GLABRA2, a Common Regulator for Epidermal Cell Fate Determination and Anthocyanin Biosynthesis in Arabidopsis

1
College of Life Science, Linyi University, Linyi 276005, China
2
Key Laboratory of Molecular Epigenetics of MOE and Institute of Genetics & Cytology, Northeast Normal University, Changchun 130024, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(20), 4997; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20204997
Submission received: 10 August 2019
/
Revised: 24 September 2019
/
Accepted: 30 September 2019
/
Published: 9 October 2019
(This article belongs to the Special Issue Cell-Specificity in Plants)
Abstract
:Epidermal cell fate determination—including trichome initiation, root hair formation, and flavonoid and mucilage biosynthesis in Arabidopsis (Arabidopsis thaliana)—are controlled by a similar transcriptional regulatory network. In the network, it has been proposed that the MYB-bHLH-WD40 (MBW) activator complexes formed by an R2R3 MYB transcription factor, a bHLH transcription factor and the WD40-repeat protein TRANSPARENT TESTA GLABRA1 (TTG1) regulate the expression of downstream genes required for cell fate determination, flavonoid or mucilage biosynthesis, respectively. In epidermal cell fate determination and mucilage biosynthesis, the MBW activator complexes activate the expression of GLABRA2 (GL2). GL2 is a homeodomain transcription factor that promotes trichome initiation in shoots, mucilage biosynthesis in seeds, and inhibits root hair formation in roots. The MBW activator complexes also activate several R3 MYB genes. The R3 MYB proteins, in turn, competing with the R2R3 MYBs for binding bHLH transcription factors, therefore inhibiting the formation of the MBW activator complexes, lead to the inhibition of trichome initiation in shoots, and promotion of root hair formation in roots. In flavonoid biosynthesis, the MBW activator complexes activate the expression of the late biosynthesis genes in the flavonoid pathway, resulting in the production of anthocyanins or proanthocyanidins. Research progress in recent years suggests that the transcriptional regulatory network that controls epidermal cell fate determination and anthocyanin biosynthesis in Arabidopsis is far more complicated than previously thought. In particular, more regulators of GL2 have been identified, and GL2 has been shown to be involved in the regulation of anthocyanin biosynthesis. This review focuses on the research progress on the regulation of GL2 expression, and the roles of GL2 in the regulation of epidermal cell fate determination and anthocyanin biosynthesis in Arabidopsis.
1. Introduction
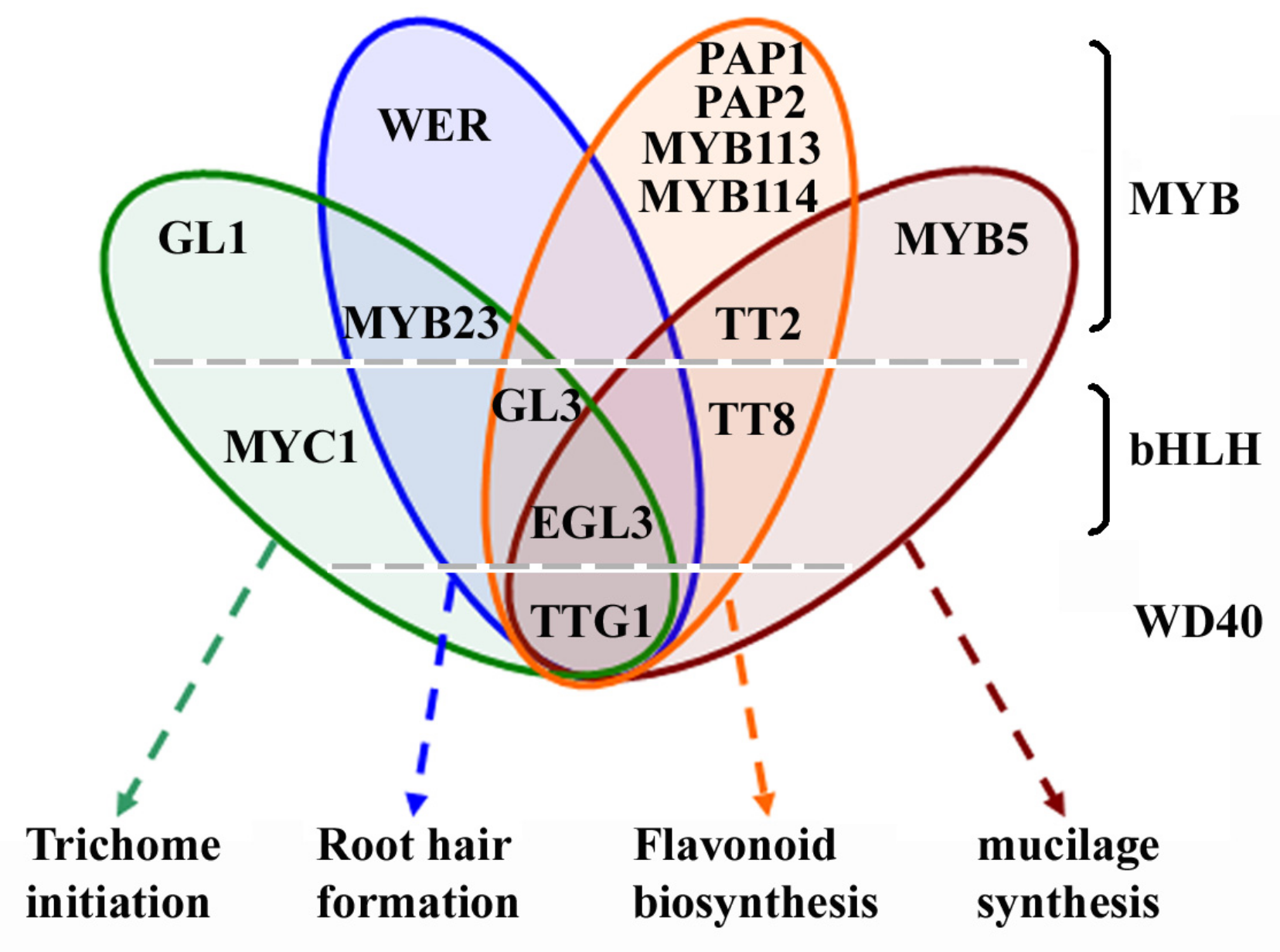
Interplay of several different types of transcription factors, including the WD40-repeat protein TRANSPARENT TESTA GLABRA1 (TTG1) [1,2], the R2R3 MYB transcription factor GLABRA1 (GL1), MYB23 and WEREWOLF (WER) [3,4,5], the bHLH (basic helix-loop-helix) transcription factors MYC1, GLABRA3 (GL3) and ENHANCER OF GL3 (EGL3) [6,7,8,9,10] regulates epidermal cell fate determination, including trichome initiation and root hair formation in Arabidopsis (Arabidopsis thaliana). Based on the characterization of gain-, and/or loss-of-function mutants of these transcription factors genes, and protein–protein interaction assays in yeast and plant cells, it has been proposed that these transcription factors form multiple MYB-bHLH-WD40 (MBW) transcription activator complexes to regulate trichome initiation and root hair formation [6,11,12,13]. The WD40 protein TTG1, the bHLH transcription factors GL3 and EGL3 and the R2R3 MYB transcription factor MYB23 are presented in all these MBW activator complexes, but some of the R2R3 MYB transcription factors involved in the trichome initiation and root hair formation regulating MBW complexes are different, i.e., WER specifically regulates root hair formation, whereas GL1 specifically regulate trichome formation (Figure 1). Unlike GL3 and EGL3, the bHLH transcription factor MYC1 has only been reported to be involved in the regulation of trichome initiation [6].
Biosynthesis of flavonoids including anthocyanin and proanthocyanidin in Arabidopsis is regulated by similar MBW activator complexes (Figure 1). Some of the components in the MBW activator complexes, including the WD40-repeat protein TTG1, the bHLH transcription factors GL3 and EGL3 are the same ones involved in the regulation of trichome initiation and root hair formation, whereas others are not. An additional bHLH transcription factor, TRANSPARENT TESTA 8 (TT8) is involved in the regulation of flavonoid biosynthesis, and the R2R3 MYB transcription factors that involved in the regulation of flavonoid biosynthesis are TT2, PRODUCTION OF ANTHOCYANIN PIGMENT 1 (PAP1), PAP2, MYB113 and MYB114 [10,12,14,15,16,17,18,19,20]. It should be noted that TTG1, EGL3, TT8, and TT2 are also regulators of mucilage biosynthesis [13], but another R2R3 MYB transcription factor, MYB5, specifically regulates mucilage biosynthesis [21]. In addition to mucilage biosynthesis, MYB5 is also involved in the regulation of trichome development [22].
In the process of trichome initiation and root hair formation, the MBW activator complexes activate the expression of GLABRA2 (GL2), a subfamily IV (HD-GLABRA2 group) homeodomain leucine zipper (HD-ZIP) protein gene [23,24], leading to promotion of trichome initiation in shoots, and inhibition of root hair formation in roots [11,12,13,25,26]. It should be mentioned that there are a total of four subfamilies of HD-ZIP proteins in Arabidopsis, and these proteins regulate multiple aspects of plant growth and development, as well as plant response to environmental stimuli [27,28].
The same MBW activator complexes can also activate the expression of single-repeat R3 MYB genes including TRY (TRIPTYCHON), CPC (CAPRICE), ETC1 (ENHANCER OF TRY AND CPC 1) and ETC3. The R3 MYB proteins, in turn, play an lateral inhibition role by competing with R2R3 MYB transcription factors GL1, MYB23 or WER for binding bHLH proteins GL3 or EGL3, thus blocking the formation of the MBW activator complexes, resulting in inhibition of trichome initiation and promotion of root hair formation [11,12,13,25]. In the process of flavonoid biosynthesis, however, the MBW activator complexes directly regulate the expression of late biosynthesis genes in the flavonoid biosynthesis pathway including FLAVANONE 3’-HYDROXYLASE (F3’H), DIHYDROFLAVONOL REDUCTASE (DFR), ANTHOCYANIDIN SYNTHASE/LEUCOANTHOCYANIDIN DIOXYGENASE (ANS/LDOX) and UDP-FLAVONOID GLUCOSYL TRANSFERASE (UFGT), leading to the accumulation of anthocyanins [12,16,17,18,20]. Whereas activation of BANYULS (BAN) by the MBW activator complexes leading to the production of proanthocyanidins [14].
Remarkable achievements have been made in recent years in the dissection of the transcriptional regulatory networks controlling epidermal cell fate determination and anthocyanin biosynthesis in Arabidopsis. In particular, GL2, the central regulator for epidermal cell fate determination has also been shown to be involved in the regulation of anthocyanin biosynthesis, and more regulators of GL2 have been identified. Focus of the present review is on the regulation of GL2, and the roles of GL2 in the regulation of epidermal cell fate determination and anthocyanin biosynthesis.
2. Regulation of GL2
As mentioned above, the MBW activator complexes formed by the WD40-repeat protein TTG1, the bHLH transcription factors GL3 or EGL3, and the R2R3 MYB transcription factors GL1, MYB23 or WER regulate the expression of GL2 [23,24]. In addition to these transcription factors, epigenetic modification of the chromosome by histone chaperones, histone deacetylase and histone acetyltransferase is involved in the regulation of GL2 expression (Figure 2).
2.1. Transcriptional Regulation of GL2 by Transcription Factors
In Arabidopsis, it has been proposed that the MBW activator complexes formed by the WD40 protein TTG1, the R2R3 MYB transcription factor GL1 or MYB23, and the bHLH transcription GL3 or EGL3 regulate trichome initiation [9,13,25,26,27,28,29,30,31,32,33], whereas MBW activator complexes formed by the WD40 protein TTG1, the R2R3 MYB transcription factor WER, and the R2R3 MYB transcription factor GL3 or EGL3 regulate root hair formation [4,13,30,31,32,33,34,35,36]. Although the R2R3 MYB transcription factors in these MBW activator complexes are different, the MBW activator complexes regulate the expression of the same homeodomain protein gene, GL2 [11,12,25]. Physically interaction of GL3/EGL3 with TTG1 and GL1 respectively, has been demonstrated in both yeast and plant cells [9,10,37,38]. The regulation of GL2 by the MBW activator complexes is supported by several different lines of evidence. First, the expression of the GL2 reporter gene PGL2:GUS was reduced in the ttg1, gl1 and gl3 mutants [7,39]. Second, ectopic expression of GL1 or R, a GL3 homologous gene in maize, in the ttg1 mutants ectopically activated GL2 expression [39]. Third, both GL1 and GL3 were found to bind to the promoter region of GL2 in Chromatin Immunoprecipitation (ChIP) assays, indicating that GL2 is a directly target of GL1 and GL3 [38,40].
Transient transfection assays in Arabidopsis mesophyll protoplasts, however, have shed new lights to the regulation of GL2 expression. Consistent with the proposal that GL2 is activated by the MBW activator complexes, transfection of TTG1, GL1, WER, GL3, or EGL3 alone into Arabidopsis protoplasts failed to activate the expression of the endogenous GL2 as detected by RT-PCR, and the expression of the PGL2:GUS reporter gene as detected by GUS activity assays [41]. On the other hand, co-transfection of GL1 or WER and GL3 activated both endogenous GL2 and the PGL2:GUS reporter gene [41]. In agreement with this, overexpression of a GL1GL3 fusion protein activated the expression of the PGL2:GUS reporter gene in the stable transformed transgenic plants [42]. Co-transfection of a GL1 or WER and GL3 or EGL3 also activated some of the R3 MYB genes, including TRY, CPC, ETC1 and ETC3, but not TRICHOMELESS1 (TCL1), TCL2, and ETC2 [43,44]. These results suggest that a complex formed by a R2R3 MYB transcription factor and a bHLH transcription factor is required and is sufficient to activate the expression of both GL2 (Figure 2) and some of the R3 MYB genes. Consistent with this, all the DNA fragments in the GL2 promoter region that can drive the proper expression of the GUS reporter gene contained at least one putative MYB-binding element and one putative bHLH binding-element [39,41]. In addition, previously experiments have shown that ectopic expression of GL1 or a GL3 homologous gene R in the ttg1 mutants ectopically activated GL2 [39], which also indicated that TTG1 is not required for the activation of GL2. However, a recent study reported that interaction of TTG1 with TT2 is required for the proper expression of GL2 [45], suggesting that TTG1 may still play a role in the regulation of GL2.
By using transient transfection assays in protoplast with truncated protein, Wang and Chen [41] further showed that both the R2 domain in GL1 and the C-terminal domain in GL3 are required for the binding of GL2 promoter, suggesting that a concurrent binding of GL1 and GL3 may be required for the activation of GL2. The R3 domain, on the other hand, has been shown to be required for the interaction of MYBs with bHLH transcription factors in yeast two hybridization assays [37]. Detailed analysis have identified [D/E]L×2[R/K]×3L×6L×3R in the R3 domain, as a conserved amino acid signature required for the interaction of MYBs with bHLH transcription factors [37]. As a matter of fact, this amino acid signature is conserved in GL1, MYB23, WER and all the Arabidopsis R3 MYB proteins [13,42,43]. In other plants such as poplar and rice, the [D/E]L×2[R/K]×3L×6L×3R amino acid signature is not fully conserved in the R3 MYBs. However, ectopic expression any of these R3 MYB genes resulted in inhibition of trichome initiation, and down regulation of GL2 [46,47]. Assays in transfected protoplast also showed that all the rice and poplar R3 MYBs interacted with GL3/EGL3, indicating that substitution of at least some of the conserved amino acid residues in the [D/E]L×2[R/K]×3L×6L×3R amino acid signature does not affect the interaction of R3 MYB with bHLH transcription factors [46,47].
Characterization of gl1-S92F, a loss-of-function mutant shed additional lights into the regulation of GL2 [42]. In the gl1-S92F mutant, a single nucleotide mutation in the GL1 gene resulted in a substitution of the Ser92 in the [D/E]L×2[R/K]×3L×6L×3R amino acid signature in the R3 domain of GL1 with a Phe (S92F). Protoplast transient transfection assays indicate that the interaction of GL1 and GL3 was not affected by the S92F amino acid substitution, however, contransfection of GL1-S92F and GL3 failed to activate the expression of GL2, and expression of GL2 in the gl1-S92F mutant was reduced when compared with that in the Col wild-type plants [42]. These results suggest that in addition to bind bHLH transcription factors, the R3 domain in GL1 is involved in the binding of GL1 to the promoter of GL2.
2.2. Regulation of GL2 by Epigenetic Modification of Chromosome
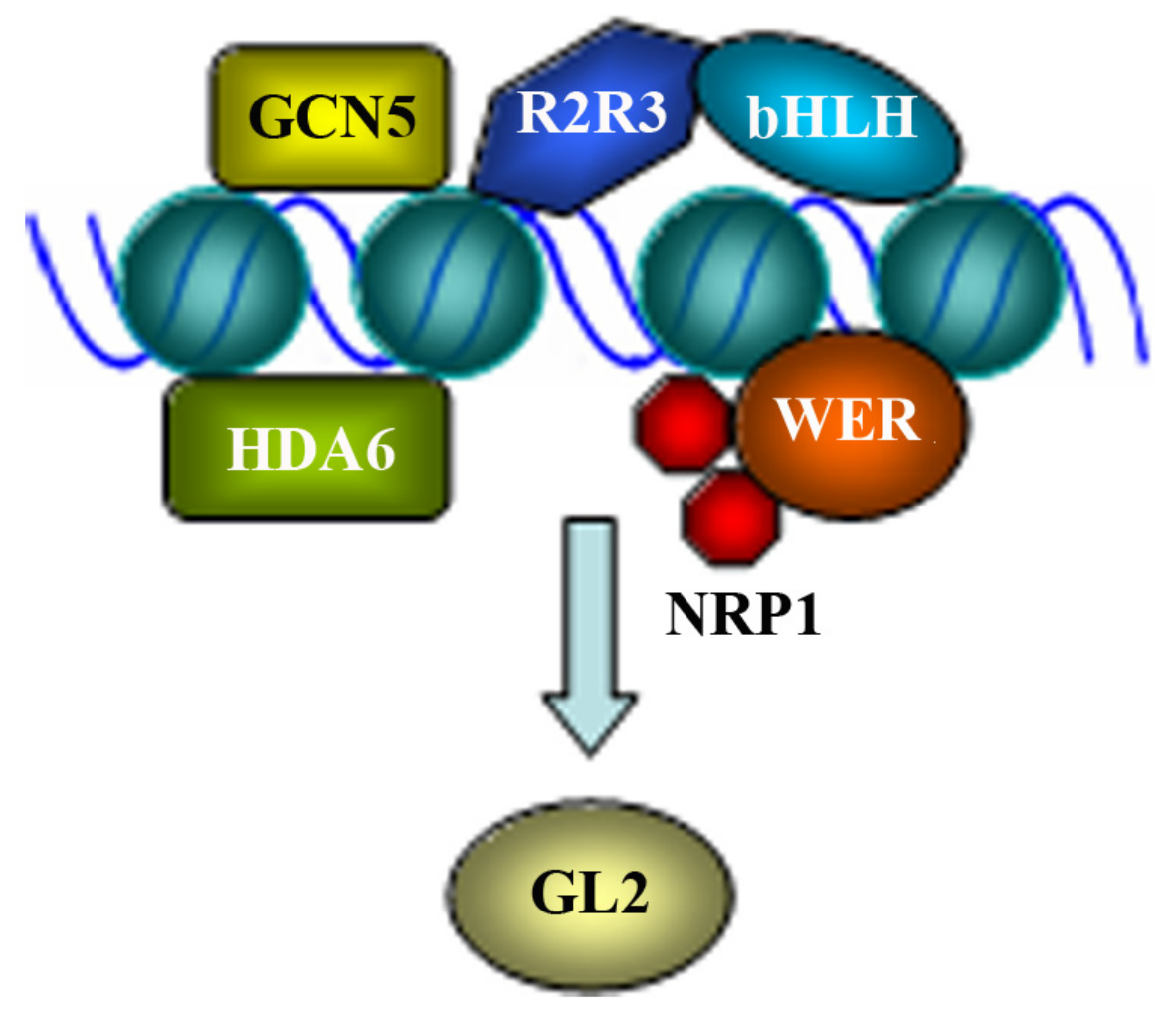
Epigenetic modification is involved in the regulation of GL2 expression (Figure 2). NAP1 (NUCLEOSOME ASSEMBLY PROTEIN1), a conserved nucleosome assembly protein in plants and animals acts as a histone chaperone. NAP1-RELATED PROTEIN1 (NRP1) and NRP2, two NAP1 homologues in Arabidopsis are involved in the regulation of root hair formation, as increased numbers of root hairs was observed in the nrp1-1 nrp2-1 double mutant [48]. Expression analysis of genes involved in the regulation of root hair formation showed a decreased expression level of GL2, suggesting that NRP1 and NRP2 are involved in the regulation of GL2 [48].
Indeed, it was soon confirmed that NRP1 is directly involved in the regulation of GL2 [49]. By using ChIP-PCR analysis, Zhu et al. [49] found that NRP1 was enriched at the promoter region of GL2 in wild type plants, but not the wer mutants, indicating that GL2 is a direct target gene of NRP1, and the enrichment of NRP1 at the promoter region of GL2 is dependent on WER. Protein–protein interaction assays in both vitro and in vivo showed that NRP1 was able to form a dimer via its N-terminal α-helix, and interacted with WER [49]. By comparing histone status and nucleosome density at GL2 promoter in the wild type and nrp1-1 nrp2-1 mutant plants, the authors further showed that the NRP proteins were able to promote histone release and decrease nucleosome density, thereby enables WER to form a stable complex with its target DNA [49].
Ectopic hair cells in the nonhair positions of root epidermis were also observed in hda6, a loss-of-function mutant of HISTONE DEACETYLASE6 (HDA6) [50]. On the other hand, increased leaf trichome density was observed in gcn5, a loss-of-function mutant of histone acetyltransferase gene GENERAL CONTROL NON-REPRESSED PROTEIN5 (GCN5) [51]. Expression level of GL2 was increased in the hda6 mutants, but decreased in the gcn5 mutants [50,51]. Consistent with this, both HDA6 and GCN5 have been found to bind directly to the promoter regions of GL2, and acetylation of histone H3 on the promoter regions of GL2 was increased in the hda6 mutant, but decrease in the gcn5 mutants [50,51]. These evidence suggest that HDA6 and GCN5 directly regulate the expression of GL2 by affecting the histone acetylation levels of the promoter regions of GL2, provided evidence that epigenetic modification of chromosome is involved in the regulation of GL2 expression.
3. Roles of GL2 in Regulating Epidermal Cell Fate Determination and Anthocyanin Biosynthesis
3.1. Regulation of Trichome Initiation and Root Hair Formation
GL2 was first identified as a regulator for trichome outgrowth, but not for trichome initiation [23]. This was further supported by the observation that the expression pattern of the PGL2:GUS reporter gene in the gl2 mutant was largely similar to that in the wild-type plants [39]. Later on, it was found that the gl2 mutant produces more root hairs, indicating that GL2 is required for the regulation of root hair formation [24]. Based on the characterization of other trichome and/or root hair related mutants, including the WD40-repeat gene mutant ttg1 [1,2], the R2R3 gene mutants gl1 and wer [3,4,7,8], the bHLH gene mutants gl3 and egl3 [9,10], and the R3 MYB gene mutants try, cpc, etc1, etc2, etc3, and tcl1 [52,53,54,55,56,57,58,59], and the analyses of protein–protein interaction in yeast cells [10,37], it was then proposed that the expression of both GL2 and the R3 MYB genes can be activated by MBW activator complexes, and work downstream of the MBW activator complexes to regulate trichome initiation and root hair formation [11,12,13,25].
Even though available evidence support that both GL2 and R3 MYBs function downstream of a MBW activator complex to regulate trichome initiation and root hair formation [11,13,25,26,32,33], some other studies suggest that the roles of GL2 in the transcription factor regulatory network that regulating trichome initiation and root hair formation is far more complicated than previously thought.
First, more feedback loops have been identified. Khosla et al [60] found that GL2 was able to activate the expression of MYB23, a functional homolog of GL1 [5], thus activating a positive feedback loop to regulate trichome initiation. MYB23 has also been shown to be a functional homolog of WER, and to be able to induce its own expression, thus also provides a positive feedback loop in regulating root hair formation [61]. Wang et al. [59] found that, in addition to compete with GL1 for binding GL3 or EGL3, blocking the formation of the complexes required for the activation of GL2, TCL1 was able to suppress the expression of GL1 by binding directly to its promoter region, added an additional negative feedback loop for the regulation of trichome initiation.
Second, the regulation of R3 MYB genes, antagonistic genes of GL2 in the regulation of trichome initiation and root hair formation, is more complicated than previously thought too. For example, it has been reported that the MYB complexes can only regulate the expression of some of the R3 MYB genes including TRY, CPC, ETC1 and ETC3, but not ETC2, TCL1 and TCL2 [43], and both the miR156 directed SQUAMOSA PROMOTER BINDING PROTEIN LIKE 9 (SPL9) and the membrane-associated NAM, ATAF1/2, and CUC (NAC) transcription factor NTM1-LIKE8 (NTL8) directly regulate the expression of TCL1 and TRY [62,63]. However, SPL9 and NTL8 did interact with each other in both yeast and plant cells, indicating that they function in different pathways to regulate the expression of TCL1 and TRY [63]. Considering that SPL9 alone can not activate the reporter gene in protoplast, it is very likely that SPL9 may interact with other proteins to regulate the expression of TCL1 and TRY [63]. In addition, Pesch et al. [64] found that TTG1 and GL1 were able to compete for binding GL3, thus differently regulating the expression of R3 MYB genes TRY and CPC. However, co-transfection of TTG1 and GL3 failed to activate the expression of GL2 in plant cells [43]. More recently, Li et al. [45] reported that TTG1 is able to interact with TT2 to regulate the expression of GL2. Considering that these studies mainly relied on fusion proteins and assays in yeast or protoplast cells [43,45,64], some of the results may need to be further examined.
Third, analysis of double and higher order mutants between gl2 and single-repeat R3 MYB gene mutants suggest that GL2 may play an essential role in regulating root hair formation but not trichome formation, i.e., GL2 is required for the inhibition of root hair formation, but may not be absolutely required for the promotion of trichome initiation in Arabidopsis [65], as mutation of R3 MYB genes in the gl2 mutant background produced trichomes, but did not affect the root hair phenotype of gl2 mutant [65]. It should be noted that these results were consistent with initial observation that GL2 functions as a regulator for trichome outgrowth, but not for trichome initiation [23].
The requirement for GL2 for the inhibition of root hair formation was further confirmed by Lin et al. [66]. They found that GL2 was able to directly suppress the expression DEFECTIVE6 (RHD6), RHD6-LIKE1 (RSL1), RSL2, LjRHL1-LIKE1 (LRL1) and LRL2, a few bHLH transcription factor genes that have previously been reported to be required for root hair formation, as root hair formation was inhibited in loss-of-function mutants of these genes [67,68,69,70,71]. In addition, Wu and Citovsky found that the plant specific proteins GL2-interacting repressor 1 (GIR1) and GIR2 interacted with GL2, and are involved in the regulation of root hair formation [72], suggest that GL2 may also function in complex with other regulators in regulating root hair formation in Arabidopsis.
3.2. Regulation of Anthocyanin Biosynthesis
As mentioned above, anthocyanin biosynthesis in Arabidopsis is another process that is regulated by similar MBW transcription activator complexes. However, instead of activating GL2, these complexes regulate anthocyanin biosynthesis by activating the expression of late biosynthesis genes in anthocyanin biosynthesis pathway, including F3’H, DFR, ANS/LDOX and UFGT [14,15,16,17,18,19,20]. By analyzing gl2-1D, an activation-tagged mutant, Wang et al. [26] found that GL2 negatively regulated anthocyanin biosynthesis in Arabidopsis. They found that anthocyanin accumulation is reduced in the gain-of-function mutant gl2-1D, and elevated in the loss-of-function mutant gl2-3. Quantitative RT-PCR results showed that expression of late biosynthesis genes, rather than early biosynthesis genes in anthocyanin biosynthesis pathway was decreased in the gl2-1D mutant, and increased in the gl2-3 mutant.
Because the expression of the late biosynthesis genes is regulated by the MBW transcriptional activator complexes [16,17,19,73], the authors thus further examined the expression of the MBW component genes in the mutants. They found that the expression of TT8, PAP1, PAP2, MYB113 and MYB114 was decreased in the gl2-1D mutant, and increased in the gl2-3 mutant [26]. Whereas chromatin immunoprecipitation assays indicated that at least some of these MBW activator complex component genes are direct targets of GL2 [26]. Consistent with these results, protoplast transient transfection assays showed that when recruited to the promoter region of the LexA-Gal4:GUS reporter gene by a fused Gal4 DNA binding domain (GD), GL2 repressed the expression of the reporter gene activated by the transcriptional activator, LD-VP, suggesting that GL2 functions as transcription repressor [26]. The identification of GL2 as a transcription repressor is also consistent with the observation that GL2 was able to directly suppress the expression RHD6, RSL1, RSL2, LRL1 and LRL2 [66].
Single-repeat R3 MYB transcription factors have been found to regulate anthocyanin biosynthesis by blocking the formation of late biosynthesis genes regulating MBW activator complexes [74,75]. The finding that GL2 negatively regulates anthocyanin biosynthesis in Arabidopsis by directly repressing the expression of some of the MBW activator complexes genes adds another feed back loop to the control of anthocyanin biosynthesis. This finding also provides further connection between the regulatory network regulating epidermal cell fate determination including trichome initiation and root hair formation, and the one regulating anthocyanin biosynthesis in Arabidopsis.
4. Challenges and Future Perspectives
More and more evidence suggest that GL2 plays a central role in the regulation of epidermal cell fate determination and anthocyanin biosynthesis. In addition, GL2 is also involved in the regulation of other processes such as hypocotyl stomatal development, seed coat development, seed oil production, and seed coat mucilage biosynthesis [12,22,35,76,77,78]. However, to uncover the functional mechanisms of GL2 in a different process is still a challenge.
Identification of direct target genes may help to reveal the functional mechanisms of GL2. As a transcription factor, it is reasonable to assume that GL2 regulates these processes by regulating its target gene expression. However, so far, only a few genes have been identified as direct target genes of GL2 (Table 1). Based on large-scale gene expression analyses, a large number of genes were found to be regulated downstream of GL2 during epidermal cell fate determination [70,79,80,81]. A combination of transcriptome analysis and ChIP-qPCR may be used to examine if any of these genes are direct targets of GL2. Transcriptome analysis by using DEX-inducible GL2 transgenic plants may narrow down candidates of GL2’s direct target genes, and ChIP-qPCR assays may be used to examine if any of the candidate genes are direct targets of GL2. On the other hand, ChIP sequencing will enable genome wide identification of transcription factor binding sites, performing a ChIP sequencing using GL2 transgenic plants will be a good start for the identification of additional target genes of GL2.
Examining how GL2 regulates its target genes will also help to reveal the functional mechanisms of GL2. Consistent with the results that GL2 functions as a transcription repressor in protoplasts transient transfection assays [26], GL2 negatively regulates the expression of several target genes including CELLULOSE SYNTHASE5 (CESA5), PLDζ1, TT8, PAP2 and MYB113 [82,83]. However, the expression of XYLOGLUCAN ENDOTRANSGLUCOSYLASE17 (XTH17) and MYB23 was reported to be positively regulated by GL2, and both of them have also been identified as direct target genes of GL2 [60,83]. It may be hard to explain how a transcription repressor may activate gene expression. One speculation is that GL2 may interact with other regulators to regulate its target genes. Indeed, it has been shown that activation of MYB23 requires both GL2 and MYB23 [60], and GIR1 and GIR2 have been found to interact with GL2 to regulate root hair formation [72]. However, considering that GIR1 contains EAR repression motif and functioned as transcription repressor [78], it is unlikely that GL2-GIR1/GIR2 may function as transcription activator complex. Therefore, it will be of interesting to identify and characterize other GL2 interacting proteins if any, and to examine if GIR1 and GIR2 are also involved in the regulation of other growth and development processes regulated by GL2. Considering that GIR1 and GIR2 are also interacting with the major transcription co-repressor TOPLESS (TPL) [84], it may also be of great interest to examine whether GL2, GIR1/GIR2 and TPL may form a repressor complex to regulate the expression of their target genes.
MYBL2, a R3-MYB-related protein, also functions as a transcription repressor [85], and a negative regulator of anthocyanin biosynthesis in Arabidopsis [85,86]. In addition to interacting with bHLH transcription factors to inhibit the activity of the MBW activator complexes [85], MYBL2 is able to directly regulate the expression of TT8 [85], one of the direct target genes of GL2. Therefore, it is worthwhile to examine the relationship between GL2 and MYBL2 in the regulation of anthocyanin biosynthesis in Arabidopsis.
It should be note that ANTHOCYANINLESS2 (ANL2), an HD-GLABRA2 group HD-ZIP transcription factor, has been shown to regulate anthocyanin biosynthesis in Arabidopsis [87]. In contradiction to the negative role of GL2, ANL2 positively regulates anthocyanin biosynthesis [86]. Examining if ANL2 may regulate the expression of the MBW activator complex component genes, and/or if it may be involved in the regulation of cell fate determination, may help to reveal the relationship between ANL2 and GL2.
Accumulated evidence suggests that anthocyanin biosynthesis in eudicots is controlled by a conserved transcription factor network [17,88]. It will be of great interest to examine if GL2 homologues in other plants may be involved in the regulation of anthocyanin biosynthesis, if so, their role in the regulation of anthocyanin biosynthesis.
Funding
This research was supported by the National Key R&D Program of China, grant number 2016YFD0101900.
Acknowledgments
The author wishes to thank all the colleagues for their work in this research field, and apologize to those whose work could not be mentioned because of the specific focus of this review. This work was supported by the National Key R&D Program of China (2016YFD0101900). The funder had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflicts of Interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Galway, M.E.; Masucci, J.D.; Lloyd, A.M.; Walbot, V.; Davis, R.W.; Schiefelbein, J.W. The TTG gene is required to specify epidermal cell fate and cell patterning in the Arabidopsis root. Dev. Biol. 1994, 166, 740–754. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R.; Davison, P.A.; Bolognesi-Winfield, A.C.; James, C.M.; Srinivasan, N.; Blundell, T.L.; Esch, J.J.; Marks, M.D.; Gray, J.C. The TRANSPARENT TESTA GLABRA1 locus, which regulates trichome differentiation and anthocyanin biosynthesis in Arabidopsis, encodes a WD40 repeat protein. Plant Cell 1999, 11, 1337–1350. [Google Scholar] [CrossRef] [PubMed]
- Oppenheimer, D.G.; Herman, P.L.; Sivakumaran, S.; Esch, J.; Marks, M.D. A myb gene required for leaf trichome differentiation in Arabidopsis is expressed in stipules. Cell 1991, 67, 483–493. [Google Scholar] [CrossRef]
- Lee, M.M.; Schiefelbein, J. WEREWOLF, a MYB-related protein in Arabidopsis, is a position-dependent regulator of epidermal cell patterning. Cell 1999, 99, 473–483. [Google Scholar] [CrossRef]
- Kirik, V.; Lee, M.M.; Wester, K.; Hermann, U.; Zheng, Z.; Oppenheimer, D.; Schiefelbein, J.; Hulskamp, M. Functional diversificatin of MYB23 and GL1 genes in trichome morphogenesis and initiation. Development 2005, 132, 1477–1485. [Google Scholar] [CrossRef] [PubMed]
- Symonds, W.; Hatlestad, G.; Lloyd, A.M. Natural allelic variation defines a role for ATMYC1: Trichome cell fate determination. PLoS Genet. 2011, 7, e1002069. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, C.; Lee, M.M.; Gonzalez, A.; Zhang, F.; Lloyd, A.; Schiefelbein, J. The bHLH genes GLABRA3 (GL3) and ENHANCER OF GLABRA3 (EGL3) specify epidermal cell fate in the Arabidopsis root. Development 2003, 130, 6431–6439. [Google Scholar] [CrossRef]
- Bernhardt, C.; Zhao, M.; Gonzalez, A.; Lloyd, A.; Schiefelbein, J. The bHLH genes GL3 and EGL3 participate in an intercellular regulatory circuit that controls cell patterning in the Arabidopsis root epidermis. Development 2005, 132, 291–298. [Google Scholar] [CrossRef]
- Payne, C.T.; Zhang, F.; Lloyd, A.M. GL3 encodes a bHLH protein that regulates trichome development in Arabidopsis through interaction with GL1 and TTG1. Genetics 2000, 156, 1349–1362. [Google Scholar]
- Zhang, F.; Gonzalez, A.; Zhao, M.; Payne, C.T.; Lloyd, A. A network of redundant bHLH proteins functions in all TTG1-dependent pathways of Arabidopsis. Development 2003, 130, 4859–4869. [Google Scholar] [CrossRef]
- Schiefelbein, J. Cell-fate specification in the epidermis: A common patterning mechanism in the root and shoot. Curr. Opin. Plant Biol. 2003, 6, 74–78. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Aoyama, T. Pathways for epidermal cell differentiation via the homeobox gene GLABRA2: Update on the roles of the classic regulator. J. Integr. Plant Biol. 2012, 54, 729–737. [Google Scholar] [PubMed]
- Wang, S.; Chen, J.G. Regulation of cell fate determination by single-repeat R3 MYB transcription factors in Arabidopsis. Front. Plant Sci. 2014, 5, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baudry, A.; Heim, M.A.; Dubreucq, B.; Caboche, M.; Weisshaar, B.; Lepiniec, L. TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. Plant J. 2004, 39, 366–380. [Google Scholar] [CrossRef] [PubMed]
- Nesi, N.; Debeaujon, I.; Jond, C.; Pelletier, G.; Caboche, M.; Lepiniec, L. The TT8 gene encodes a basic helix-loop-helix domain protein required for expression of DFR and BAN genes in Arabidopsis siliques. Plant Cell 2000, 12, 1863–1878. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Petroni, K.; Tonelli, C. Recent advances on the regulation of anthocyanin synthesis in reproductive Organs. Plant Sci. 2011, 181, 219–229. [Google Scholar] [CrossRef]
- Patra, B.; Schluttenhofer, C.; Wu, Y.; Pattanaik, S.; Yuan, L. Transcriptional regulation of secondary metabolite biosynthesis in plants. Biochim. Biophys. Acta 2013, 1829, 1236–1247. [Google Scholar] [CrossRef]
- Xu, W.; Grain, D.; Bobet, S.; Le Gourrierec, J.; Thévenin, J.; Kelemen, Z.; Lepiniec, L.; Dubos, C. Complexity and robustness of the flavonoid transcriptional regulatory network revealed by comprehensive analyses of MYB-bHLH-WDR complexes and their targets in Arabidopsis seed. New Phytol. 2014, 202, 132–144. [Google Scholar] [CrossRef]
- Xu, W.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB–bHLH–WDR complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef]
- Li, S.F.; Milliken, O.N.; Pham, H.; Seyit, R.; Napoli, R.; Preston, J.; Koltunow, A.M.; Parish, R.W. The Arabidopsis MYB5 transcription factor regulates mucilage synthesis, seed coat development, and trichome morphogenesis. Plant Cell 2009, 21, 72–89. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Mendenhall, J.; Huo, Y.; Lloyd, A. TTG1 complex MYBs, MYB5 and TT2, control outer seed coat differentiation. Dev. Biol. 2009, 325, 412–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rerie, W.G.; Feldmann, K.A.; Marks, M.D. The GLABRA2 gene encodes a homeo domain protein required for normal trichome development in Arabidopsis. Genes Dev. 1994, 8, 1388–1399. [Google Scholar] [CrossRef] [PubMed]
- Di Cristina, M.; Sessa, G.; Dolan, L.; Linstead, P.; Baima, S.; Ruberti, I.; Morelli, G. The Arabidopsis Athb-10 (GLABRA2) is an HD-Zip protein required for regulation of root hair development. Plant J. 1996, 10, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Schiefelbein, J.; Huang, L.; Zheng, X. Regulation of epidermal cell fate in Arabidopsis roots: The importance of multiple feed back loops. Front. Plant Sci. 2014, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, X.; Hu, Q.; Dai, X.; Tian, H.; Zheng, K.; Wang, X.; Mao, T.; Chen, J.G.; Wang, S. Characterization of an activation-tagged mutant uncovers a role of GLABRA2 in anthocyanin biosynthesis in Arabidopsis. Plant J. 2015, 83, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Elhiti, M.; Stasolla, C. Structure and function of homodomain-leucine zipper (HD-Zip) proteins. Plant Signal Behav. 2009, 4, 86–88. [Google Scholar] [CrossRef] [Green Version]
- Herman, P.L.; Marks, M.D. Trichome development in Arabidopsis thaliana. II. Isolation and complementation of the GLABROUS1 Gene. Plant Cell 1989, 1, 1051–1055. [Google Scholar] [CrossRef]
- Esch, J.J.; Chen, M.A.; Sanders, M.; Hillestad, M.; Ndkium, S.; Idelkope, B.; Neizer, J.; Marks, M.D. A contradictory GLABRA3 allele helps define gene interactions controlling trichome development in Arabidopsis. Development 2003, 130, 5885–5894. [Google Scholar] [CrossRef]
- Pesch, M.; Hülskamp, M. Creating a two-dimensional pattern de novo during Arabidopsis trichome and root hair initiation. Curr. Opin. Genet. Dev. 2004, 14, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Ishida, T.; Kurata, T.; Okada, K.; Wada, T. A Genetic Regulatory Network in the Development of Trichomes and Root Hairs. Annu. Rev. Plant Biol. 2008, 59, 364–386. [Google Scholar] [CrossRef] [PubMed]
- Pesch, M.; Hülskamp, M. One, two, three…models for trichome patterning in Arabidopsis? Curr. Opin. Plant Biol. 2009, 12, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Hülskamp, M.; Misra, S.; Jürgens, G. Genetic dissection of trichome cell development in Arabidopsis. Cell 1994, 76, 555–566. [Google Scholar] [CrossRef]
- Hung, C.Y.; Lin, Y.; Zhang, M.; Pollock, S.; Marks, M.D.; Schiefelbein, J. A common position-dependent mechanism controls cell type patterning and GLABRA2 regulation in the root and hypocotyl epidermis of Arabidopsis. Plant Physiol. 1998, 117, 73–84. [Google Scholar] [CrossRef]
- Schellmann, S.; Schnittger, A.; Kirik, V.; Wada, T.; Okada, K.; Beermann, A.; Thumfahrt, J.; Jürgens, G.; Hülskamp, M. TRIPTYCHON and CAPRICE mediate lateral inhibition during trichome and root hair patterning in Arabidopsis. EMBO J. 2002, 21, 5036–5046. [Google Scholar] [CrossRef]
- Zimmermann, I.M.; Heim, M.A.; Weisshaar, B.; Uhrig, J.F. Comprehensive identification of Arabidopsis thaliana MYB transcription factors interacting with R/B-like BHLH proteins. Plant J. 2004, 40, 22–34. [Google Scholar] [CrossRef]
- Zhao, M.; Morohashi., K.; Hatlestad., G.; Grotewold., E.; Lloyd, A. The TTG1-bHLH-MYB complex controls trichome cell fate and patterning through direct targeting of regulatory loci. Development 2008, 135, 1991–1999. [Google Scholar] [CrossRef] [Green Version]
- Szymanski, D.B.; Jilk, R.A.; Pollock, S.M.; Marks, M.D. Control of GL2 expression in Arabidopsis leaves and trichomes. Development 1998, 125, 1161–1171. [Google Scholar]
- Morohashi, K.; Zhao, M.; Yang, M.; Read, B.; Lloyd, A.; Lamb, R.; Grotewold, E. Participation of the Arabidopsis bHLH factor GL3 in trichome initiation regulatory events. Plant Physiol. 2007, 145, 736–746. [Google Scholar] [CrossRef]
- Wang, S.; Chen, J.G. Arabidopsis transient expression analysis reveals that activation of GLABRA2 may require concurrent bindings of GLABRA1 and GLABRA3 to the Promoter of GLABRA2. Plant Cell Physiol. 2008, 49, 1792–1804. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhou, L.; Zhang, W.; Cai, L.; Guo, H.; Tian, H.; Schiefelbein, J.; Wang, S. A single amino acid substitution in the R3 domain of GLABRA1 leads to inhibition of trichome initiation in Arabidopsis without affecting its interaction with GLABRA3. Plant Cell Environ. 2016, 39, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hubbard, L.; Chang, Y.; Guo, J.; Schiefelbein, J.; Chen, J.G. Comprehensive analysis of single-repeat R3 MYB proteins in epidermal cell patterning and their transcriptional regulation in Arabidopsis. BMC Plant Biol. 2008, 8, 81. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Xia, K.; Chen, J.G.; Wang, S. Functional characterization of TRICHOMELESS2, a new single repeat R3 MYB transcription factor in the regulation of trichome patterning in Arabidopsis. BMC Plant Biol. 2011, 11, 176. [Google Scholar] [CrossRef]
- Li, C.; Zhang, B.; Chen, B.; Ji, L.; Yu, H. Site-specific phosphorylation of TRANSPARENT TESTA GLABRA1 mediates carbon partitioning in Arabidopsis seeds. Nat Commun. 2018, 9, 571. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Zheng, K.; Wang, X.; Tian, H.; Wang, X.; Wang, S. Control of trichome initiation in Arabidopsis by poplar single-repeat R3 MYB transcription factors. Front. Plant Sci. 2014, 5, 262. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Tian, H.; Hu, Q.; Guo, H.; Yang, L.; Cai, L.; Wang, X.; Liu, B.; Wang, S. Ectopic expression of R3 MYB transcription factor gene OsTCL1 in Arabidopsis, but not rice, affects trichome and root hair formation. Sci. Rep. 2016, 6, 19254. [Google Scholar] [CrossRef]
- Zhu, Y.; Dong, A.; Meyer, D.; Pichon, O.; Renou, J.P.; Cao, K.; Shen, W.H. Arabidopsis NRP1 and NRP2 encode histone chaperones and are required for maintaining postembryonic root growth. Plant Cell 2006, 18, 2879–2892. [Google Scholar] [CrossRef]
- Zhu, Y.; Rong, L.; Luo, Q.; Wang, B.; Zhou, N.; Yang, Y.; Zhang, C.; Feng, H.; Zheng, L.; Shen, W.H.; et al. The Histone Chaperone NRP1 Interacts with WEREWOLF to Activate GLABRA2 in Arabidopsis Root Hair Development. Plant Cell 2017, 29, 260–276. [Google Scholar] [CrossRef]
- Li, D.X.; Chen, W.Q.; Xu, Z.H.; Bai, S.N. HISTONE DEACETYLASE6-defective mutants show increased expression and acetylation of ENHANCER OF TRIPTYCHON AND CAPRICE1 and GLABRA2 with small but significant effects on root epidermis cellular pattern. Plant Physiol. 2015, 168, 1448–1458. [Google Scholar] [CrossRef]
- Wang, T.; Jia, Q.; Wang, W.; Hussain, S.; Ahmed, S.; Adnan; Zhou, D.X.; Ni, Z.; Wang, S. GCN5 modulates trichome initiation in Arabidopsis by manipulating histone acetylation of core trichome initiation regulator genes. Plant Cell Rep. 2019, 38, 755–765. [Google Scholar] [CrossRef]
- Wada, T.; Tachibana, T.; Shimura, Y.; Okada, K. Epidermal cell differentiation in Arabidopsis determined by a Myb homolog, CPC. Science 1997, 277, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Schnittger, A.; Folkers, U.; Schwab, B.; Jürgens, G.; Hülskamp, M. Generation of a spacing pattern: The role of TRIPTYCHON in trichome patterning in Arabidopsis. Plant Cell 1999, 11, 1105–1116. [Google Scholar] [CrossRef]
- Esch, J.J.; Chen, M.A.; Hillestad, M.; Marks, M.D. Comparison of TRY and the closely related At1g01380 gene in controlling Arabidopsis trichome patterning. Plant J. 2004, 40, 860–869. [Google Scholar] [CrossRef]
- Kirik, V.; Simon, M.; Hülskamp, M.; Schiefelbein, J. The ENHANCER OF TRY AND CPC1 gene acts redundantly with TRIPTYCHON and CAPRICE in trichome and root hair cell patterning in Arabidopsis. Dev. Biol. 2004, 268, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Kirik, V.; Simon, M.; Wester, K.; Schiefelbein, J.; Hülskamp, M. ENHANCER of TRY and CPC 2(ETC2) reveals redundancy in the region-specific control of trichome development of Arabidopsis. Plant Mol. Biol. 2004, 55, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; Lee, M.M.; Lin, Y.; Gish, L.; Schiefelbein, J. Distinct and overlapping roles of single-repeat MYB genes in root epidermal patterning. Dev. Biol. 2007, 311, 566–578. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, R.; Iwata, M.; Sano, R.; Inoue, K.; Okada, K.; Wada, T. Arabidopsis CAPRICE-LIKE MYB 3 (CPL3) controls endoreduplication and flowering development in addition to trichome and root hair formation. Development 2008, 135, 1335–1345. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kwak, S.H.; Zeng, Q.; Ellis, B.E.; Chen, X.Y.; Schiefelbein, J.; Chen, J.G. TRICHOMELESS1 regulates trichome patterning by suppressing GLABRA1 in Arabidopsis. Development 2007, 134, 3873–3882. [Google Scholar] [CrossRef]
- Khosla, A.; Paper, J.M.; Boehler, A.P.; Bradley, A.M.; Neumann, T.R.; Schrick, K. HD-Zip proteins GL2 and HDG11 have redundant functions in Arabidopsis trichomes, and GL2 activates a positive feedback loop via MYB23. Plant Cell 2014, 26, 2184–2200. [Google Scholar] [CrossRef]
- Kang, Y.H.; Kirik, V.; Hulskamp, M.; Nam, K.H.; Hagely, K.; Lee, M.M.; Schiefelbein, J. The MYB23 gene provides a positive feedback loop for cell fate specification in the Arabidopsis root epidermis. Plant Cell 2009, 21, 1080–1094. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Cai, W.J.; Wang, S.; Shan, C.M.; Wang, L.J.; Chen, X.Y. Temporal control of trichome distribution by microRNA156-targeted SPL genes in Arabidopsis thaliana. Plant Cell 2010, 22, 2322–2335. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Wang, X.; Guo, H.; Cheng, Y.; Hou, C.; Chen, J.G.; Wang, S. NTL8 regulates trichome formation in Arabidopsis by directly activating R3 MYB genes TRY and TCL1. Plant Physiol. 2017, 174, 2363–2375. [Google Scholar] [CrossRef] [PubMed]
- Pesch, M.; Schultheiß, I.; Klopffleisch, K.; Uhrig, J.F.; Koegl, M.; Clemen, C.S.; Simon, R.; Weidtkamp-Peters, S.; Hülskamp, M. TRANSPARENT TESTA GLABRA1 and GLABRA1 Compete for Binding to GLABRA3 in Arabidopsis. Plant Physiol. 2015, 168, 584–597. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Barron, C.; Schiefelbein, J.; Chen, J.G. Distinct relationships between GLABRA2 and single-repeat R3 MYB transcription factors in the regulation of trichome and root hair patterning in Arabidopsis. New Phytol. 2010, 185, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Ohashi, Y.; Kato, M.; Tsuge, T.; Gu, H.; Qu, L.J.; Aoyama, T. GLABRA2 Directly Suppresses Basic Helix-Loop-Helix Transcription Factor Genes with Diverse Functions in Root Hair Development. Plant Cell 2015, 27, 2894–2906. [Google Scholar]
- Masucci, J.D.; Schiefelbein, J.W. The rhd6 mutation of Arabidopsis thaliana alters root-hair initiation through an auxin- and ethylene-associated process. Plant Physiol. 1994, 106, 1335–1346. [Google Scholar] [CrossRef]
- Menand, B.; Yi, K.; Jouannic, S.; Hoffmann, L.; Ryan, E.; Linstead, P.; Schaefer, D.G.; Dolan, L. An ancient mechanism controls the development of cells with a rooting function in land plants. Science 2007, 316, 1477–1480. [Google Scholar] [CrossRef]
- Karas, B.; Amyot, L.; Johansen, C.; Sato, S.; Tabata, S.; Kawaguchi, M.; Szczyglowski, K. Conservation of lotus and Arabidopsis basic helix-loop-helix proteins reveals new players in root hair development. Plant Physiol. 2009, 151, 1175–1185. [Google Scholar] [CrossRef]
- Yi, K.; Menand, B.; Bell, E.; Dolan, L. A basic helix-loop-helix transcription factor controls cell growth and size in root hairs. Nat. Genet. 2010, 42, 64–67. [Google Scholar] [CrossRef]
- Bruex, A.; Kainkaryam, R.M.; Wieckowski, Y.; Kang, Y.H.; Bernhardt, C.; Xia, Y.; Zheng, X.; Wang, J.Y.; Lee, M.M.; Benfey, P.; et al. A gene regulatory network for root epidermis cell differentiation in Arabidopsis. PLoS Genet. 2012, 8, e1002446. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Citovsky, V. Adaptor proteins GIR1 and GIR2. I. Interaction with the repressor GLABRA2 and regulation of root hair development. Biochem. Biophys. Res. Commun. 2017, 488, 547–553. [Google Scholar] [CrossRef]
- Li, S. Transcriptional control of flavonoid biosynthesis: Fine-tuning of the MYB-bHLH-WD40 (MBW) complex. Plant Signal. Behav. 2014, 9, e27522. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.F.; Fitzsimmons, K.; Khandelwal, A.; Kranz, R.G. CPC, a single-repeat R3 MYB, is a negative regulator of anthocyanin biosynthesis in Arabidopsis. Mol. Plant 2009, 2, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Nemie-Feyissa, D.; Olafsdottir, S.M.; Heidari, B.; Lillo, C. Nitrogen depletion and small R3-MYB transcription factors affecting antho cyanin accumulation in Arabidopsis leaves. Phytochemistry 2014, 98, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Western, T.L.; Young, D.S.; Dean, G.H.; Tan, W.L.; Samuels, A.L.; Haughn, G.W. MUCILAGE-MODIFIED4 encodes a putative pectin biosynthetic enzyme developmentally regulated by APETALA2, TRANSPARENT TESTA GLABRA1, and GLABRA2 in the Arabidopsis seed coat. Plant Physiol. 2004, 134, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Sinkevicius, K.W.; Selinger, D.A. Tarczynski MC The homeobox gene GLABRA2 affects seed oil content in Arabidopsis. Plant Mol. Biol. 2006, 60, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Katavic, V.; Yu, Y.; Kunst, L.; Haughn, G. Arabidopsis glabra2 mutant seeds deficient in mucilage biosynthesis produce more oil. Plant J. 2012, 69, 37–46. [Google Scholar] [CrossRef]
- Lieckfeldt, E.; Simon-Rosin, U.; Kose, F.; Zoeller, D.; Schliep, M.; Fisahn, J. Gene expression profiling of single epidermal, basal and trichome cells of Arabidopsis thaliana. J. Plant Physiol. 2007, 165, 1227–1237. [Google Scholar] [CrossRef]
- Marks, M.D.; Betancur, L.; Gilding, E.; Chen, F.; Bauer, S.; Wenger, J.P.; Dixon, R.A.; Haigler, C.H. A new method for isolating large quantities of Arabidopsis trichomes for transcriptome, cell wall and other types of analyses. Plant J. 2008, 56, 483–492. [Google Scholar] [CrossRef]
- Won, S.K.; Lee, Y.J.; Lee, H.Y.; Hoe, Y.K.; Cho, M.; Cho, H.T. Ciselement- and transcriptome-based screening of root hair-specific genes and their functional characterization in Arabidopsis. Plant Physiol. 2009, 150, 1459–1473. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, Y.; Oka, A.; Rodrigues-Pousada, R.; Possenti, M.; Ruberti, I.; Morelli, G.; Aoyama, T. Modulation of phospholipid signalling by GLABRA2 in root-hair pattern formation. Science 2003, 300, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Tominaga-Wada, R.; Iwata, M.; Sugiyama, J.; Kotake, T.; Ishida, T.; Yokoyama, R.; Nishitani, K.; Okada, K.; Wada, T. The GLABRA2 homeodomain protein directly regulates CESA5 and XTH17 gene expression in Arabidopsis roots. Plant J. 2009, 60, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Citovsky, V. Adaptor proteins GIR1 and GIR2. II. Interaction with the co-repressor TOPLESS and promotion of histone deacetylation of target chromatin. Biochem. Biophys. Res. Commun. 2017, 488, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Umemura, Y.; Ohme-Takagi, M. AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant J. 2008, 55, 954–967. [Google Scholar] [CrossRef] [PubMed]
- Dubos, C.; Le Gourrierec, J.; Baudry, A.; Huep, G.; Lanet, E.; Debeaujon, I.; Routaboul, J.M.; Alboresi, A.; Weisshaar, B.; Lepiniec, L. MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J. 2008, 55, 940–953. [Google Scholar] [CrossRef]
- Kubo, H.; Peeters, A.J.; Aarts, M.G.; Pereira, A.; Koornneef, M. ANTHOCYANINLESS2, a homeobox gene affecting anthocyanin distribution and root development in Arabidopsis. Plant Cell 1999, 11, 1217–1226. [Google Scholar] [CrossRef]
- Albert, N.W.; Davies, K.M.; Lewis, D.H.; Zhang, H.; Montefiori, M.; Brendolise, C.; Boase, M.R.; Ngo, H.; Jameson, P.E.; Schwinn, K.E. A conserved network of transcriptional activators and repressors regulates anthocyanin pigmentation in eudicots. Plant Cell 2014, 26, 962–980. [Google Scholar] [CrossRef]
Figure 1.
Components of the epidermal cell fate determination, flavonoid and mucilage biosynthesis regulating MBW activator complexes. Some of the components are presented in all the MBW activator complexes, some are not. The WD40-repeat protein TTG1, and the bHLH transcription factors EGL3 are involved in the regulation of epidermal cell fate determination, flavonoid and mucilage biosynthesis. The bHLH transcription factor MYC1 is involved in the regulation of trichome initiation. TT8 is involved in the regulation of flavonoid and mucilage biosynthesis. The MYB transcription factors involved in the regulation of epidermal cell fate determination, flavonoid and mucilage biosynthesis are largely different. WER is involved in the regulation of root hair formation. GL1 is involved in the regulation of trichome initiation, MYB5 is involved in the regulation of mucilage biosynthesis, and PAP1, PAP2, MYB113 and MYB114 are involved in the regulation of flavonoid biosynthesis. However, MYB23 is involved in the regulation of trichome initiation and root hair formation, whereas TT2 is involved in the regulation of flavonoid and mucilage biosynthesis.
Figure 1.
Components of the epidermal cell fate determination, flavonoid and mucilage biosynthesis regulating MBW activator complexes. Some of the components are presented in all the MBW activator complexes, some are not. The WD40-repeat protein TTG1, and the bHLH transcription factors EGL3 are involved in the regulation of epidermal cell fate determination, flavonoid and mucilage biosynthesis. The bHLH transcription factor MYC1 is involved in the regulation of trichome initiation. TT8 is involved in the regulation of flavonoid and mucilage biosynthesis. The MYB transcription factors involved in the regulation of epidermal cell fate determination, flavonoid and mucilage biosynthesis are largely different. WER is involved in the regulation of root hair formation. GL1 is involved in the regulation of trichome initiation, MYB5 is involved in the regulation of mucilage biosynthesis, and PAP1, PAP2, MYB113 and MYB114 are involved in the regulation of flavonoid biosynthesis. However, MYB23 is involved in the regulation of trichome initiation and root hair formation, whereas TT2 is involved in the regulation of flavonoid and mucilage biosynthesis.

Figure 2.
Regulation of GL2 expression. Several different regulators are involved in the regulation of GL2, and the expression of GL2 may be regulated at different ways. (i) Interaction of R2R3 MYB protein GL1 or WER and bHLH protein GL3 or EGL3 are required and sufficient to activate GL2, and concurrent binding of the R2R3 MYB and bHLH proteins to the promoter region of GL2 is required for the activation. (ii) Dimer of histone chaperone NAP1-RELATED PROTEIN1 (NRP1) interacts with WER and bind to the promoter region of GL2 to regulate its expression. NRP2 is also involved in the regulation of GL2. (iii) Histone deacetylase HDA6 binds directly to the promoter region of GL2 to regulate its expression. (iv) Histone acetyltransferase GCN5 binds directly to the promoter region of GL2 to regulate its expression. It is unclear if HDA6 and GCN5 regulated expression of GL2 is depend on the R2R3 MYB proteins GL1 or WER and the bHLH proteins GL3 or EGL3, and if the regulators function in sequential to regulate the expression of GL2.
Figure 2.
Regulation of GL2 expression. Several different regulators are involved in the regulation of GL2, and the expression of GL2 may be regulated at different ways. (i) Interaction of R2R3 MYB protein GL1 or WER and bHLH protein GL3 or EGL3 are required and sufficient to activate GL2, and concurrent binding of the R2R3 MYB and bHLH proteins to the promoter region of GL2 is required for the activation. (ii) Dimer of histone chaperone NAP1-RELATED PROTEIN1 (NRP1) interacts with WER and bind to the promoter region of GL2 to regulate its expression. NRP2 is also involved in the regulation of GL2. (iii) Histone deacetylase HDA6 binds directly to the promoter region of GL2 to regulate its expression. (iv) Histone acetyltransferase GCN5 binds directly to the promoter region of GL2 to regulate its expression. It is unclear if HDA6 and GCN5 regulated expression of GL2 is depend on the R2R3 MYB proteins GL1 or WER and the bHLH proteins GL3 or EGL3, and if the regulators function in sequential to regulate the expression of GL2.


{kind=link}
{kind=link}
Table 1.
Confirmed target genes of GL2 and their functions in Arabidopsis.
| Targets | Functions | References |
|---|---|---|
| MYB23 | Trichome and root hair formation | Khosla et al., 2014 [60] |
| XTH17 | Secondary cell wall formation | Tominaga-Wada et al., 2009 [83] |
| CESA5 | Secondary cell wall formation | Tominaga-Wada et al., 2009 [83] |
| PLDζ1 | Root hair formation | Ohashi et al.,2003 [82] |
| MUM4 | Seed oil production | Shi et al., 2011 [78], Western et al., 2004 [76] |
| TT8 | Anthocyanin biosynthesis | Wang et al., 2015 [26] |
| PAP1 | Anthocyanin biosynthesis | Wang et al., 2015 [26] |
| PAP2 | Anthocyanin biosynthesis | Wang et al., 2015 [26] |
| MYB113 | Anthocyanin biosynthesis | Wang et al., 2015 [26] |
| MYB114 | Anthocyanin biosynthesis | Wang et al., 2015 [26] |
| RHD6 | Root hair formation | Lin et al., 2015 [66] |
| RSL1 | Root hair formation | Lin et al., 2015 [66] |
| RSL2 | Root hair formation | Lin et al., 2015 [66] |
| LRL1 | Root hair formation | Lin et al., 2015 [66] |
| LRL2 | Root hair formation | Lin et al., 2015 [66] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, S.; Wang, S. GLABRA2, a Common Regulator for Epidermal Cell Fate Determination and Anthocyanin Biosynthesis in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 4997. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20204997
AMA Style
Chen S, Wang S. GLABRA2, a Common Regulator for Epidermal Cell Fate Determination and Anthocyanin Biosynthesis in Arabidopsis. International Journal of Molecular Sciences. 2019; 20(20):4997. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20204997
Chicago/Turabian StyleChen, Siyu, and Shucai Wang. 2019. "GLABRA2, a Common Regulator for Epidermal Cell Fate Determination and Anthocyanin Biosynthesis in Arabidopsis" International Journal of Molecular Sciences 20, no. 20: 4997. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20204997
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.