] and may modulate antipsychotic response in schizophrenia [94]. RORA shows a circadian expression and senses fatty acids, cholesterol, and oxysterols [95]. It is upregulated in the hippocampus of patients with Alzheimer’s disease [44] and also regulates the expression of the tight junction protein CLDND1 in human brain endothelial cells [96].
- The cAMP responsive element binding protein 3-like 1 (CREB3L1) gene codes for the membrane-bound ER stress transducer OASIS, which has an important role in osteoblast differentiation during bone development [97]. OASIS promotes the expansion of the Golgi and the synthesis of transport factors to synchronize the rise in cargo load with the amplified capacity of the secretory pathway [98]. It is preferentially expressed in astrocytes in the central nervous system (CNS) [99] and modulated by the UPR to finely control astrocyte differentiation [100]. OASIS is also involved in the terminal differentiation of mucus-secreting goblet cells in the large intestine [101].
- Doublesex and mab-3 related transcription factor 1 (DMRT1) promotes spermatogenesis and prevents female reprogramming of male gonadal cells [105].
- Inhibitor of DNA binding 4 (ID4) is up-regulated during embryogenesis and its highest expression in adult mice occurs in testes, brain and kidneys [109]. ID4 is required for the correct timing of neuronal differentiation [110] and essential for oligodendrogenesis [111]. Lack of ID4 drastically reduces osteoblast differentiation [112], but instead promotes de novo steroidogenesis [113].
- Estrogen-related receptor gamma (ESRRG) regulates testicular steroidogenesis through direct and indirect regulation of steroidogenic gene expression [114]. It positively regulates adipocyte differentiation [115] and modulates cell proliferation and estrogen signaling in breast cancer [116]. ESRRG also plays a key role in vascular calcification [117] and is involved in the determination of bone density in women [118]. It crosstalks with ATF6a to coordinate ER stress response [119] and is implicated in hearing loss and mild developmental delays [120]. ESSRG is highly expressed in the nervous system of mice embryos [121], and mice lacking neuronal ESSRG in the cerebral cortex and hippocampus exhibit defects in spatial learning and memory [122]. ESSRG is an essential coordinator of cardiac metabolism and function [123] and regulates cardiac, gastric, and renal K+ homeostasis [124].
- Myogenic factor 6 (MYF6) is involved in muscle differentiation, whereas HOXD8 is associated with the distal gut-associated mesoderm [125]. MESP1 is a key TF required for the induction of the cardiovascular gene expression program [126], and KLF13 regulates cardiac muscle development during the embryonic period [127] and mediates repression or activation under the control of SREBP-Sp1 [128].
- Zinc finger and BTB domain containing 7C (ZBTB7C) modulates DNA binding of SREBP-1c and Sp1 facilitating fatty acid synthesis [129].
- Histone H4 transcription factor (HINFP) is a co-activator in the sterol-regulated transcription of PCSK9, a target gene of SREBP2 that modulates the degradation of hepatic LDL receptors [130]. It is the only known TF required for the expression of histone H4 genes, and its loss compromises embryo implantation [131].
- ETS homologous factor (EHF) is epithelial-specific and regulates epithelial response to injury, including inflammation, efficient would repair, and barrier maintenance [132].
- Early growth response 3 (EGR3) regulates the expression of about 330 genes, 35% of which are involved in immune responses and inflammatory processes, and 15% crosstalk with the NF-κB signaling pathway [133]. It also regulates genes involved in synaptic plasticity, memory, and cognition [134] and has been associated with schizophrenia, bipolar disorder, and depression [135].
- Binding sites for NF-kB, which play a key role in inflammatory processes, are present also in the promoter region of adhesion molecules, cytokines, and growth factors [136].
- The MYC gene locates near GRINA at 8q24.21. Myc proto-oncogene (c-Myc) is a master regulator of oncogenesis, aerobic glucolysis [137], and immune response [138], coordinating protein synthesis through the transcriptional control of the ribosome’s biogenesis [139]. The expression of GRINA is upregulated in breast cancer, colorectal cancer and gastric cancer [13]. A recent article [87] confirmed that GRINA is upregulated in gastric cancer and that its regulation is mediated by c-Myc. The authors found that upon knockdown of GRINA, antiapoptotic Bcl-2 and Bcl-XL were downregulated, while proapoptotic Bax and Bak were upregulated. TMBIM6, TMBIM4, and TMBIM2 have also been shown to interact with the antiapoptotic proteins Bcl-2 and Bcl-XL [12]. Knockdown of GRINA also decreased CyclinD1 and CyclinE expression, which are involved in the G1 phase transition, and inhibited phosphorylation of Akt and the downstream effector mTOR, indicating that GRINA modulates aerobic glycolysis though that pathway.
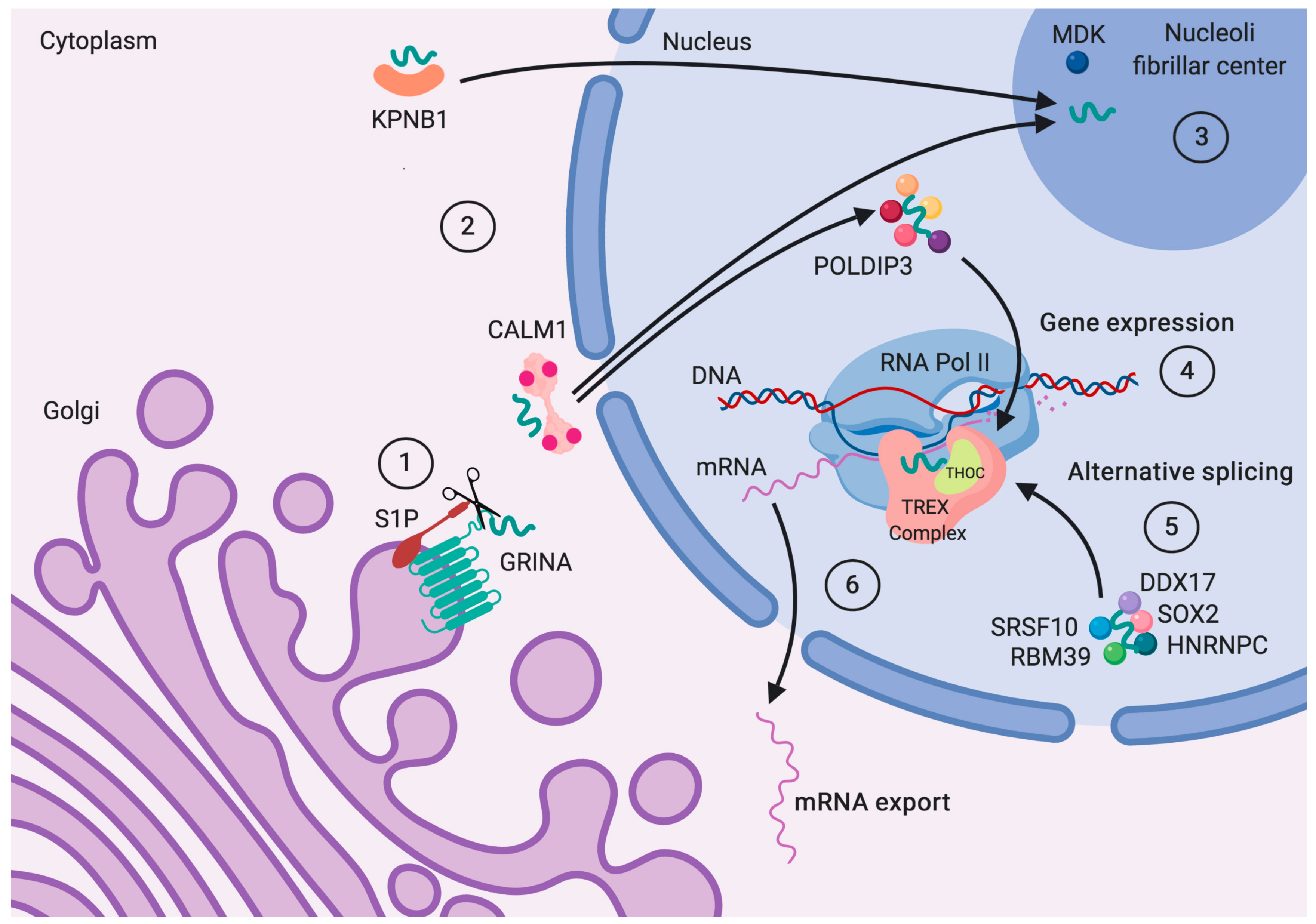
7. The Draft Protein–Protein Interaction Network of GRINA
7.1. Ticket to the Nucleus
7.2. Role in Vesicle Traffic and Cell Adhesion
7.3. Alternative Splicing to Face Changing Conditions
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| LFG | Lifeguard |
| IDR | Intrinsically Disordered Region |
| MoRF | Molecular Recognition Feature |
| ER | Endoplasmic Reticulum |
| SREBP | Sterol Regulatory Element-Binding Element |
| LDL | Low-Density Lipoprotein |
| ABM1 | ALG2-Binding Motif 1 |
| IBM | IAP-Binding Motif |
| NLS | Nuclear Location Sequence |
| TF | Transcription Factor |
References
- Moriyoshi, K.; Masu, M.; Ishii, T.; Shigemoto, R.; Mizuno, N.; Nakanishi, S. Molecular cloning and characterization of the rat NMDA receptor. Nature 1991, 354, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.N.; Tilakaratne, N.; Johnson, P.S.; Alien, A.E.; Michaelis, E.K. Cloning of cDNA for the glutamate-binding subunit of an NMDA receptor complex. Nature 1991, 354, 70–73. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.L. NMDA receptors cloned at last. Nature 1991, 354, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, E.K. Molecular biology of glutamate receptors in the central nervous system and their role in excitotoxicity, oxidative stress and aging. Prog. Neurobiol. 1998, 54, 369–415. [Google Scholar] [CrossRef]
- Hui, D.; Kumar, K.N.; Mach, J.R.; Srinivasan, A.; Pal, R.; Bao, X.; Agbas, A.; Höfner, G.; Wanner, K.T.; Michaelis, E.K. A rat brain bicistronic gene with an internal ribosome entry site codes for a phencyclidine-binding protein with cytotoxic activity. J. Biol. Chem. 2009, 284, 2245–2257. [Google Scholar] [CrossRef] [PubMed]
- Aistrup, G.L.; Szentirmay, M.; Kumar, K.N.; Babcock, K.K.; Schowen, R.L.; Michaelis, E.K. Ion channel properties of a protein complex with characteristics of a glutamate/N-methyl-D-aspartate receptor. FEBS Lett. 1996, 394, 141–148. [Google Scholar] [CrossRef]
- Sato, K.; Mick, G.; Kiyama, H.; Tohyama, M. Expression patterns of a glutamate-binding protein in the rat central nervous system: Comparison with N-methyl-D-aspartate receptor subunit 1 in rat. Neuroscience 1995, 64, 459–475. [Google Scholar] [CrossRef]
- Mattson, M.P.; Wang, H.; Michaelis, E.K. Developmental expression, compartmentalization, and possible role in excitotoxicity of a putative NMDA receptor protein in cultured hippocampal neurons. Brain Res. 1991, 565, 94–108. [Google Scholar] [CrossRef]
- Pal, R.; Eaton, M.J.; Islam, S.; Hake-Frendscho, M.; Kumar, K.N.; Michaelis, E.K. Immunocytochemical and in situ hybridization studies of the expression and distribution of three subunits of a complex with N-methyl-D-aspartate receptor-like properties. Neuroscience 1999, 94, 1291–1311. [Google Scholar] [CrossRef]
- Nielsen, J.A.; Chambers, M.A.; Romm, E.; Lee, L.Y.H.; Berndt, J.A.; Hudson, L.D. Mouse transmembrane BAX inhibitor motif 3 (Tmbim3) encodes a 38 kDa transmembrane protein expressed in the central nervous system. Mol. Cell. Biochem. 2011, 357, 73–81. [Google Scholar] [CrossRef]
- Lisak, D.A.; Schacht, T.; Enders, V.; Habicht, J.; Kiviluoto, S.; Schneider, J.; Henke, N.; Bultynck, G.; Methner, A. The transmembrane Bax inhibitor motif (TMBIM) containing protein family: Tissue expression, intracellular localization and effects on the ER CA2+-filling state. Biochim. Biophys. Acta 2015, 1853, 2104–2114. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q. TMBIM-mediated Ca2+ homeostasis and cell death. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Rivera, D.; Hetz, C. TMBIM protein family: Ancestral regulators of cell death. Oncogene 2015, 34, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Gamboa-Tuz, S.D.; Pereira-Santana, A.; Zhao, T.; Schranz, M.E.; Castano, E.; Rodriguez-Zapata, L.C. New insights into the phylogeny of the TMBIM superfamily across the three of life: Comparative genomics and synteny networks reveal independent evolution of the BI and LFG families in plants. Mol. Phylogenet. Evol. 2018, 126, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Carrara, G.; Parsons, M.; Saraiva, N.; Smith, G.L. Golgi anti-apoptotic protein: A tale of camels, calcium, channels and cancer. Open Biol. 2017, 7, 170045. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, M.; Smith, T.F.; Sudmant, P.H.; Goldberger, G. Pseudogenization of testis-specific Lfg5 predates human/Neanderthal divergence. J. Hum. Genet. 2014, 59, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Omasits, U.; Ahrens, C.H.; Müller, S.; Wollscheid, B. Protter: Interactive protein feature visualization and integration with experimental proteomic data. Bioinformatics 2014, 30, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Lewis, T.B.; Wood, S.; Michaelis, E.K.; Dupont, B.R.; Leach, R.J. Localization of a gene for a glutamate binding subunit of a NMDA receptor (GRINA) to 8q24. Genomics 1996, 32, 131–133. [Google Scholar] [CrossRef]
- Patel, A.K.; Bhatt, V.D.; Tripathi, A.K.; Sajnani, M.R.; Jakhesara, S.J.; Koringa, P.G.; Joshi, C.G. Identification of novel splice variants in horn cancer by RNA-Seq analysis in Zebu cattle. Genomics 2013, 101, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Gouw, M.; Michael, S.; Sámano-Sánchez, H.; Kumar, M.; Zeke, A.; Lang, B.; Bely, B.; Chemes, L.B.; Davey, N.E.; Deng, Z.; et al. The eukaryotic linear motif resource—2018 update. Nucleic Acids Res. 2018, 46, D428–D434. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Bruni, R.; Kloss, B.; Assur, Z.; Kloppmann, E.; Rost, B.; Hendrickson, W.A.; Liu, Q. Structural basis for a pH-sensitive calcium leak across membranes. Science 2014, 344, 1131–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelsen, K.; Yuan, H.; Schwappach, B. Hide and run: Arginine-based endoplasmic-reticulum-sorting motifs in the assembly of heteromultimeric membrane proteins. EMBO Rep. 2005, 6, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Standley, S.; Roche, K.W.; McCallum, J.; Sans, N.; Wenthold, R.J. PDZ domain suppression of an ER retention signal in NMDA receptor NR1 splice variants. Neuron 2000, 28, 887–898. [Google Scholar] [CrossRef]
- Chan, W.Y.; Soloviev, M.M.; Ciruela, F.; McIlhinney, R.A.J. Molecular determinants of metabotropic glutamate receptor 1B trafficking. Mol. Cell. Neurosci. 2001, 17, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, S.E.; Harikumar, K.B.; Hait, N.C.; Allegood, J.; Strub, G.M.; Kim, E.Y.; MacEyka, M.; Jiang, H.; Luo, C.; Kordula, T.; et al. Sphingosine-1-phosphate is a missing cofactor for the E3 ubiquitin ligase TRAF2. Nature 2010, 465, 1084–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radivojac, P.; Vacic, V.; Haynes, C.; Cocklin, R.R.; Mohan, A.; Heyen, J.W.; Goebl, M.G.; Iakoucheva, L.M. Identification, analysis, and prediction of protein ubiquitination sites. Proteins Struct. Funct. Bioinform. 2010, 78, 365–380. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Rivera, D.; Armisén, R.; Colombo, A.; Martínez, G.; Eguiguren, A.L.; Díaz, A.; Kiviluoto, S.; Rodríguez, D.; Patron, M.; Rizzuto, R.; et al. TMBIM3/GRINA is a novel unfolded protein response (UPR) target gene that controls apoptosis through the modulation of ER calcium homeostasis. Cell Death Differ. 2012, 19, 1013–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallmann, R.T.; Moravcikova, L.; Ondacova, K.; Lacinova, L.; Klugbauer, N. Grina/TMBIM3 modulates voltage-gated CaV2.2 Ca2+ channels in a G-protein-like manner. Cell Calcium 2019, 80, 71–78. [Google Scholar] [CrossRef]
- Ringsevjen, H.; Umbach Hansen, H.M.; Hussain, S.; Hvalby, Ø.; Jensen, V.; Walaas, S.I.; Davanger, S. Presynaptic increase in IP3 receptor type 1 concentration in the early phase of hippocampal synaptic plasticity. Brain Res. 2019, 1706, 125–134. [Google Scholar] [CrossRef]
- Yamaji, T.; Nishikawa, K.; Hanada, K. Transmembrane BAX Inhibitor Motif containing (TMBIM) family proteins perturbs a trans-Golgi network enzyme, Gb3 synthase, and reduces Gb3 biosynthesis. J. Biol. Chem. 2010, 285, 35505–35518. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.J.; Sodhi, J.S.; McGuffin, L.J.; Buxton, B.F.; Jones, D.T. Prediction and functional analysis of native disorder in proteins from the three kingdoms of life. J. Mol. Biol. 2004, 337, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Tsafou, K.; Tiwari, P.B.; Forman-Kay, J.D.; Metallo, S.J.; Toretsky, J.A. Targeting intrinsically disordered transcription factors: Changing the paradigm. J. Mol. Biol. 2018, 430, 2321–2341. [Google Scholar] [CrossRef] [PubMed]
- Van der Lee, R.; Buljan, M.; Lang, B.; Weatheritt, R.J.; Daughdrill, G.W.; Dunker, A.K.; Fuxreiter, M.; Gough, J.; Gsponer, J.; Jones, D.T.; et al. Classification of intrinsically disordered regions and proteins. Chem. Rev. 2014, 114, 6589–6631. [Google Scholar] [CrossRef] [PubMed]
- Elam, W.A.; Schrank, T.P.; Campagnolo, A.J.; Hilser, V.J. Evolutionary conservation of the polyproline II conformation surrounding intrinsically disordered phosphorylation sites. Protein Sci. 2013, 22, 405–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ball, L.J.; Kühne, R.; Schneider-Mergener, J.; Oschkinat, H. Recognition of proline-rich motifs by protein-protein-interaction domains. Angewandte Chem. Int. Ed. 2005, 44, 2852–2869. [Google Scholar] [CrossRef]
- Oates, M.E.; Romero, P.; Ishida, T.; Ghalwash, M.; Mizianty, M.J.; Xue, B.; Dosztányi, Z.; Uversky, V.N.; Obradovic, Z.; Kurgan, L.; et al. D2P2: Database of disordered protein predictions. Nucleic Acids Res. 2013, 41, D508–D516. [Google Scholar] [CrossRef] [PubMed]
- Amanchy, R.; Periaswamy, B.; Mathivanan, S.; Reddy, R.; Tattikota, S.G.; Pandey, A. A curated compendium of phosphorylation motifs. Nat. Biotechnol. 2007, 25, 285–286. [Google Scholar] [CrossRef]
- Schwartz, D.; Gygi, S.P. An iterative statistical approach to the identification of protein phosphorylation motifs from large-scale data sets. Nat. Biotechnol. 2005, 23, 1391–1398. [Google Scholar] [CrossRef]
- Naaby-Hansen, S.; Mandal, A.; Wolkowicz, M.J.; Sen, B.; Westbrook, V.A.; Shetty, J.; Coonrod, S.A.; Klotz, K.L.; Kim, Y.H.; Bush, L.A.; et al. CABYR, a novel calcium-binding tyrosine phosphorylation-regulated fibrous sheath protein involved in capacitation. Dev. Biol. 2002, 242, 236–254. [Google Scholar] [CrossRef]
- Holt, C. Unfolded phosphopolypeptides enable soft and hard tissues to coexist in the same organism with relative ease. Curr. Opin. Struct. Biol. 2013, 23, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Dong, Q.; Yang, J.; Zhang, Y. Recognizing metal and acid radical ion-binding sites by integrating ab initio modeling with template-based transferals. Bioinformatics 2016, 32, 3260–3269. [Google Scholar] [CrossRef] [PubMed]
- Maki, M.; Suzuki, H.; Shibata, H. Structure and function of ALG-2, a penta-EF-hand calcium-dependent adaptor protein. Sci. China Life Sci. 2011, 54, 770–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acquaah-Mensah, G.K.; Agu, N.; Khan, T.; Gardner, A. A regulatory role for the insulin- and BDNF-Linked RORA in the hippocampus: Implications for Alzheimer’s disease. J. Alzheimer Dis. 2015, 44, 827–838. [Google Scholar] [CrossRef] [PubMed]
- McGourty, C.A.; Akopian, D.; Walsh, C.; Gorur, A.; Werner, A.; Schekman, R.; Bautista, D.; Rape, M. Regulation of the CUL3 ubiquitin ligase by a calcium-dependent co-adaptor. Cell 2016, 167, 525–538. [Google Scholar] [CrossRef]
- Janowicz, A.; Michalak, M.; Krebs, J. Stress induced subcellular distribution of ALG-2, RBM22 and hSlu7. Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 1045–1049. [Google Scholar] [CrossRef]
- Suzuki, H.; Kawasaki, M.; Inuzuka, T.; Okumura, M.; Kakiuchi, T.; Shibata, H.; Wakatsuki, S.; Maki, M. Structural Basis for Ca2+-Dependent Formation of ALG-2/Alix Peptide Complex: Ca2+/EF3-Driven Arginine Switch Mechanism. Structure 2008, 16, 1562–1573. [Google Scholar] [CrossRef]
- Lania, G.; Nanayakkara, M.; Maglio, M.; Auricchio, R.; Porpora, M.; Conte, M.; De Matteis, M.A.; Rizzo, R.; Luini, A.; Discepolo, V.; et al. Constitutive alterations in vesicular trafficking increase the sensitivity of cells from celiac disease patients to gliadin. Commun. Biol. 2019, 2, 190. [Google Scholar] [CrossRef]
- Kurakin, A.; Bredesen, D.E. An unconventional IAP-binding motif revealed by target-assisted iterative screening (TAIS) of the BIR3-cIAP1 domain. J. Mol. Recognit. 2007, 20, 39–50. [Google Scholar] [CrossRef]
- Garcia-Quintanilla, A.; Miranzo-Navarro, D. Extraintestinal manifestations of celiac disease: 33-mer gliadin binding to glutamate receptor GRINA as a new explanation. BioEssays 2016, 38, 427–439. [Google Scholar] [CrossRef]
- Čiháková, D.; Eaton, W.W.; Talor, M.V.; Harkus, U.H.; Demyanovich, H.; Rodriguez, K.; Feldman, S.; Kelly, D.L. Gut permeability and mimicry of the Glutamate Ionotropic Receptor NMDA type Subunit Associated with protein 1 (GRINA) as potential mechanisms related to a subgroup of people with schizophrenia with elevated antigliadin antibodies (AGA IgG). Schizophr. Res. 2019, 208, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tsai, C.-C. Atrophin Proteins: An overview of a new class of nuclear receptor corepressors. Nucl. Recept. Signal. 2008, 6, e009. [Google Scholar] [CrossRef]
- Rauscher, S.; Baud, S.; Miao, M.; Keeley, F.W.W.; Pomès, R. Proline and glycine control protein self-organization into elastomeric or amyloid fibrils. Structure 2006, 14, 1667–1676. [Google Scholar] [CrossRef] [PubMed]
- Breusegem, S.Y.; Seaman, M.N.J. Genome-wide RNAi screen reveals a role for multipass membrane proteins in endosome-to-Golgi retrieval. Cell Rep. 2014, 9, 1931–1945. [Google Scholar] [CrossRef]
- Protter, D.S.W.; Rao, B.S.; Van Treeck, B.; Lin, Y.; Mizoue, L.; Rosen, M.K.; Parker, R. Intrinsically disordered regions can contribute promiscuous interactions to RNP granule assembly. Cell Rep. 2018, 30, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Glabe, C.G.; Clark, D. The sequence of the Arbacia punctulata bindin cDNA and implications for the structural basis of species-specific sperm adhesion and fertilization. Dev. Biol. 1991, 143, 282–288. [Google Scholar] [CrossRef]
- Glabe, C.G. Interaction of the sperm adhesive protein, bindin, with phospholipid vesicles. I. Specific association of bindin with gel-phase phospholipid vesicles. J. Cell Biol. 1985, 100, 794–799. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Hu, L.; Smith, T.F.; Goldberger, G. LFG: A candidate apoptosis regulatory gene family. Apoptosis 2009, 14, 1255–1265. [Google Scholar] [CrossRef]
- Thul, P.J.; Åkesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Björk, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, eaal3321. [Google Scholar] [CrossRef]
- Szilágyi, A.; Skolnick, J. Efficient prediction of nucleic acid binding function from low-resolution protein structures. J. Mol. Biol. 2006, 358, 922–933. [Google Scholar] [CrossRef] [PubMed]
- Haze, K.; Yoshida, H.; Yanagi, H.; Yura, T.; Mori, K. Mammalian transcription factor ATF6 is synthesized as a transmembrane protein and activated by proteolysis in response to endoplasmic reticulum stress. Mol. Biol. Cell 1999, 10, 3787–3799. [Google Scholar] [CrossRef] [PubMed]
- Simonis, N.; Rual, J.-F.; Carvunis, A.-R.; Tasan, M.; Lemmens, I.; Hirozane-Kishikawa, T.; Hao, T.; Sahalie, J.M.; Venkatesan, K.; Gebreab, F.; et al. Empirically controlled mapping of the Caenorhabditis elegans protein-protein interactome network. Nat. Methods 2009, 6, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Espenshade, P.J. SREBPs: Sterol-regulated transcription factors. J. Cell Sci. 2006, 119, 973–976. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Armstrong, C.M.; Bertin, N.; Ge, H.; Milstein, S.; Boxem, M.; Vidalain, P.O.; Han, J.D.J.; Chesneau, A.; Hao, T.; et al. A map of the interactome network of the metazoan C. elegans. Science 2004, 303, 540–543. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.J.; Cansizoglu, A.E.; Süel, K.E.; Louis, T.H.; Zhang, Z.; Chook, Y.M. Rules for nuclear localization sequence recognition by karyopherin beta 2. Cell 2006, 126, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Schipke, J.; Pohlmann, A.; Diestel, R.; Binz, A.; Rudolph, K.; Nagel, C.-H.; Bauerfeind, R.; Sodeik, B. The C terminus of the large tegument protein pUL36 contains multiple capsid binding sites that function differently during assembly and cell entry of herpes simplex virus. J. Virol. 2012, 86, 3682–3700. [Google Scholar] [CrossRef] [PubMed]
- Schlieker, C.; Korbel, G.A.; Kattenhorn, L.M.; Ploegh, H.L. A Deubiquitinating Activity Is Conserved in the Large Tegument Protein of the Herpesviridae. J. Virol. 2005, 79, 15582–15585. [Google Scholar] [CrossRef] [PubMed]
- Hou, N.S.; Gutschmidt, A.; Choi, D.Y.; Pather, K.; Shi, X.; Watts, J.L.; Hoppe, T.; Taubert, S. Activation of the endoplasmic reticulum unfolded protein response by lipid disequilibrium without disturbed proteostasis in vivo. Proc. Natl. Acad. Sci. USA 2014, 111, E2271–E2280. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Vought, B.W.; Satterlee, J.S.; Walker, A.K.; Jim Sun, Z.Y.; Watts, J.L.; DeBeaumont, R.; Mako Saito, R.; Hyberts, S.G.; Yang, S.; et al. An ARC/Mediator subunit required for SREBP control of cholesterol and lipid homeostasis. Nature 2006, 442, 700–704. [Google Scholar] [CrossRef]
- Do, D.N.; Schenkel, F.S.; Miglior, F.; Zhao, X.; Ibeagha-Awemu, E.M. Genome wide association study identifies novel potential candidate genes for bovine milk cholesterol content. Sci. Rep. 2018, 8, 13239. [Google Scholar] [CrossRef] [PubMed]
- Petrov, A.M.; Kasimov, M.R.; Zefirov, A.L. Brain cholesterol metabolism and its defects: Linkage to neurodegenerative diseases and synaptic dysfunction. Acta Nat. 2016, 8, 58–73. [Google Scholar] [CrossRef]
- Lu, M.; McComish, B.J.; Burdon, K.P.; Taylor, B.V.; Körner, H. The Association Between Vitamin D and Multiple Sclerosis Risk: 1,25(OH)2D3 Induces Super-Enhancers Bound by VDR. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Riek, A.E.; Oh, J.; Sprague, J.E.; Timpson, A.; de las Fuentes, L.; Bernal-Mizrachi, L.; Schechtman, K.B.; Bernal-Mizrachi, C. Vitamin D suppression of endoplasmic reticulum stress promotes an antiatherogenic monocyte/macrophage phenotype in type 2 diabetic patients. J. Biol. Chem. 2012, 287, 38482–38494. [Google Scholar] [CrossRef] [PubMed]
- Mark, K.A.; Dumas, K.J.; Bhaumik, D.; Schilling, B.; Davis, S.; Oron, T.R.; Sorensen, D.J.; Lucanic, M.; Brem, R.B.; Melov, S.; et al. Vitamin D promotes protein homeostasis and longevity via the stress response pathway genes skn-1, ire-1, and xbp-1. Cell Rep. 2016, 17, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Park, J.E.; Pichiah, P.B.T.; Cha, Y.-S. Vitamin D and metabolic diseases: Growing roles of vitamin D. J. Obes. Metab. Syndr. 2018, 27, 223–232. [Google Scholar] [CrossRef]
- Zhou, R.; Chun, R.F.; Lisse, T.S.; Garcia, A.J.; Xu, J.; Adams, J.S.; Hewison, M. Vitamin D and alternative splicing of RNA. J. Steroid Biochem. Mol. Biol. 2015, 148, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Meijer, H.A.; Bushell, M.; Hill, K.; Gant, T.W.; Willis, A.E.; Jones, P.; de Moor, C.H. A novel method for poly(A) fractionation reveals a large population of mRNAs with a short poly(A) tail in mammalian cells. Nucleic Acids Res. 2007, 35, e132. [Google Scholar] [CrossRef]
- Sapkota, D.; Lake, A.M.; Yang, W.; Yang, C.; Wesseling, H.; Guise, A.; Uncu, C.; Dalal, J.S.; Kraft, A.W.; Lee, J.M.; et al. Cell-type-specific profiling of alternative translation identifies regulated protein isoform variation in the mouse brain. Cell Rep. 2019, 26, 594–607. [Google Scholar] [CrossRef]
- Wu, J.Q.; Wang, X.; Beveridge, N.J.; Tooney, P.A.; Scott, R.J.; Carr, V.J.; Cairns, M.J. Transcriptome sequencing revealed significant alteration of cortical promoter usage and splicing in schizophrenia. PLoS ONE 2012, 7, e36351. [Google Scholar] [CrossRef]
- Bowden, N.A.; Scott, R.J.; Tooney, P.A. Altered gene expression in the superior temporal gyrus in schizophrenia. BMC Genom. 2008, 9, 199. [Google Scholar] [CrossRef] [PubMed]
- Pujato, M.; Kieken, F.; Skiles, A.A.; Tapinos, N.; Fiser, A. Prediction of DNA binding motifs from 3D models of transcription factors; identifying TLX3 regulated genes. Nucleic Acids Res. 2014, 42, 13500–13512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouillard, A.D.; Gundersen, G.W.; Fernandez, N.F.; Wang, Z.; Monteiro, C.D.; McDermott, M.G.; Ma’ayan, A. The harmonizome: A collection of processed datasets gathered to serve and mine knowledge about genes and proteins. Database 2016, 2016, baw100. [Google Scholar] [CrossRef] [PubMed]
- Kulakovskiy, I.; Levitsky, V.; Oshchepkov, D.; Bryzgalov, L.; Vorontsov, I.; Makeev, V. From binding motifs in chip-seq data to improved models of transcription factor binding sites. J. Bioinform. Comput. Biol. 2013, 11, 1340004. [Google Scholar] [CrossRef] [PubMed]
- Jolma, A.; Yan, J.; Whitington, T.; Toivonen, J.; Nitta, K.R.; Rastas, P.; Morgunova, E.; Enge, M.; Taipale, M.; Wei, G.; et al. DNA-binding specificities of human transcription factors. Cell 2013, 152, 327–339. [Google Scholar] [CrossRef] [PubMed]
- Seitz, V.; Butzhammer, P.; Hirsch, B.; Hecht, J.; Gütgemann, I.; Ehlers, A.; Lenze, D.; Oker, E.; Sommerfeld, A.; von der Wall, E.; et al. Deep sequencing of MYC DNA-binding sites in Burkitt lymphoma. PLoS ONE 2011, 6, e26837. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.H.; Li, Q.; Hu, H.; Ni, B.; Liu, X.; Huang, C.; Zhang, Z.Z.; Zhao, G. Transmembrane protein GRINA modulates aerobic glycolysis and promotes tumor progression in gastric cancer. J. Exp. Clin. Cancer Res. 2018, 37, 308. [Google Scholar] [CrossRef]
- Matys, V. TRANSFAC® and its module TRANSCompel®: Transcriptional gene regulation in eukaryotes. Nucleic Acids Res. 2006, 34, D108–D110. [Google Scholar] [CrossRef]
- Weirauch, M.T.; Yang, A.; Albu, M.; Cote, A.G.; Montenegro-Montero, A.; Drewe, P.; Najafabadi, H.S.; Lambert, S.A.; Mann, I.; Cook, K.; et al. Determination and inference of eukaryotic transcription factor sequence specificity. Cell 2014, 158, 1431–1443. [Google Scholar] [CrossRef]
- Drutel, G. ARNT2, a transcription factor for brain neuron survival? Eur. J. Neurosci. 1999, 11, 1545–1553. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Y.; Zhang, J.; Liu, Y.; Zhang, Y.; Su, Z. Retinoic acid receptor-related orphan receptor α stimulates adipose tissue inflammation by modulating endoplasmic reticulum stress. J. Biol. Chem. 2017, 292, 13959–13969. [Google Scholar] [CrossRef] [PubMed]
- Halbach, M.V.; Gispert, S.; Stehning, T.; Damrath, E.; Walter, M.; Auburger, G. Atxn2 Knockout and CAG42-Knock-in Cerebellum Shows Similarly Dysregulated Expression in Calcium Homeostasis Pathway. Cerebellum 2017, 16, 68–81. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, A.; Matsuzaki, Y.; Konno, A.; Hirai, H. Plasticity of the developmentally arrested staggerer cerebellum in response to exogenous RORα. Brain Struct. Funct. 2016, 221, 2879–2889. [Google Scholar] [CrossRef] [PubMed]
- Porcelli, S.; Lee, S.J.; Han, C.; Patkar, A.A.; Albani, D.; Jun, T.Y.; Pae, C.U.; Serretti, A. Hot Genes in Schizophrenia: How Clinical Datasets Could Help to Refine their Role. J. Mol. Neurosci. 2018, 64, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, S.; Stevens, J.R.; Billon, C.; Girardet, C.; Sitaula, S.; Leon, A.S.; Rao, D.C.; Skinner, J.S.; Rankinen, T.; Bouchard, C.; et al. Adropin: An endocrine link between the biological clock and cholesterol homeostasis. Mol. Metab. 2018, 8, 51–64. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, H.; Shima, A.; Uda, A.; Ezaki, H.; Michihara, A. The retinoic acid receptor-related orphan receptor a positively regulates tight junction protein claudin domain-containing 1 mRNA expression in human brain endothelial cells. J. Biochem. 2017, 161, 441–540. [Google Scholar] [CrossRef] [PubMed]
- Guillemyn, B.; Kayserili, H.; Demuynck, L.; Sips, P.; De Paepe, A.; Syx, D.; Coucke, P.J.; Malfait, F.; Symoens, S. A homozygous pathogenic missense variant broadens the phenotypic and mutational spectrum of CREB3L1-related osteogenesis imperfecta. Hum. Mol. Genet. 2019, 28, 1801–1809. [Google Scholar] [CrossRef] [PubMed]
- García, I.A.; Torres Demichelis, V.; Viale, D.L.; Di Giusto, P.; Ezhova, Y.; Polishchuk, R.S.; Sampieri, L.; Martinez, H.; Sztul, E.; Alvarez, C. CREB3L1-mediated functional and structural adaptation of the secretory pathway in hormone-stimulated thyroid cells. J. Cell Sci. 2017, 130, 4155–4167. [Google Scholar] [CrossRef] [Green Version]
- Saito, A. Physiological functions of endoplasmic reticulum stress transducer OASIS in central nervous system. Anat. Sci. Int. 2014, 89, 11–20. [Google Scholar] [CrossRef]
- Saito, A.; Kanemoto, S.; Kawasaki, N.; Asada, R.; Iwamoto, H.; Oki, M.; Miyagi, H.; Izumi, S.; Sanosaka, T.; Nakashima, K.; et al. Unfolded protein response, activated by OASIS family transcription factors, promotes astrocyte differentiation. Nat. Commun. 2012, 3, 967. [Google Scholar] [CrossRef] [Green Version]
- Asada, R.; Saito, A.; Kawasaki, N.; Kanemoto, S.; Iwamoto, H.; Oki, M.; Miyagi, H.; Izumi, S.; Imaizumi, K. The endoplasmic reticulum stress transducer OASIS is involved in the terminal differentiation of goblet cells in the large intestine. J. Biol. Chem. 2012, 287, 8144–8153. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.; Oda, A.; Kanai, Y.; Goitsuka, R. Germ cell-intrinsic requirement for the homeodomain transcription factor PKnox1/Prep1 in adult spermatogenesis. PLoS ONE 2018, 13, e0190702. [Google Scholar] [CrossRef] [PubMed]
- Ricci, S.; Viggiano, D.; Cimmino, I.; Perruolo, G.; Cabaro, S.; Liotti, A.; Fiory, F.; Spinelli, R.; Di Carlo, A.; Beguinot, F.; et al. Prep1 deficiency affects olfactory perception and feeding behavior by impairing BDNF-TrkB mediated neurotrophic signaling. Mol. Neurobiol. 2018, 55, 6801–6815. [Google Scholar] [CrossRef] [PubMed]
- Sicouri, L.; Pisati, F.; Pece, S.; Blasi, F.; Longobardi, E. Prep1 (Pknox1) transcription factor contributes to pubertal mammary gland branching morphogenesis. Int. J. Dev. Biol. 2018, 62, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Lindeman, R.E.; Gearhart, M.D.; Minkina, A.; Krentz, A.D.; Bardwell, V.J.; Zarkower, D. Sexual cell-fate reprogramming in the ovary by DMRT1. Curr. Biol. 2015, 25, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Wawrzik, M.; Spiess, A.N.; Herrmann, R.; Buiting, K.; Horsthemke, B. Expression of SNURF-SNRPN upstream transcripts and epigenetic regulatory genes during human spermatogenesis. Eur. J. Hum. Genet. 2009, 17, 1463–1470. [Google Scholar] [CrossRef]
- Berger, J.; Sansom, O.; Clarke, A.; Bird, A. MBD2 is required for correct spatial gene expression in the gut. Mol. Cell. Biol. 2007, 27, 4049–4057. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Song, J.; He, X.; Zhang, M.; Hu, S.; Zhang, S.; Yu, Q.; Yang, P.; Xiong, F.; Wang, D.W.; et al. Loss of Mbd2 protects mice against high-fat diet-induced obesity and insulin resistance by regulating the homeostasis of energy storage and expenditure. Diabetes 2016, 65, 3384–3395. [Google Scholar] [CrossRef]
- Riechmann, V.; Van Crüchten, I.; Sablitzky, F. The expression pattern of Id4, a novel dominant negative helix-loop-helix protein, is distinct from Id1, 1d2 and Id3. Nucleic Acids Res. 1994, 22, 749–755. [Google Scholar] [CrossRef]
- Bedford, L.; Walker, R.; Kondo, T.; Van Crüchten, I.; King, E.R.; Sablitzky, F. Id4 is required for the correct timing of neural differentiation. Dev. Biol. 2005, 280, 386–395. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.S.; Zhang, Y.H.; Cai, Q.Y.; Yao, Z.X. ID2: A negative transcription factor regulating oligodendroglia differentiation. J. Neurosci. Res. 2012, 90, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Tokuzawa, Y.; Yagi, K.; Yamashita, Y.; Nakachi, Y.; Nikaido, I.; Bono, H.; Ninomiya, Y.; Kanesaki-Yatsuka, Y.; Akita, M.; Motegi, H.; et al. Id4, a new candidate gene for senile osteoporosis, acts as a molecular switch promoting osteoblast differentiation. PLoS Genet. 2010, 6, e1001019. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Knowell, A.E.; Korang-Yeboah, M.; Sharma, P.; Joshi, J.; Glymph, S.; Chinaranagari, S.; Nagappan, P.; Palaniappan, R.; Bowen, N.J.; et al. Inhibitor of differentiation 4 (ID4) inactivation promotes de novo steroidogenesis and castration-resistant prostate cancer. Mol. Endocrinol. 2014, 28, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Kumar, S.; Lee, B.; Kim, K.J.; Seo, J.E.; Choi, H.S.; Lee, K. Estrogen receptor-related receptor γ regulates testicular steroidogenesis through direct and indirect regulation of steroidogenic gene expression. Mol. Cell. Endocrinol. 2017, 452, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Kubo, M.; Ijichi, N.; Ikeda, K.; Horie-Inoue, K.; Takeda, S.; Inoue, S. Modulation of adipogenesis-related gene expression by estrogen-related receptor γ during adipocytic differentiation. Biochim. Biophys. Acta Gene Regul. Mech. 2009, 1789, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Ijichi, N.; Shigekawa, T.; Ikeda, K.; Horie-Inoue, K.; Fujimura, T.; Tsuda, H.; Osaki, A.; Saeki, T.; Inoue, S. Estrogen-related receptor γ Modulates cell proliferation and estrogen signaling in breast cancer. J. Steroid Biochem. Mol. Biol. 2011, 123, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Choi, Y.K.; Do, J.Y.; Choi, Y.K.; Ha, C.M.; Lee, S.J.; Jeon, J.H.; Lee, W.K.; Choi, H.S.; Park, K.G.; et al. Estrogen-related receptor γ plays a key role in vascular calcification through the upregulation of BMP2 expression. Arterioscler Thromb. Vasc. Biol. 2015, 35, 2384–2390. [Google Scholar] [CrossRef] [PubMed]
- Elfassihi, L.; Giroux, S.; Bureau, A.; Laflamme, N.; Cole, D.E.C.; Rousseau, F. Association with replication between estrogen-related receptor γ (ESRRγ) polymorphisms and bone phenotypes in women of European ancestry. J. Bone Miner. Res. 2010, 25, 901–911. [Google Scholar]
- Misra, J.; Kim, D.K.; Choi, W.; Koo, S.H.; Lee, C.H.; Back, S.H.; Kaufman, R.J.; Choi, H.S. Transcriptional cross talk between orphan nuclear receptor ERRγ and transmembrane transcription factor ATF6α coordinates endoplasmic reticulum stress response. Nucleic Acids Res. 2013, 41, 6960–6974. [Google Scholar] [CrossRef]
- Schilit, S.L.P.; Currall, B.B.; Yao, R.; Hanscom, C.; Collins, R.L.; Pillalamarri, V.; Lee, D.Y.; Kammin, T.; Zepeda-Mendoza, C.J.; Mononen, T.; et al. Estrogen-related receptor gamma implicated in a phenotype including hearing loss and mild developmental delay. Eur. J. Hum. Genet. 2016, 24, 1622–1626. [Google Scholar] [CrossRef] [Green Version]
- Hermans-Borgmeyer, I.; Süsens, U.; Borgmeyer, U. Developmental expression of the estrogen receptor-related receptor γ in the nervous system during mouse embryogenesis. Mech. Dev. 2000, 97, 197–199. [Google Scholar] [CrossRef]
- Pei, L.; Mu, Y.; Leblanc, M.; Alaynick, W.; Barish, G.D.; Pankratz, M.; Tseng, T.W.; Kaufman, S.; Liddle, C.; Yu, R.T.; et al. Dependence of hippocampal function on ERRγ-regulated mitochondrial metabolism. Cell Metab. 2015, 21, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; McDonald, C.; Petrenko, N.B.; Leblanc, M.; Wang, T.; Giguere, V.; Evans, R.M.; Patel, V.V.; Pei, L. Estrogen-related receptor α (ERRα) and ERRγ are essential coordinators of cardiac metabolism and function. Mol. Cell. Biol. 2015, 35, 1281–1298. [Google Scholar] [CrossRef] [PubMed]
- Alaynick, W.A.; Way, J.M.; Wilson, S.A.; Benson, W.G.; Pei, L.; Downes, M.; Yu, R.; Jonker, J.W.; Holt, J.A.; Rajpal, D.K.; et al. ERRγ regulates cardiac, gastric, and renal potassium homeostasis. Mol. Endocrinol. 2009, 24, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Gibson, J.D.; Miyamoto, S.; Sail, V.; Verma, R.; Rosenberg, D.W.; Nelson, C.E.; Giardina, C. Intestinal lineage commitment of embryonic stem cells. Differentiation 2011, 81, 1–10. [Google Scholar] [CrossRef]
- Lescroart, F.; Wang, X.; Lin, X.; Swedlund, B.; Gargouri, S.; Sànchez-Dànes, A.; Moignard, V.; Dubois, C.; Paulissen, C.; Kinston, S.; et al. Defining the earliest step of cardiovascular lineage segregation by single-cell RNA-seq. Science 2018, 359, 1177–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Z.H.; Zhong, Y.; Chen, Y.-C.; Zhang, Z.-W. Research progress on the roles of Krüppel-like factors in muscle tissues. Yi Chuan Hered. 2018, 40, 733–748. [Google Scholar]
- Natesampillai, S.; Fernandez-Zapico, M.E.; Urrutia, R.; Veldhuis, J.D. A novel functional interaction between the Sp1-like protein KLF13 and SREBP-Sp1 activation complex underlies regulation of low density lipoprotein receptor promoter function. J. Biol. Chem. 2006, 281, 3040–3047. [Google Scholar] [CrossRef]
- Jeon, B.-N.; Kim, Y.-S.; Choi, W.-I.; Koh, D.-I.; Kim, M.-K.; Yoon, J.-H.; Kim, M.-Y.; Hur, B.; Paik, P.D.-H.; Hur, M.-W. Kr-pok increases FASN expression by modulating the DNA binding of SREBP-1c and Sp1 at the proximal promoter. J. Lipid Res. 2012, 53, 755–766. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, J. The novel function of HINFP as a co-activator in sterol-regulated transcription of PCSK9 in HepG2 cells. Biochem. J. 2012, 443, 757–768. [Google Scholar] [CrossRef]
- Ghule, P.N.; Xie, R.L.; Colby, J.L.; Rivera-Pérez, J.A.; Jones, S.N.; Lian, J.B.; Stein, J.L.; van Wijnen, A.J.; Stein, G.S. Maternal expression and early induction of histone gene transcription factor Hinfp sustains development in pre-implantation embryos. Dev. Biol. 2016, 419, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Fossum, S.L.; Mutolo, M.J.; Yang, R.; Dang, H.; O’Neal, W.K.; Knowles, M.R.; Leir, S.H.; Harris, A. Ets homologous factor regulates pathways controlling response to injury in airway epithelial cells. Nucleic Acids Res. 2014, 42, 13588–13598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, V.T.; Pio, R.; Jia, Z.; Mercola, D. Early Growth Response 3 regulates genes of inflammation and directly activates IL6 and IL8 expression in prostate cancer. Br. J. Cancer 2015, 112, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffenseller, B.; Kapczinski, F.; Gallitano, A.L.; Klamt, F. EGR3 immediate early gene and the brain-derived neurotrophic factor in bipolar disorder. Front. Behav. Neurosci. 2018, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Meyers, K.T.; Marballi, K.K.; Brunwasser, S.J.; Renda, B.; Charbel, M.; Marrone, D.F.; Gallitano, A.L. The immediate early gene Egr3 is required for hippocampal induction of Bdnf by electroconvulsive stimulation. Front. Behav. Neurosci. 2018, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.W.; Narayanan, R. The NF-κB transcription factor in oncogenesis. Anticancer Res. 1996, 16, 589–596. [Google Scholar] [PubMed]
- Qin, Y.; Hu, Q.; Xu, J.; Ji, S.; Dai, W.; Liu, W.; Xu, W.; Sun, Q.; Zhang, Z.; Ni, Q.; et al. PRMT5 enhances tumorigenicity and glycolysis in pancreatic cancer via the FBW7/cMyc axis. Cell Commun. Signal. 2019, 17, 30. [Google Scholar] [CrossRef]
- Casey, S.C.; Baylot, V.; Felsher, D.W. The MYC oncogene is a global regulator of the immune response. Blood 2018, 131, 2007–2015. [Google Scholar] [CrossRef] [Green Version]
- Van Riggelen, J.; Yetil, A.; Felsher, D.W. MYC as a regulator of ribosome biogenesis and protein synthesis. Nat. Rev. Cancer 2010, 10, 301–309. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Ogris, C.; Guala, D.; Mateusz, K.; Sonnhammer, E.L.L. FunCoup 4: New species, data, and visualization. Nucleic Acids Res. 2018, 46, D601–D607. [Google Scholar] [CrossRef] [PubMed]
- Kosyna, F.; Depping, R. Controlling the Gatekeeper: Therapeutic targeting of nuclear transport. Cells 2018, 7, 221. [Google Scholar] [CrossRef] [PubMed]
- Mahboubi, H.; Seganathy, E.; Kong, D.; Stochaj, U. Identification of novel stress granule components that are involved in nuclear transport. PLoS ONE 2013, 8, e68356. [Google Scholar] [CrossRef] [PubMed]
- Hanover, J.A.; Love, D.C.; Prinz, W.A. Calmodulin-driven nuclear entry: Trigger for sex determination and terminal differentiation. J. Biol. Chem. 2009, 284, 12593–12597. [Google Scholar] [CrossRef] [PubMed]
- Ben-Johny, M.; Yue, D.T. Calmodulin regulation (calmodulation) of voltage-gated calcium channels. J. Gen. Physiol. 2014, 143, 679–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mruk, K.; Farley, B.M.; Ritacco, A.W.; Kobertz, W.R. Calmodulation meta-analysis: Predicting calmodulin binding via canonical motif clustering. J. Gen. Physiol. 2014, 144, 105–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, C.H.; Tang, C.Y.; Pan, C.Y. Calmodulin modulates the Ca2+-dependent inactivation and expression level of bovine CaV2.2 expressed in HEK293T cells. IBRO Rep. 2017, 2, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Khanna, R.; Li, Q.; Bewersdorf, J.; Stanley, E.F. The presynaptic CaV2.2 channel-transmitter release site core complex. Eur. J. Neurosci. 2007, 26, 547–559. [Google Scholar]
- Thomas, J.R.; Hagen, J.; Soh, D.; Lee, A. Molecular moieties masking Ca2+-dependent facilitation of voltage-gated CaV2.2 Ca2+ channels. J. Gen. Physiol. 2018, 150, 83–94. [Google Scholar] [CrossRef]
- Kobayashi, H.; Saragai, S.; Naito, A.; Ichio, K.; Kawauchi, D.; Murakami, F. Calm1 signaling pathway is essential for the migration of mouse precerebellar neurons. J. Cell Sci. 2015, 142, 375–384. [Google Scholar]
- Iuliano, O.; Yoshimura, A.; Prospéri, M.T.; Martin, R.; Knölker, H.J.; Coudrier, E. Myosin 1b promotes axon formation by regulating actin wave propagation and growth cone dynamics. J. Cell Biol. 2018, 217, 2033–2046. [Google Scholar] [CrossRef]
- Salas-Cortes, L. Myosin Ib modulates the morphology and the protein transport within multi-vesicular sorting endosomes. J. Cell Sci. 2005, 118, 4823–4832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Růžičková, Š.; Staněk, D. Mutations in spliceosomal proteins and retina degeneration. RNA Biol. 2017, 14, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Dardenne, E.; PolayEspinoza, M.; Fattet, L.; Germann, S.; Lambert, M.P.; Neil, H.; Zonta, E.; Mortada, H.; Gratadou, L.; Deygas, M.; et al. RNA helicases DDX5 and DDX17 dynamically orchestrate transcription, miRNA, and splicing programs in cell differentiation. Cell Rep. 2014, 7, 1900–1913. [Google Scholar] [CrossRef] [PubMed]
- Germann, S.; Gratadou, L.; Zonta, E.; Dardenne, E.; Gaudineau, B.; Fougère, M.; Samaan, S.; Dutertre, M.; Jauliac, S.; Auboeuf, D. Dual role of the ddx5/ddx17 RNA helicases in the control of the pro-migratory NFAT5 transcription factor. Oncogene 2012, 31, 4536–4549. [Google Scholar] [CrossRef] [Green Version]
- Samaan, S.; Tranchevent, L.C.; Dardenne, E.; Polay Espinoza, M.; Zonta, E.; Germann, S.; Gratadou, L.; Dutertre, M.; Auboeuf, D. The Ddx5 and Ddx17 RNA helicases are cornerstones in the complex regulatory array of steroid hormone-signaling pathways. Nucleic Acids Res. 2014, 42, 2197–2207. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Yoon, J.G.; Li, L.; Tsai, Y.S.; Zheng, S.; Hood, L.; Goodlett, D.R.; Foltz, G.; Lin, B. Landscape of the SOX2 protein-protein interactome. Proteomics 2011, 11, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, H.; Gopal, K.; Gupta, N.; Jung, K.; Alshareef, A.; Ye, X.; Wu, F.; Li, L.; Lai, R. DDX17 (P72), a Sox2 binding partner, promotes stem-like features conferred by Sox2 in a small cell population in estrogen receptor-positive breast cancer. Cell. Signal. 2016, 28, 42–50. [Google Scholar] [CrossRef]
- Mai, S.; Qu, X.; Li, P.; Ma, Q.; Liu, X.; Cao, C. Functional interaction between nonreceptor tyrosine kinase c-Abl and SR-Rich protein RBM39. Biochem. Biophys. Res. Commun. 2016, 473, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Giot, L.; Bader, J.S.; Brouwer, C.; Chaudhuri, A.; Kuang, B.; Li, Y.; Hao, Y.L.; Ooi, C.E.; Godwin, B.; Vitols, E.; et al. A protein interaction map of Drosophila melanogaster. Science 2003, 302, 1727–1736. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, S.; Zheng, R.; Yue, F.; Lin, S.H.S.; Rahmeh, A.A.; Lee, E.Y.C.; Zhang, Z.; Lee, M.Y.W.T. PDIP46 (DNA polymerase δ; interacting protein 46) is an activating factor for human DNA polymerase δ. Oncotarget 2016, 7, 6294–6313. [Google Scholar] [CrossRef]
- Kroczynska, B.; Mehrotra, S.; Majchrzak-Kita, B.; Arslan, A.D.; Altman, J.K.; Stein, B.L.; McMahon, B.; Kozlowski, P.; Kahle, P.J.; Eklund, E.A.; et al. Regulatory effects of SKAR in interferon signaling and its role in the generation of type I IFN responses. Proc. Natl. Acad. Sci. USA 2014, 111, 11377–11382. [Google Scholar] [CrossRef] [PubMed]
- Richardson, C.J.; Bröenstrup, M.; Fingar, D.C.; Jülich, K.; Ballif, B.A.; Gygi, S.; Blenis, J. SKAR is a specific target of S6 kinase 1 in cell growth control. Curr. Biol. 2004, 14, 1540–1549. [Google Scholar] [CrossRef] [PubMed]
- Folco, E.G.; Lee, C.S.; Dufu, K.; Yamazaki, T.; Reed, R. The proteins PDIP3 and ZC11A associate with the human TREX complex in an ATP-dependent manner and function in mRNA export. PLoS ONE 2012, 7, e43804. [Google Scholar] [CrossRef] [PubMed]
- Shiga, A.; Ishihara, T.; Miyashita, A.; Kuwabara, M.; Kato, T.; Watanabe, N.; Yamahira, A.; Kondo, C.; Yokoseki, A.; Takahashi, M.; et al. Alteration of POLDIP3 splicing associated with loss of function of TDP-43 in tissues affected with ALS. PLoS ONE 2012, 7, e43120. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.; Feng, Y.; Manley, J.L. Dephosphorylated SRp38 acts as a splicing repressor in response to heat shock. Nature 2004, 427, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.; Manley, J.L. The SR protein SRp38 represses splicing in M phase cells. Cell 2002, 111, 407–417. [Google Scholar] [CrossRef]
- Sumter, T.F.; Xian, L.; Huso, T.; Koo, M.; Chang, Y.-T.; Almasri, T.N.; Chia, L.; Inglis, C.; Reid, D.; Resar, L.M.S. The high mobility group A1 (HMGA1) transcriptome in cancer and development. Curr. Mol. Med. 2016, 16, 353–393. [Google Scholar] [CrossRef]
- Kadomatsu, K.; Muramatsu, T. Midkine and pleiotrophin in neural development and cancer. Cancer Lett. 2004, 204, 127–143. [Google Scholar] [CrossRef]
- Muramatsu, H. α(4)β(1)-and α(6)β(1)-integrins are functional receptors for midkine, a heparin-binding growth factor. J. Cell Sci. 2004, 117, 5405–5415. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.C. Midkine translocated to nucleoli and involved in carcinogenesis. World J. Gastroenterol. 2009, 15, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.C.; Shao, J.Z.; Min, L.S.; Xiao, Y.T.; Xiang, L.X.; Ma, Z.H. Midkine accumulated in nucleolus of HepG2 cells involved in rRNA transcription. World J. Gastroenterol. 2008, 14, 6249–6253. [Google Scholar] [CrossRef] [PubMed]
- Shibata, Y.; Muramatsu, T.; Hirai, M.; Inui, T.; Kimura, T.; Saito, H.; McCormick, L.M.; Bu, G.; Kadomatsu, K. Nuclear targeting by the growth factor midkine. Mol. Cell. Biol. 2002, 22, 6788–6796. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Dunker, A.K.; Uversky, V.N.; Kurgan, L. Molecular recognition features (MoRFs) in three domains of life. Mol. Biosyst. 2016, 12, 697–710. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
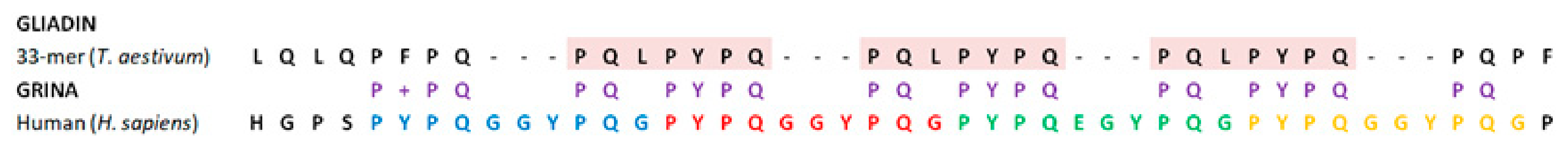{kind=link}
{kind=link}
{kind=link}
| ELM Name | Motif | Position | C.S.1 | Pattern & Description | Cell Compartment |
|---|---|---|---|---|---|
| CLV_NRD_NRD_1 | IRK RRK | 162–164 221–223 | 1.0 0.98 | (.RK)|(RR[^KR]) N-Arg dibasic convertase (NRD/Nardilysin) cleavage site (X-|-R-K or R-|-R-X) | extracellular, Golgi, cell surface |
| CLV_PCSK_SKI1_1 | RQAFI | 158–162 | 1.0 | [RK].[AILMFV][LTKF]. Subtilisin/kexin isozyme-1 (SKI1/S1P) cleavage site ([RK]-X-[hydrophobic]-[LTKF]-|-X) | ER, Golgi, extracellular |
| LIG_BIR_II_1 | MSHEK | 1–5 | 1.0 | ^M{0,1}[AS]... Inhibitor of Apoptosis (IAP)-binding motifs (IBM) are found in pro-apoptotic proteins and bind to type II BIR domains present in IAPs | cytosol, mitochondrion |
| LIG_EF_ALG2_ABM_1 | PYPQGGYP PYPQGGYP PYPQGGYP | 63–70 73–80 93–100 | 0.85 0.85 0.85 | P[PG]{0,1}YP.{1,6}Y[QS]{0,1}P This isoform-specific ALG2-binding motif binds to the EF hand domains of the proapoptotic Ca2+-binding ALG2 protein in a Ca2+-dependent manner | nucleus, cytosol, ESCRT I complex, endosome, ER |
| LIG_SH3_3 | NYPPPNP PPNPGYP GPQPPMP YAQPPYP EGYPQGP | 13–19 16–22 24–30 32–38 87–93 | 0.73 0.73 0.5 0.77 0.82 | ...[PV]..P This is the motif recognized by those SH3 domains with a non-canonical class I recognition specificity | plasma membrane, focal adhesion, cytosol |
| LIG_FHA_2 | PATNWDD MQTRYDF | 148–154 273–279 | 0.72 1.0 | ..(T)..[DE]. Phospho-Thr motif binding a subset of FHA domains that have a preference for an acidic amino acid at the pT+3 position | nucleus, replication fork |
| LIG_TRAF2_1 | SPEE | 340–343 | 1.0 | [PSAT].[QE]E Major TRAF2-binding consensus motif. Members of the tumor necrosis factor receptor (TNFR) superfamily initiate intracellular signaling by recruiting the C-domain of the TNFR-associated factors (TRAFs) through their cytoplasmic tails | cytosol |
| DOC_WW_Pin1_4 | LSLSPE | 337–342 | 1.0 | ...([ST])P. The Class IV WW domain interaction motif is recognised primarily by the Pin1 phosphorylation-dependent prolyl isomerase | nucleus, cytosol |
| MOD_CK2_1 | LSLSPEE | 337–343 | 1.0 | ...([ST])..E CK2 phosphorylation site | nucleus, cytosol |
| MOD_ProDKin_1 | LSLSPEE | 337–343 | 1.0 | ...([ST])P.. Pro-Directed Kinase (e.g., MAPK) phosphorylation site in higher eukaryotes | nucleus, cytosol |
| MOD_Plk_1 | DDKSIRQ | 153–159 | 0.96 | .[DNE][^PG][ST](([FYILMVW]..)|([^PEDGKN][FWYLIVM]).) Ser/Thr residue phosphorylated by the Plk1 kinase | nucleus, spindle, γ-tubulin complex, midbody, cytosol, kinetochore, nuclear condensin complex, cleavage furrow, nucleoplasm, microtubule organizing center |
| TRG_ER_diArg_1 | FRR | 220–222 | 1.0 | ([LIVMFYWPR]R[^YFWDE]{0,1}R)|(R[^YFWDE]{0,1}R[LIVMFYWPR]) The di-Arg ER retention motif is defined by two consecutive arginine residues (RR) or with a single residue insertion (RXR). The motif is completed by an adjacent hydrophobic/Arg residue which may be on either side of the Arg pair | cytosol, ER, ER-Golgi transport vesicle |
| Classification | TF | Description | References |
|---|---|---|---|
| Basic domain | ARNT2 | Aryl-hydrocarbon receptor nuclear translocator 2 | [84] |
| MYF6 | Myogenic factor 6 (Herculin) | [84] | |
| ID4 | Inhibitor of DNA binding 4, dominant negative helix-loop-helix protein | [85] | |
| CREB3L1 | cAMP responsive element binding protein 3-like 1 | [85] | |
| MESP1 | Mesoderm posterior bHLH transcription factor 1 | [85] | |
| c-MYC | Myc proto-oncogene | [86,87] | |
| Immunoglobulin fold | NFκB1 | Nuclear factor of kappa light polypeptide gene enhancer in B-cells 1 | [85,88] |
| Helix-turn-helix domain | HOXD8 | Homeobox D8 | [85] |
| EHF | ETS homologous factor | [84] | |
| PKNOX1 | PBX/knotted 1 homeobox 1 | [84] | |
| Zinc-coordinating DNA-binding domain | DMRT1 | Doublesex and mab-3 related transcription factor 1 | [89] |
| ZBTB6 | Zinc finger and BTB domain containing 6 | [88] | |
| ZBTB7C | Zinc finger and BTB domain containing 7C | [85] | |
| ESRRG | Estrogen-related receptor gamma | [84] | |
| MBD2 | Methyl-CpG binding domain protein 2 | [84] | |
| RORA | RAR-related orphan receptor A | [84] | |
| EGR3 | Early growth response 3 | [85] | |
| HINFP | Histone H4 transcription factor | [85] | |
| KLF13 | Kruppel-like factor 13 | [85] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiménez-González, V.; Ogalla-García, E.; García-Quintanilla, M.; García-Quintanilla, A. Deciphering GRINA/Lifeguard1: Nuclear Location, Ca2+ Homeostasis and Vesicle Transport. Int. J. Mol. Sci. 2019, 20, 4005. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20164005
Jiménez-González V, Ogalla-García E, García-Quintanilla M, García-Quintanilla A. Deciphering GRINA/Lifeguard1: Nuclear Location, Ca2+ Homeostasis and Vesicle Transport. International Journal of Molecular Sciences. 2019; 20(16):4005. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20164005
Chicago/Turabian StyleJiménez-González, Víctor, Elena Ogalla-García, Meritxell García-Quintanilla, and Albert García-Quintanilla. 2019. "Deciphering GRINA/Lifeguard1: Nuclear Location, Ca2+ Homeostasis and Vesicle Transport" International Journal of Molecular Sciences 20, no. 16: 4005. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20164005