Short and Long-Term Changes in Social Odor Recognition and Plasma Cytokine Levels Following Oxygen (16O) Ion Radiation Exposure

 , ,
, ,
Abstract
:1. Introduction
2. Results
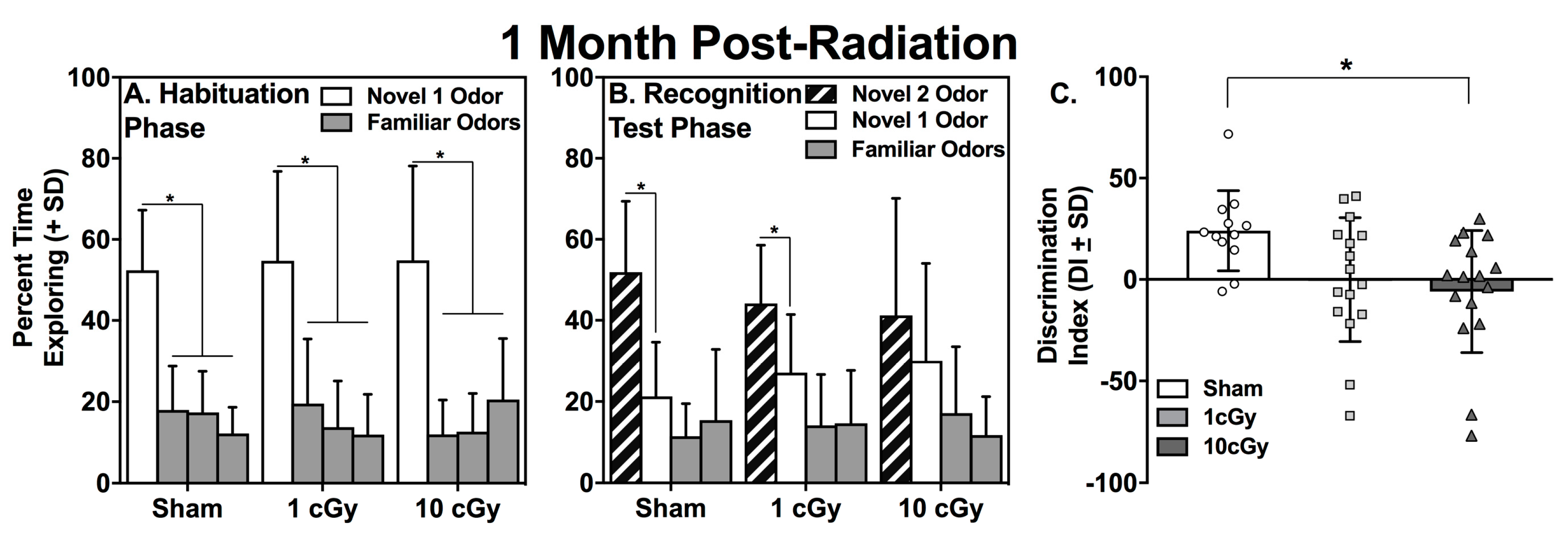
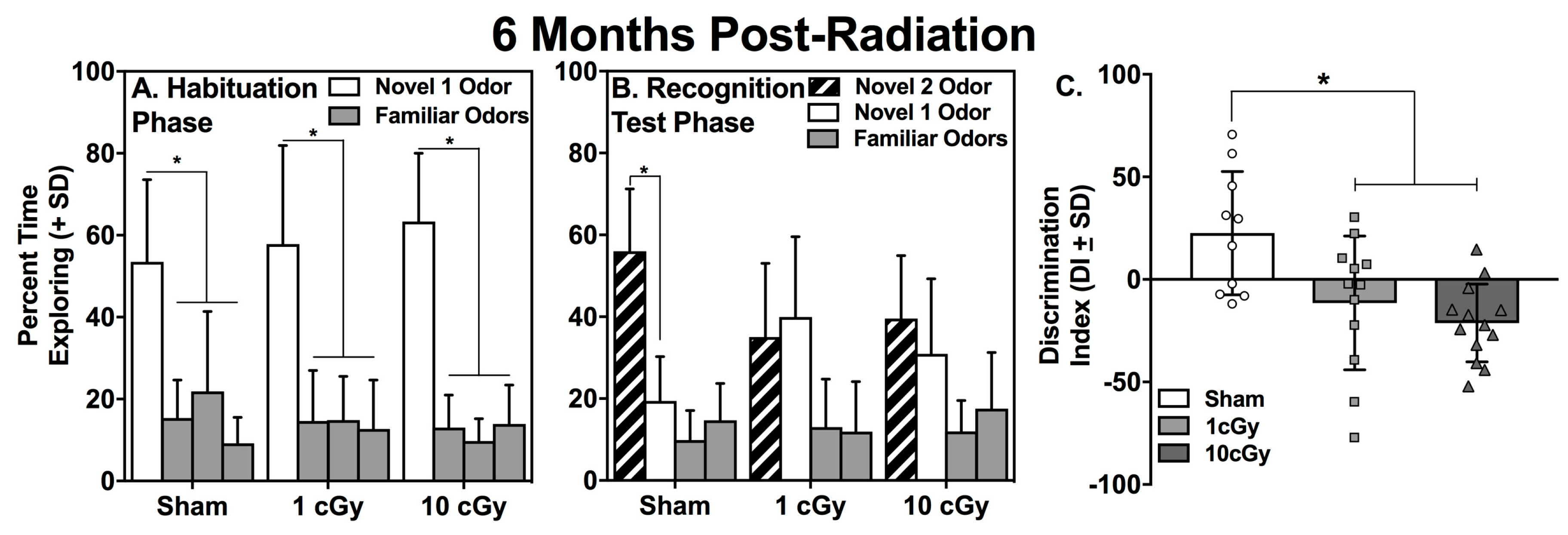
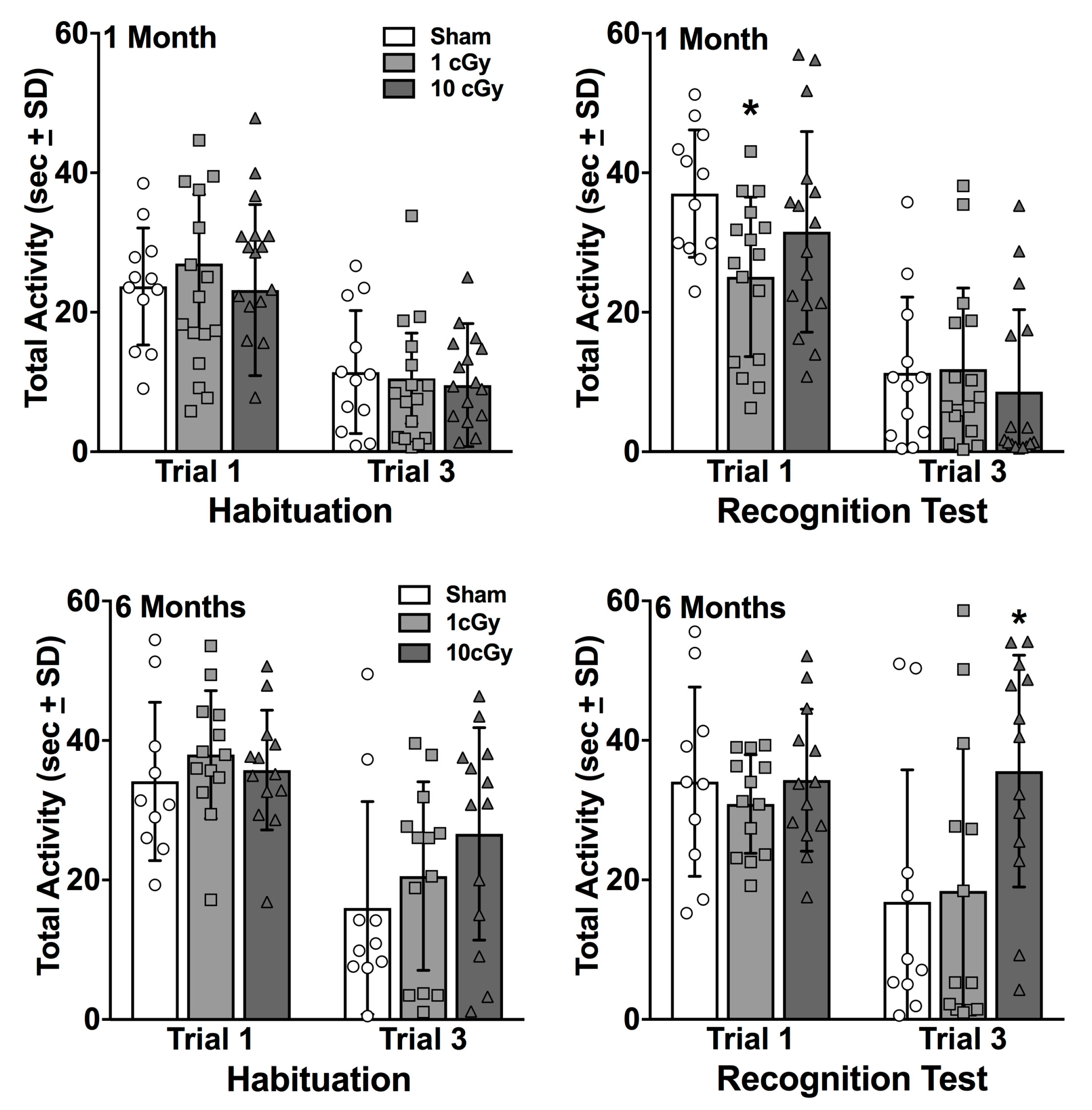
2.1. Habituation to the Novel 1 Odor is Unaffected by 16O Ion Exposure
2.2. Social Recognition is Dose-Dependently Impaired at One Month, but Not at Six Months, Following 16O Ion Exposure
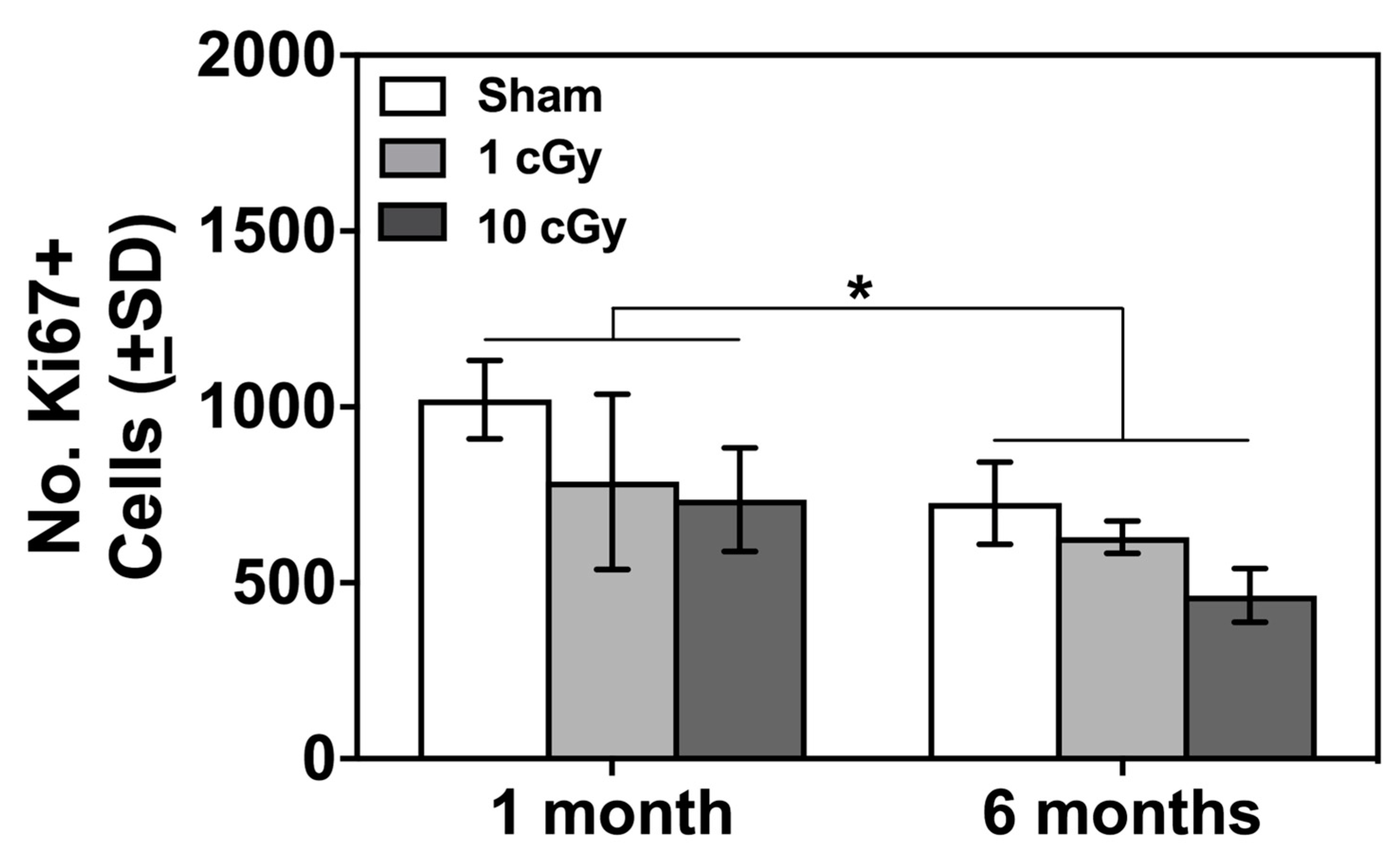
2.3. Ki67+ Cells in SVZ Unaffected by Radiation 16O Ion Exposure
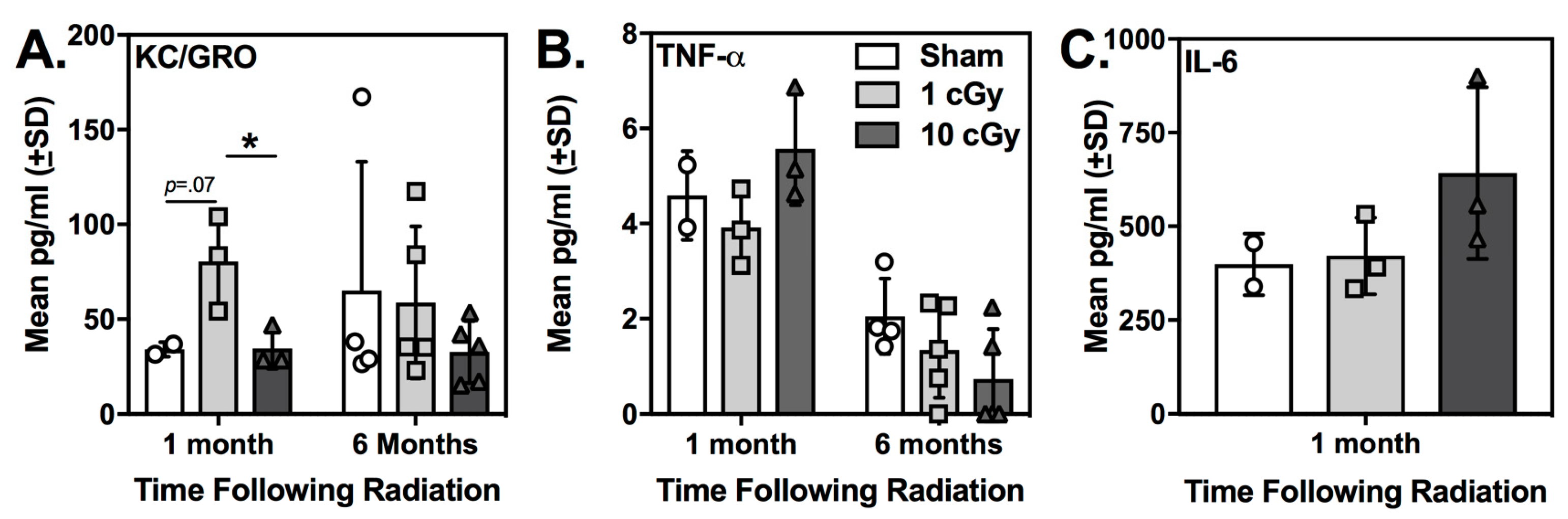
2.4. Plasma KC/GRO (CXCL1) was Elevated at One Month Following 16O Ion Exposure in 1 cGy-Exposed Rats
3. Discussion
4. Methods
4.1 Subjects and Apparatus
4.2. Radiation Procedures
4.3. Social Odor Recognition Memory Test (SORM)
4.4. Familiarization
4.5. Habituation
4.6. Recognition Test
4.7. Immunohistochemistry
4.8. Rat V-Plex Proinflammatory Cytokine Panel
4.9. Data Analysis
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Zeitlin, C.; Hassler, D.M.; Cucinotta, F.A.; Ehresmann, B.; Wimmer-Schweingruber, R.F.; Brinza, D.E.; Kang, S.; Weigle, G.; Böttcher, S.; Böhm, E.; et al. Measurements of energetic particle radiation in transit to Mars on the Mars Science Laboratory. Science 2013, 340, 1080–1084. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.A.; Townsend, L.W.; Norbury, J.W. Heavy ion contributions to organ dose equivalent for the 1977 galactic cosmic ray spectrum. Adv. Space Res. 2013, 51, 1792–1799. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Alp, M.; Sulzman, F.M.; Wang, M. Space radiation risks to the central nervous system. Life Sci. Space Res. 2014, 2, 54–69. [Google Scholar] [CrossRef] [Green Version]
- Parihar, V.K.; Allen, B.; Tran, K.K.; Macaraeg, T.G.; Chu, E.M.; Kwok, S.F.; Chmielewski, N.N.; Craver, B.M.; Baulch, J.E.; Acharya, M.M.; et al. What happens to your brain on the way to Mars. Sci. Adv. 2015, 1, e1400256. [Google Scholar] [CrossRef] [PubMed]
- Parihar, V.K.; Allen, B.D.; Caressi, C.; Kwok, S.; Chu, E.; Tran, K.K.; Chmielewski, N.N.; Giedzinski, E.; Acharya, M.M.; Britten, R.A.; et al. Cosmic radiation exposure and persistent cognitive dysfunction. Sci. Rep. 2016, 6, 34774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Impey, S.; Jopson, T.; Pelz, C.; Tafessu, A.; Fareh, F.; Zuloaga, D.; Marzulla, T.; Riparip, L.K.; Stewart, B.; Rosi, S.; et al. Short- and long-term effects of 56Fe irradiation on cognition and hippocampal DNA methylation and gene expression. BMC Genom. 2016, 17, 825. [Google Scholar] [CrossRef]
- Krukowski, K.; Feng, X.; Paladini, M.S.; Chou, A.; Sacramento, K.; Grue, K.; Riparip, L.K.; Jones, T.; Campbell-Beachler, M.; Nelson, G.; et al. Temporary microglia-depletion after cosmic radiation modifies phagocytic activity and prevents cognitive deficits. Sci. Rep. 2018, 8, 7857. [Google Scholar] [CrossRef] [Green Version]
- Krukowski, K.; Jones, T.; Campbell-Beachler, M.; Nelson, G.; Rosi, S. Peripheral T Cells as a Biomarker for Oxygen-Ion-Radiation-Induced Social Impairments. Radiat. Res. 2018, 190, 186–193. [Google Scholar] [CrossRef]
- Britten, R.A.; Davis, L.K.; Johnson, A.M.; Keeney, S.; Siegel, A.; Sanford, L.D.; Singletary, S.J.; Lonart, G. Low (20 cGy) doses of 1 GeV/u (56)Fe–particle radiation lead to a persistent reduction in the spatial learning ability of rats. Radiat. Res. 2012, 177, 146–151. [Google Scholar] [CrossRef]
- Britten, R.A.; Jewell, J.S.; Davis, L.K.; Miller, V.D.; Hadley, M.M.; Semmes, O.J.; Lonart, G.; Dutta, S.M. Changes in the Hippocampal Proteome Associated with Spatial Memory Impairment after Exposure to Low (20 cGy) Doses of 1 GeV/n (56)Fe Radiation. Radiat. Res. 2017, 187, 287–297. [Google Scholar] [CrossRef]
- Britten, R.A.; Jewell, J.S.; Miller, V.D.; Davis, L.K.; Hadley, M.M.; Wyrobek, A.J. Impaired Spatial Memory Performance in Adult Wistar Rats Exposed to Low (5-20 cGy) Doses of 1 GeV/n (56)Fe Particles. Radiat. Res. 2016, 185, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.A.; Miller, V.D.; Hadley, M.M.; Jewell, J.S.; Macadat, E. Performance in hippocampus- and PFC-dependent cognitive domains are not concomitantly impaired in rats exposed to 20cGy of 1GeV/n (56)Fe particles. Life Sci. Space Res. 2016, 10, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.A.; Wellman, L.; Sanford, L.D. Hadron-induced impairment of executive function: The exacerbating impact of sleep deprivation. In Proceedings of the 2017 NASA Human Research Program Investigators’ Workshop, Galveston, TX, USA, 23 January 2017. [Google Scholar]
- Limoli, C.; Parihar, V.K.; Baulch, J.E.; Acharya, M.M.; Vlkolinsky, R.; Nelson, G.; Britten, R.A.; Soltesz, I. Neurocognitive complications associated with exposure to cosmic radiation. In Proceedings of the NASA Human Research Program Investigators’ Workshop, Galveston, TX, USA, 23–26 January 2018. [Google Scholar]
- Poulose, S.M.; Bielinski, D.F.; Carrihill-Knoll, K.; Rabin, B.M.; Shukitt-Hale, B. Exposure to 16O-particle radiation causes aging-like decrements in rats through increased oxidative stress, inflammation and loss of autophagy. Radiat. Res. 2011, 176, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Poulose, S.M.; Carrihill-Knoll, K.L.; Ramirez, F.; Bielinski, D.F.; Heroux, N.; Shukitt-Hale, B. Acute Effects of Exposure to (56)Fe and (16)O Particles on Learning and Memory. Radiat. Res. 2015, 184, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.M.; Decicco-Skinner, K.L.; Roma, P.G.; Hienz, R.D. Individual Differences in Attentional Deficits and Dopaminergic Protein Levels following Exposure to Proton Radiation. Radiat. Res. 2014, 181, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.N.; Young, L.J.; Insel, T.R. The neuroendocrine basis of social recognition. Front. Neuroendocrinol. 2002, 23, 200–224. [Google Scholar] [CrossRef] [PubMed]
- Bicks, L.K.; Koike, H.; Akbarian, S.; Morishita, H. Prefrontal Cortex and Social Cognition in Mouse and Man. Front. Psychol. 2015, 6, 1805. [Google Scholar] [CrossRef]
- Millan, M.J.; Bales, K.L. Towards improved animal models for evaluating social cognition and its disruption in schizophrenia: The CNTRICS initiative. Neurosci. Biobehav. Rev. 2013, 37 Pt B, 2166–2180. [Google Scholar] [CrossRef]
- Davis, C.M.; Roma, P.G.; Armour, E.; Gooden, V.L.; Brady, J.V.; Weed, M.R.; Hienz, R.D. Effects of X-ray radiation on complex visual discrimination learning and social recognition memory in rats. PLoS ONE 2014, 9, e104393. [Google Scholar] [CrossRef]
- Mange, A.; Cao, Y.; Zhang, S.; Hienz, R.D.; Davis, C.M. Whole-body Oxygen (16O) Ion Exposure-Induced Impairments in Social Odor Recognition Memory in Rats are Dose and Time Dependent. Radiat. Res. 2018, 189, 292–299. [Google Scholar] [CrossRef]
- Spinetta, M.J.; Woodlee, M.T.; Feinberg, L.M.; Stroud, C.; Schallert, K.; Cormack, L.K.; Schallert, T. Alcohol-induced retrograde memory impairment in rats: Prevention by caffeine. Psychopharmacology 2008, 201, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, L.M.; Allen, T.A.; Ly, D.; Fortin, N.J. Recognition memory for social and non-social odors: Differential effects of neurotoxic lesions to the hippocampus and perirhinal cortex. Neurobiol. Learn. Memory 2012, 97, 7–16. [Google Scholar] [CrossRef]
- O’Dell, S.J.; Feinberg, L.M.; Marshall, J.F. A neurotoxic regimen of methamphetamine impairs novelty recognition as measured by a social odor-based task. Behav. Brain Res. 2011, 216, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Perez, E.C.; Rodgers, S.P.; Inoue, T.; Pedersen, S.E.; Leasure, J.L.; Gaber, M.W. Olfactory Memory Impairment Differs by Sex in a Rodent Model of Pediatric Radiotherapy. Front. Behav. Neurosci. 2018, 12, 158. [Google Scholar] [CrossRef] [PubMed]
- Gil-Perotin, S.; Haines, J.D.; Kaur, J.; Marin-Husstege, M.; Spinetta, M.J.; Kim, K.H.; Duran-Moreno, M.; Schallert, T.; Zindy, F.; Roussel, M.F.; et al. Roles of p53 and p27(Kip1) in the regulation of neurogenesis in the murine adult subventricular zone. Eur. J. Neurosci. 2011, 34, 1040–1052. [Google Scholar] [CrossRef]
- Hellstrom, N.A.; Bjork-Eriksson, T.; Blomgren, K.; Kuhn, H.G. Differential recovery of neural stem cells in the subventricular zone and dentate gyrus after ionizing radiation. Stem Cells 2009, 27, 634–641. [Google Scholar] [CrossRef]
- Lazarini, F.; Gabellec, M.M.; Moigneu, C.; de Chaumont, F.; Olivo-Marin, J.C.; Lledo, P.M. Adult neurogenesis restores dopaminergic neuronal loss in the olfactory bulb. J. Neurosci. 2014, 34, 14430–14442. [Google Scholar] [CrossRef] [PubMed]
- Krukowski, K.; Grue, K.; Frias, E.S.; Pietrykowski, J.; Jones, T.; Nelson, G.; Rosi, S. Female mice are protected from space radiation-induced maladaptive responses. Brain Behav. Immun. 2018, 74, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Casero, D.; Gill, K.; Sridharan, V.; Koturbash, I.; Nelson, G.; Hauer-Jensen, M.; Boerma, M.; Braun, J.; Cheema, A.K. Space-type radiation induces multimodal responses in the mouse gut microbiome and metabolome. Microbiome 2017, 5, 105. [Google Scholar] [CrossRef] [Green Version]
- Marples, B.; Joiner, M.C. The response of Chinese hamster V79 cells to low radiation doses: Evidence of enhanced sensitivity of the whole cell population. Radiat. Res. 1993, 133, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Marples, B.; Collis, S.J. Low-dose hyper-radiosensitivity: Past, present, and future. Int. J. Radiat. Oncol. Biol. Phys. 2008, 70, 1310–1318. [Google Scholar] [CrossRef] [PubMed]
- Suman, S.; Rodriguez, O.C.; Winters, T.A.; Fornace, A.J., Jr.; Albanese, C.; Datta, K. Therapeutic and space radiation exposure of mouse brain causes impaired DNA repair response and premature senescence by chronic oxidant production. Aging 2013, 5, 607–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raber, J.; Allen, A.R.; Sharma, S.; Allen, B.; Rosi, S.; Olsen, R.H.; Davis, M.J.; Eiwaz, M.; Fike, J.R.; Nelson, G.A. Effects of Proton and Combined Proton and (56)Fe Radiation on the Hippocampus. Radiat. Res. 2016, 185, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Raber, J.; Rudobeck, E.; Campbell-Beachler, M.; Allen, A.R.; Allen, B.; Rosi, S.; Nelson, G.A.; Ramachandran, S.; Turner, J.; Fike, J.R.; et al. (28)Silicon radiation-induced enhancement of synaptic plasticity in the hippocampus of naive and cognitively tested mice. Radiat. Res. 2014, 181, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Favre, C.J.; Fike, J.R.; Kubinova, L.; Anderson, E.; Campbell-Beachler, M.; Jones, T.; Smith, A.; Rightnar, S.; Nelson, G.A. High-LET radiation-induced response of microvessels in the Hippocampus. Radiat. Res. 2010, 173, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Kiffer, F.; Alexander, T.; Anderson, J.; Groves, T.; Wang, J.; Sridharan, V.; Boerma, M.; Allen, A. Late effects of 16O irradiation on social and cognitive behavior in female mice. In Proceedings of the NASA Human Research Program Investigators’ Workshop, Galveston, TX, USA, 23–26 January 2018. [Google Scholar]
- Whoolery, C.W.; Walker, A.K.; Richardson, D.R.; Lucero, M.J.; Reynolds, R.P.; Beddow, D.H.; Clark, K.L.; Shih, H.Y.; LeBlanc, J.A.; Cole, M.G.; et al. Whole-Body Exposure to 28Si-Radiation Dose-Dependently Disrupts Dentate Gyrus Neurogenesis and Proliferation in the Short Term and New Neuron Survival and Contextual Fear Conditioning in the Long Term. Radiat. Res. 2017, 188, 612–631. [Google Scholar] [CrossRef] [PubMed]
- Bordiuk, O.L.; Smith, K.; Morin, P.J.; Semenov, M.V. Cell proliferation and neurogenesis in adult mouse brain. PLoS ONE 2014, 9, e111453. [Google Scholar] [CrossRef] [PubMed]
- Mastrodonato, A.; Barbati, S.A.; Leone, L.; Colussi, C.; Gironi, K.; Rinaudo, M.; Piacentini, R.; Denny, C.A.; Grassi, C. Olfactory memory is enhanced in mice exposed to extremely low-frequency electromagnetic fields via Wnt/β-catenin dependent modulation of subventricular zone neurogenesis. Sci. Rep. 2018, 8, 262. [Google Scholar] [CrossRef] [PubMed]
- Lazarini, F.; Mouthon, M.A.; Gheusi, G.; de Chaumont, F.; Olivo-Marin, J.C.; Lamarque, S.; Abrous, D.N.; Boussin, F.D.; Lledo, P.M. Cellular and behavioral effects of cranial irradiation of the subventricular zone in adult mice. PLoS ONE 2009, 4, e7017. [Google Scholar] [CrossRef] [PubMed]
- Siopi, E.; Denizet, M.; Gabellec, M.M.; de Chaumont, F.; Olivo-Marin, J.C.; Guilloux, J.P.; Lledo, P.M.; Lazarini, F. Anxiety- and Depression-Like States Lead to Pronounced Olfactory Deficits and Impaired Adult Neurogenesis in Mice. J. Neurosci. 2016, 36, 518–531. [Google Scholar] [CrossRef] [Green Version]
- Zanni, G.; Deutsch, H.; Rivera, P.; Shih, H.-Y.; LeBlanc, J.; Amaral, W.; Lucero, M.; Redfield, R.; DeSalle, M.; Chen, B.; et al. Whole-Body 12C Irradiation Transiently Decreases Mouse Hippocampal Dentate Gyrus Proliferation and Immature Neuron Number, but Does Not Change New Neuron Survival Rate. Int. J. Mol. Sci. 2018, 19, 3078. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Meaney, M.J.; Duman, R.S.; Kaffman, A. Affiliative Behavior Requires Juvenile, But Not Adult Neurogenesis. J. Neurosci. 2011, 31, 14335–14345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omari, K.M.; Lutz, S.E.; Santambrogio, L.; Lira, S.A.; Raine, C.S. Neuroprotection and remyelination after autoimmune demyelination in mice that inducibly overexpress CXCL1. Am. J. Pathol. 2009, 174, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Kalm, M.; Fukuda, A.; Fukuda, H.; Ohrfelt, A.; Lannering, B.; Bjork-Eriksson, T.; Blennow, K.; Marky, I.; Blomgren, K. Transient inflammation in neurogenic regions after irradiation of the developing brain. Radiat. Res. 2009, 171, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Sanzari, J.K.; Nuth, M.; Kennedy, A.R. Induction of cytokine gene expression in human thyroid epithelial cells irradiated with HZE particles (iron ions). Radiat. Res. 2009, 172, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Ko, J. Neuroanatomical substrates of rodent social behavior: The medial prefrontal xrtex an its projection patterns. Front Neural Circuits 2017, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Beery, A.K.; Kaufer, D. Stress, social behavior, and resilience: Insights from rodents. Neurobiol. Stress 2015, 1, 116–127. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N1 Odor Trial 1 (s) | Familiar Odors Trial 1 (s) | t value | p value | N1 Odor Trial 3 (s) | t value | p value |
|---|---|---|---|---|---|---|---|
| Sham | 14.9 ± 2.85 | 3.63 ± 0.38 | 3.959 | 0.002 | 5.12 ± 1.95 | 2.790 | 0.024 |
| 1 cGy | 16.01 ± 2.8 | 3.65 ± 0.43 | 4.092 | 0.001 | 4.56 ± 1.41 | 4.607 | <0.001 |
| 10 cGy | 14.54 ± 2.85 | 2.88 ± 0.34 | 4.095 | 0.001 | 4.22 ± 2.03 | 4.494 | 0.001 |
| Group | N1 Odor Trial 1 (s) | Familiar Odors Trial 1 (s) | t value | p value | N1 Odor Trial 3 (s) | t value | p value |
|---|---|---|---|---|---|---|---|
| Sham | 19.40 ± 3.80 | 4.89 ± 0.49 | 3.637 | 0.005 | 4.50 ± 2.10 | 3.140 | 0.012 |
| 1 cGy | 22.57 ± 3.24 | 5.14 ± 0.72 | 4.669 | 0.001 | 7.78 ± 2.07 | 4.175 | 0.001 |
| 10 cGy | 23.28 ± 2.67 | 4.16 ± 0.52 | 6.463 | <0.001 | 12.8 ± 3.28 | 2.581 | 0.024 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, C.B.; Mange, A.; Granata, L.; Johnson, B.; Hienz, R.D.; Davis, C.M. Short and Long-Term Changes in Social Odor Recognition and Plasma Cytokine Levels Following Oxygen (16O) Ion Radiation Exposure. Int. J. Mol. Sci. 2019, 20, 339. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020339
Jones CB, Mange A, Granata L, Johnson B, Hienz RD, Davis CM. Short and Long-Term Changes in Social Odor Recognition and Plasma Cytokine Levels Following Oxygen (16O) Ion Radiation Exposure. International Journal of Molecular Sciences. 2019; 20(2):339. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020339
Chicago/Turabian StyleJones, Carli B., Ami Mange, Lauren Granata, Benjamin Johnson, Robert D. Hienz, and Catherine M. Davis. 2019. "Short and Long-Term Changes in Social Odor Recognition and Plasma Cytokine Levels Following Oxygen (16O) Ion Radiation Exposure" International Journal of Molecular Sciences 20, no. 2: 339. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20020339