Differences in Expression of Human Leukocyte Antigen Class II Subtypes and T Cell Subsets in Behçet’s Disease with Arthritis

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics of Patients
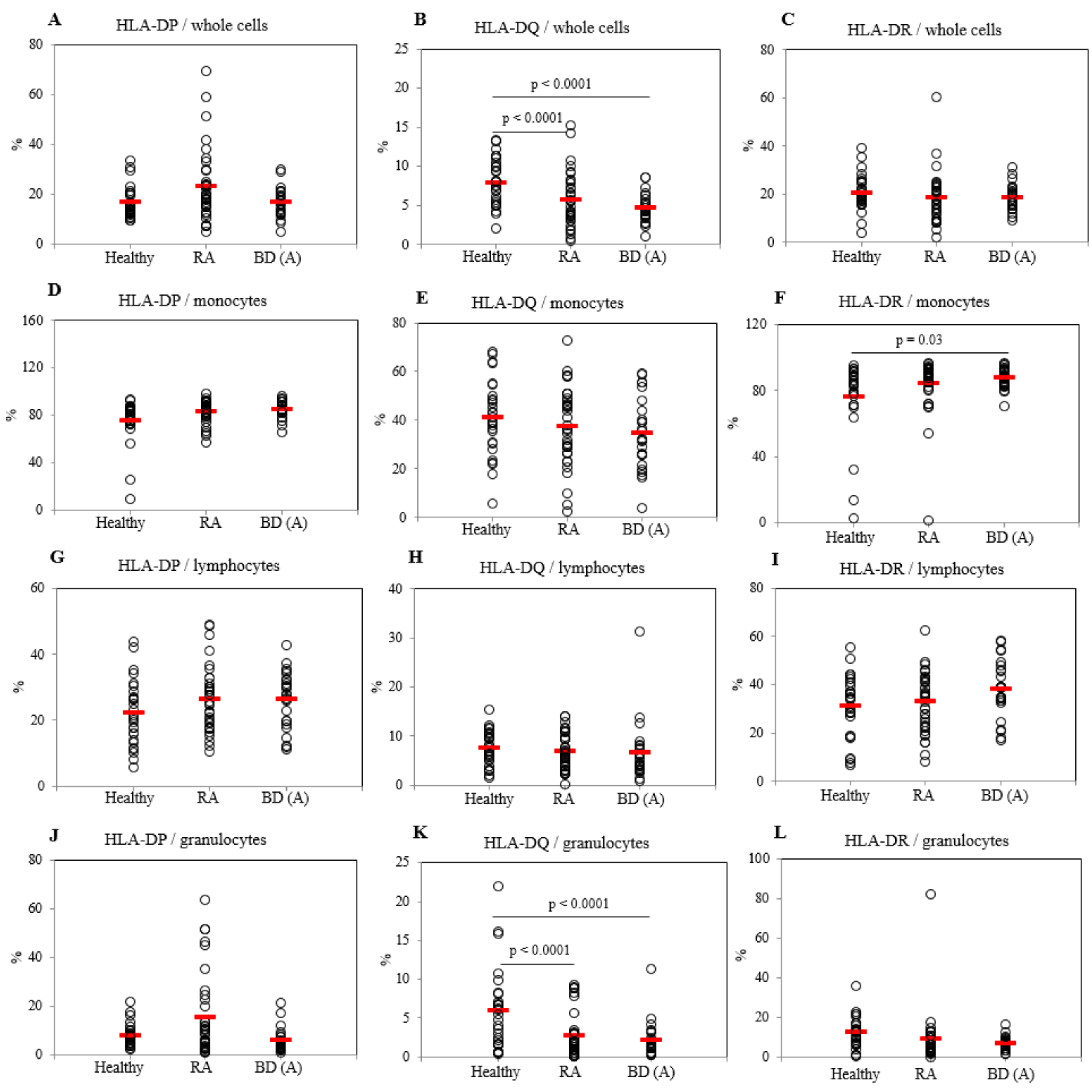
2.2. Frequencies of HLA-DP, DQ, and DR Positive Cells in Active BD Patients
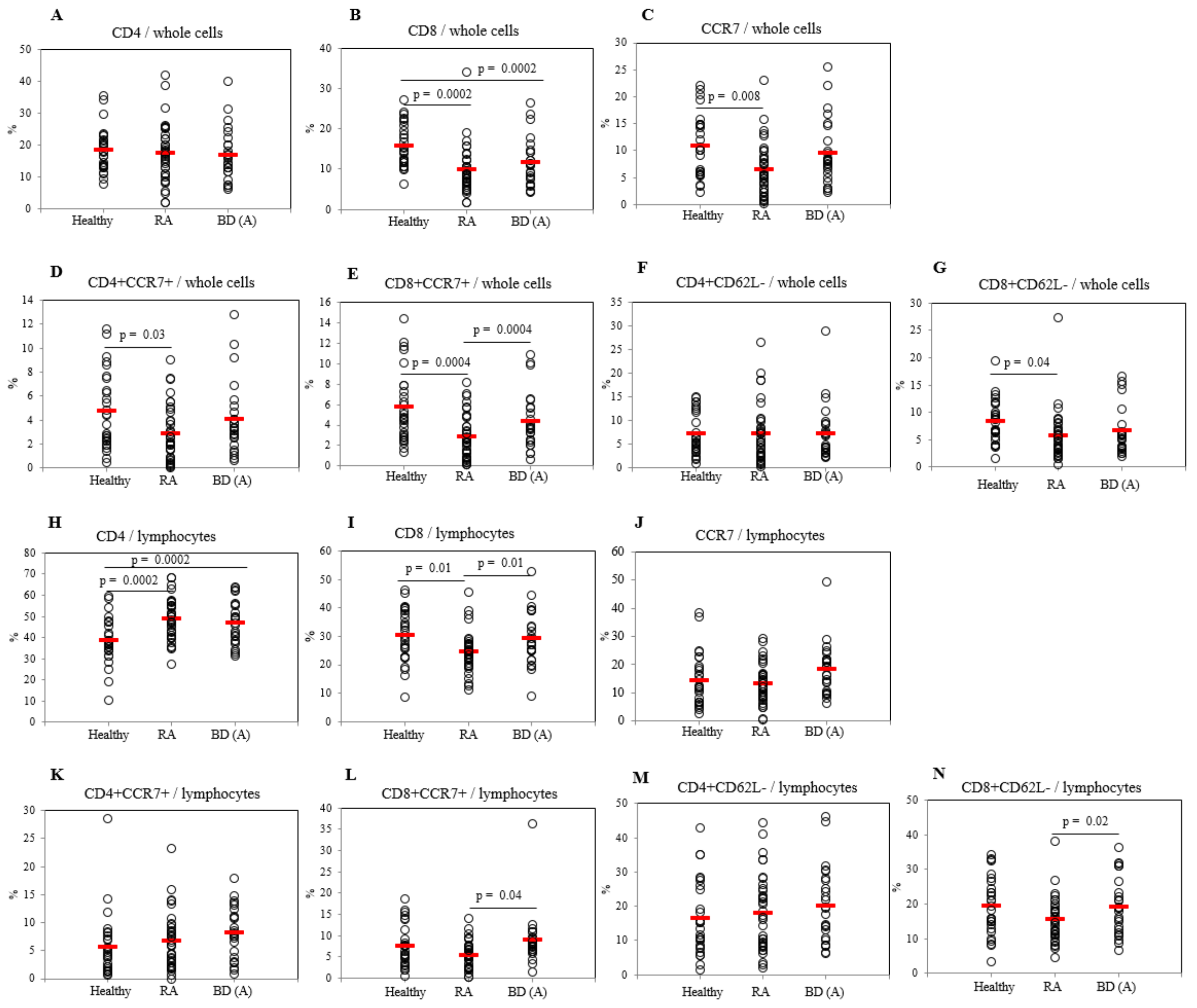
2.3. Differential Frequencies of CCR7+ Cells between Active BD and Healthy Controls
2.4. Differential Frequencies of Memory T Cells between Active and Inactive BD Patients
2.5. Correlation between Frequencies of HLA Class II Subtypes Expressing Cells and Disease Activity Markers in BD Patients
2.6. Correlation between Frequencies of CCR7 Expressing Cells and Disease Activity Markers in BD Patients
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Flow Cytometry Analysis of Patients’ Blood Samples
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, D.Y.; Cho, S.; Choi, M.J.; Sohn, S.; Lee, E.S.; Bang, D. Immunopathogenic Role of Herpes Simplex Virus in Behcet’s Disease. Genet. Res. Int. 2013, 2013, 638273. [Google Scholar] [CrossRef]
- Fietta, P. Behcet’s disease: familial clustering and immunogenetics. Clin. Exp. Rheumatol. 2005, 23, S96–S105. [Google Scholar]
- Parlakgul, G.; Guney, E.; Erer, B.; Kilicaslan, Z.; Direskeneli, H.; Gul, A.; Saruhan-Direskeneli, G. Expression of regulatory receptors on gammadelta T cells and their cytokine production in Behcet’s disease. Arthritis Res. 2013, 15, R15. [Google Scholar] [CrossRef]
- Trachtenberg, E.; Vinson, M.; Hayes, E.; Hsu, Y.M.; Houtchens, K.; Erlich, H.; Klitz, W.; Hsia, Y.; Hollenbach, J. HLA class I (A, B, C) and class II (DRB1, DQA1, DQB1, DPB1) alleles and haplotypes in the Han from southern China. Tissue Antigens 2007, 70, 455–463. [Google Scholar] [CrossRef]
- Hashimoto, K.; Maruyama, H.; Nishiyama, M.; Asaba, K.; Ikeda, Y.; Takao, T.; Iwasaki, Y.; Kumon, Y.; Suehiro, T.; Tanimoto, N.; et al. Susceptibility alleles and haplotypes of human leukocyte antigen DRB1, DQA1, and DQB1 in autoimmune polyglandular syndrome type III in Japanese population. Horm. Res. 2005, 64, 253–260. [Google Scholar] [CrossRef]
- Mangalam, A.K.; Taneja, V.; David, C.S. HLA class II molecules influence susceptibility versus protection in inflammatory diseases by determining the cytokine profile. J. Immunol. 2013, 190, 513–518. [Google Scholar] [CrossRef]
- Meyer, C.G.; May, J.; Schnittger, L. HLA-DP—Part of the concert. Immunol. Today 1997, 18, 58–61. [Google Scholar] [CrossRef]
- Turesson, C. Endothelial expression of MHC class II molecules in autoimmune disease. Curr. Pharm. Des. 2004, 10, 129–143. [Google Scholar] [CrossRef]
- Castillo Palma, M.J.; Sanchez Roman, J.; Ocana Medina, C.; Gonzalez Escribano, M.F.; Nunez Roldan, A.; Lopez-Checa, F. Serologic and molecular HLA typing in patients from Andalucia with Behcet’s disease. Genetic and clinical correlations. Med. Clin. 1996, 106, 121–125. [Google Scholar]
- Heldt, C.; Listing, J.; Sozeri, O.; Blasing, F.; Frischbutter, S.; Muller, B. Differential expression of HLA class II genes associated with disease susceptibility and progression in rheumatoid arthritis. Arthritis Rheum. 2003, 48, 2779–2787. [Google Scholar] [CrossRef]
- Germain, R.N. T-cell development and the CD4-CD8 lineage decision. Nat. Rev. Immunol. 2002, 2, 309–322. [Google Scholar] [CrossRef]
- Butcher, E.C.; Picker, L.J. Lymphocyte homing and homeostasis. Science 1996, 272, 60–66. [Google Scholar] [CrossRef]
- Gunn, M.D.; Kyuwa, S.; Tam, C.; Kakiuchi, T.; Matsuzawa, A.; Williams, L.T.; Nakano, H. Mice lacking expression of secondary lymphoid organ chemokine have defects in lymphocyte homing and dendritic cell localization. J. Exp. Med. 1999, 189, 451–460. [Google Scholar] [CrossRef]
- Mahnke, Y.D.; Brodie, T.M.; Sallusto, F.; Roederer, M.; Lugli, E. The who’s who of T-cell differentiation: human memory T-cell subsets. Eur. J. Immunol. 2013, 43, 2797–2809. [Google Scholar] [CrossRef]
- Shimizu, J.; Yoshikawa, H.; Takada, E.; Hirotsu, C.; Suzuki, N. Unbalanced helper T cell function in Behcet’s disease. Inflamm. Regen. 2011, 31, 296–301. [Google Scholar] [CrossRef]
- Tursen, U. Pathophysiology of the Behcet’s Disease. Pathol. Res. Int. 2012, 2012, 493015. [Google Scholar] [CrossRef]
- Thorsby, E.; Lie, B.A. HLA associated genetic predisposition to autoimmune diseases: Genes involved and possible mechanisms. Transpl. Immunol. 2005, 14, 175–182. [Google Scholar] [CrossRef]
- Eckels, D.D.; Lake, P.; Lamb, J.R.; Johnson, A.H.; Shaw, S.; Woody, J.N.; Hartzman, R.J. SB-restricted presentation of influenza and herpes simplex virus antigens to human T-lymphocyte clones. Nature 1983, 301, 716–718. [Google Scholar] [CrossRef]
- Kamatani, Y.; Wattanapokayakit, S.; Ochi, H.; Kawaguchi, T.; Takahashi, A.; Hosono, N.; Kubo, M.; Tsunoda, T.; Kamatani, N.; Kumada, H.; et al. A genome-wide association study identifies variants in the HLA-DP locus associated with chronic hepatitis B in Asians. Nat. Genet. 2009, 41, 591–595. [Google Scholar] [CrossRef]
- Petersdorf, E.W.; Gooley, T.; Malkki, M.; Anasetti, C.; Martin, P.; Woolfrey, A.; Smith, A.; Mickelson, E.; Hansen, J.A. The biological significance of HLA-DP gene variation in haematopoietic cell transplantation. Br. J. Haematol. 2001, 112, 988–994. [Google Scholar] [CrossRef]
- Hoffman, R.W.; Shaw, S.; Francis, L.C.; Larson, M.G.; Petersen, R.A.; Chylack, L.T.; Glass, D.N. HLA-DP antigens in patients with pauciarticular juvenile rheumatoid arthritis. Arthritis Rheum. 1986, 29, 1057–1062. [Google Scholar] [CrossRef]
- Armstrong, M.A.; Crockard, A.D.; Hawkins, S.A.; Gamble, L.A.; Shah, S.; Bell, A.L. Class II major histocompatibility complex antigen expression on unstimulated and gamma-interferon stimulated monocytes from patients with multiple sclerosis, rheumatoid arthritis and normal controls. Autoimmunity 1991, 9, 261–268. [Google Scholar] [CrossRef]
- Mizuki, N.; Ohno, S.; Kamata, K.; Nakamura, S.; Ishihara, M.; Sato, K.; Inaba, G.; Tsuji, K.; Inoko, H. Immunogenetic mechanism of Behcet’s disease. Nippon Ganka Gakkai Zasshi 1991, 95, 783–789. [Google Scholar]
- Koehler, P.; Wieser, H.; Konitzer, K. Chapter 3 - Treatment of Celiac Disease. In Celiac Disease and Gluten; Koehler, P., Wieser, H., Konitzer, K., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 149–171. [Google Scholar]
- Salgame, P.; Convit, J.; Bloom, B.R. Immunological suppression by human CD8+ T cells is receptor dependent and HLA-DQ restricted. Proc. Natl. Acad. Sci. USA 1991, 88, 2598–2602. [Google Scholar] [CrossRef]
- Fugger, L.; Svejgaard, A. The HLA-DQ7 and -DQ8 associations in DR4-positive rheumatoid arthritis patients. A combined analysis of data available in the literature. Tissue Antigens 1997, 50, 494–500. [Google Scholar] [CrossRef]
- Haag, S.; Tuncel, J.; Thordardottir, S.; Mason, D.E.; Yau, A.C.; Dobritzsch, D.; Backlund, J.; Peters, E.C.; Holmdahl, R. Positional identification of RT1-B (HLA-DQ) as susceptibility locus for autoimmune arthritis. J. Immunol. 2015, 194, 2539–2550. [Google Scholar] [CrossRef]
- Nabozny, G.H.; Baisch, J.M.; Cheng, S.; Cosgrove, D.; Griffiths, M.M.; Luthra, H.S.; David, C.S. HLA-DQ8 transgenic mice are highly susceptible to collagen-induced arthritis: a novel model for human polyarthritis. J. Exp. Med. 1996, 183, 27–37. [Google Scholar] [CrossRef]
- Fugger, L.; Svejgaard, A. Association of MHC and rheumatoid arthritis. HLA-DR4 and rheumatoid arthritis: studies in mice and men. Arthritis Res. 2000, 2, 208–211. [Google Scholar] [CrossRef]
- Mizuki, N.; Inoko, H.; Mizuki, N.; Tanaka, H.; Kera, J.; Tsuiji, K.; Ohno, S. Human leukocyte antigen serologic and DNA typing of Behcet’s disease and its primary association with B51. Invest. Ophthalmol. Vis. Sci. 1992, 33, 3332–3340. [Google Scholar]
- Sun, A.; Hsieh, R.P.; Chu, C.T.; Wang, J.T.; Liu, B.Y.; Chiang, C.P. Some specific human leukocyte antigen (HLA)-DR/DQ haplotypes are more important than individual HLA-DR and -DQ phenotypes for the development of mucocutaneous type of Behcet’s disease and for disease shift from recurrent aphthous stomatitis to mucocutaneous type of Behcet’s disease. J. Oral Pathol. Med. 2001, 30, 402–407. [Google Scholar]
- Haveman, J.W.; van den Berg, A.P.; van den Berk, J.M.; Mesander, G.; Slooff, M.J.; de Leij, L.H.; The, T.H. Low HLA-DR expression on peripheral blood monocytes predicts bacterial sepsis after liver transplantation: Relation with prednisolone intake. Transpl. Infect. Dis. 1999, 1, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Kang, J.S.; Kong, J.M.; Bae, S.; Yu, Y.; Chung, H.; Yu, H.G. Maturation profiles of peripheral blood dendritic cells in patients with endogenous uveitis. Immunol. Lett. 2012, 142, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Lehner, T.; Welsh, K.I.; Batchelor, J.R. The relationship of HLA-B and DR phenotypes to Behcet’s syndrome, recurrent oral ulceration and the class of immune complexes. Immunology 1982, 47, 581–587. [Google Scholar] [PubMed]
- Shang, Y.B.; Zhai, N.; Li, J.P.; Han, S.X.; Ren, Q.S.; Song, F.J.; Chen, H.D. Study on association between polymorphism of HLA-DRB1 alleles and Behcet’s disease. J. Eur. Acad. Derm. Venereol. 2009, 23, 1419–1422. [Google Scholar] [CrossRef]
- Kawashima, D.; Oshitani, N.; Jinno, Y.; Watanabe, K.; Nakamura, S.; Higuchi, K.; Arakawa, T. Augmented expression of secondary lymphoid tissue chemokine and EBI1 ligand chemokine in Crohn’s disease. J. Clin. Pathol. 2005, 58, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- McNamee, E.N.; Masterson, J.C.; Veny, M.; Collins, C.B.; Jedlicka, P.; Byrne, F.R.; Ng, G.Y.; Rivera-Nieves, J. Chemokine receptor CCR7 regulates the intestinal T(H)1/T(H)17/T(reg) balance during Crohn’s-like murine ileitis. J. Leukoc. Biol. 2015, 97, 1011–1022. [Google Scholar] [CrossRef]
- Pender, M.P.; Csurhes, P.A.; Pfluger, C.M.; Burrows, S.R. Deficiency of CD8+ effector memory T cells is an early and persistent feature of multiple sclerosis. Mult. Scler. 2014, 20, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Bjorkdahl, O.; Barber, K.A.; Brett, S.J.; Daly, M.G.; Plumpton, C.; Elshourbagy, N.A.; Tite, J.P.; Thomsen, L.L. Characterization of CC-chemokine receptor 7 expression on murine T cells in lymphoid tissues. Immunology 2003, 110, 170–179. [Google Scholar] [CrossRef]
- Von Andrian, U.H.; Mackay, C.R. T-cell function and migration. Two sides of the same coin. N. Engl. J. Med. 2000, 343, 1020–1034. [Google Scholar] [CrossRef]
- Arbones, M.L.; Ord, D.C.; Ley, K.; Ratech, H.; Maynard-Curry, C.; Otten, G.; Capon, D.J.; Tedder, T.F. Lymphocyte homing and leukocyte rolling and migration are impaired in L-selectin-deficient mice. Immunity 1994, 1, 247–260. [Google Scholar] [CrossRef]
- Sallusto, F.; Lenig, D.; Forster, R.; Lipp, M.; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 1999, 401, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, A.; Mueller, Y.M.; Thomas, P.; Bojczuk, P.; O’Connors, C.; Katsikis, P.D. Decreased effector memory CD45RA+ CD62L- CD8+ T cells and increased central memory CD45RA- CD62L+ CD8+ T cells in peripheral blood of rheumatoid arthritis patients. Arthritis Res. 2003, 5, R91–R96. [Google Scholar] [CrossRef]
- Ellingsen, T.; Hansen, I.; Thorsen, J.; Moller, B.K.; Tarp, U.; Lottenburger, T.; Andersen, L.S.; Skjodt, H.; Pedersen, J.K.; Lauridsen, U.B.; et al. Upregulated baseline plasma CCL19 and CCR7 cell-surface expression on monocytes in early rheumatoid arthritis normalized during treatment and CCL19 correlated with radiographic progression. Scand. J. Rheumatol. 2014, 43, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liu, F.; Wang, Q.J.; Rosenberg, S.A.; Morgan, R.A. The Shedding of CD62L (L-Selectin) Regulates the Acquisition of Lytic Activity in Human Tumor Reactive T Lymphocytes. PLoS ONE 2011, 6, e22560. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Choi, B.; Sayeed, H.M.; Suh, C.H.; Kim, Y.W.; Kim, H.A.; Sohn, S. Characteristic patterns of HLA presentation and T cell differentiation in adult-onset Still’s disease. Int. J. Immunopathol. Pharm. 2018, 32, 2058738418791284. [Google Scholar] [CrossRef]
- Criteria for diagnosis of Behcet’s disease. International Study Group for Behcet’s Disease. Lancet 1990, 335, 1078–1080.
- Arnett, F.C.; Edworthy, S.M.; Bloch, D.A.; McShane, D.J.; Fries, J.F.; Cooper, N.S.; Healey, L.A.; Kaplan, S.R.; Liang, M.H.; Luthra, H.S.; et al. The American Rheumatism Association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheum. 1988, 31, 315–324. [Google Scholar] [CrossRef]
- Choi, B.; Suh, C.H.; Kim, H.A.; Sayeed, H.M.; Sohn, S. The Correlation of CD206, CD209, and Disease Severity in Behcet’s Disease with Arthritis. Mediat. Inflamm. 2017, 2017, 7539529. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Patients | Age | OU | GU | Arth | EN | Disease Severity Score | Leukocyte | ESR | CRP | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | ||||||
| N = 25 (M = 5, F = 20) | 48.8 ± 7.6 | 13 (52.0%) | 6 (24.0%) | 25 (100%) | 7 (28.0%) | 3.04 ± 1.1 | 1.27 ± 1.0 | 8008.0 ± 3480.1 | 6836.4 ± 1937.1 | 29.8 ± 25.8 | 18.3 ±13.6 | 1.01 ± 2.02 | 0.26 ± 0.37 |
| P value | P = 0.007 | P = 0.168 | P = 0.083 | P = 0.314 | |||||||||
| Ordrer | Colchicine | Steroid | AZP | Bucillamine | HCQ | SZP | NSAIDs | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | 1st | 2nd | |
| Number (%) | 19 (76.2%) | 9 (81.8%) | 21 (84.0%) | 7 (63.6%) | 7 (28.0%) | 3 (27.3%) | 1 (8.0%) | 2 (18.2%) | 8 (32.0%) | 4 (36.4%) | 10 (40.0%) | 5 (45.5%) | 16 (64.0%) | 7 (63.6%) |
| Disease Activity Markers | Correlation Coefficient, r (p-value) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HLA-DP | HLA-DQ | HLA-DR | |||||||||||
| PBL | Monocyte | Lymphocyte | Granulocyte | PBL | Monocyte | Lymphocyte | Granulocyte | PBL | Monocyte | Lymphocyte | Granulocyte | ||
| Disease severity score | −0.015 (0.945) | 0.202 (0.334) | 0.138 (0.510) | 0.193 (0.355) | −0.033 (0.876) | 0.365 (0.073) | 0.091 (0.665) | −0.088 (0.677) | −0.004 (0.984) | 0.379 (0.062) | 0.374 (0.066) | −0.041 (0.847) | |
| Leukocyte number | 0.025 (0.907) | −0.226 (0.278) | −0.169 (0.419) | 0.123 (0.557) | −0.021 (0.919) | −0.323 (0.115) | −0.394 (0.051) | 0.364 (0.073) | 0.218 (0.294) | −0.104 (0.622) | −0.117 (0.579) | 0.289 (0.161) | |
| ESR | −0.142 (0.498) | 0.044 (0.834) | −0.289 (0.161) | 0.180 (0.389) | −0.225 (0.279) | 0.055 (0.792) | 0.010 (0.964) | −0.230 (0.269) | −0.33 (0.059) | 0.107 (0.610) | 0.060 (0.777) | −0.034 (0.872) | |
| CRP | −0.179 (0.393) | 0.213 (0.306) | −0.289 (0.161) | 0.001 (0.996) | −0.438 (0.029) | −0.213 (0.307) | −0.290 (0.160) | −0.078 (0.709) | −0.234 (0.260) | 0.385 (0.058) | −0.005 (0.982) | −0.062 (0.769) | |
| Disease Activity Markers | Correlation Coefficient, r (p-value) | ||||||
|---|---|---|---|---|---|---|---|
| CD4+ | CD8+ | CCR7+ | CD4+CCR7+ | CD8+CCR7+ | CD4+CD62L- | CD8+CD62L- | |
| Leukocyte number | 0.316 (0.124) | 0.378 (0.063) | 0.431 (0.031) | 0.351 (0.086) | 0.417 (0.038) | 0.325 (0.113) | 0.266 (0.198) |
| ESR | −0.382 (0.060) | −0.376 (0.064) | −0.258 (0.213) | −0.385 (0.057) | −0.391 (0.053) | −0.267 (0.197) | −0.339 (0.097) |
| CRP | −0.161 (0.442) | −0.198 (0.342) | −0.119 (0.572) | −0.187 (0.370) | −0.184 (0.380) | −0.063 (0.764) | −0.109 (0.603) |
| Correlation Coefficient, r (p-value) | ||||||
|---|---|---|---|---|---|---|
| CD4+ Naïve T | CD4+ Effector Memory T | CD4+ Central Memory T | CD8+ Naïve T | CD8+ Effector Memory T | CD8+ Central Memory T | |
| Disease Severity Score | 0.259 (0.212) | −0.358 (0.079) | 0.075 (0.723) | 0.463 (0.020) | −0.214 (0.305) | 0.163 (0.435) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, S.M.S.; Kim, H.-A.; Choi, B.; Jung, J.-Y.; Lee, S.-M.; Suh, C.-H.; Sohn, S. Differences in Expression of Human Leukocyte Antigen Class II Subtypes and T Cell Subsets in Behçet’s Disease with Arthritis. Int. J. Mol. Sci. 2019, 20, 5044. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205044
Islam SMS, Kim H-A, Choi B, Jung J-Y, Lee S-M, Suh C-H, Sohn S. Differences in Expression of Human Leukocyte Antigen Class II Subtypes and T Cell Subsets in Behçet’s Disease with Arthritis. International Journal of Molecular Sciences. 2019; 20(20):5044. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205044
Chicago/Turabian StyleIslam, S. M. Shamsul, Hyoun-Ah Kim, Bunsoon Choi, Ju-Yang Jung, Sung-Min Lee, Chang-Hee Suh, and Seonghyang Sohn. 2019. "Differences in Expression of Human Leukocyte Antigen Class II Subtypes and T Cell Subsets in Behçet’s Disease with Arthritis" International Journal of Molecular Sciences 20, no. 20: 5044. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20205044