Vitamin E but Not GSH Decreases Reactive Oxygen Species Accumulation and Enhances Sperm Production during In Vitro Maturation of Frozen-Thawed Prepubertal Mouse Testicular Tissue

 , and
, and
Abstract
:1. Introduction
2. Results
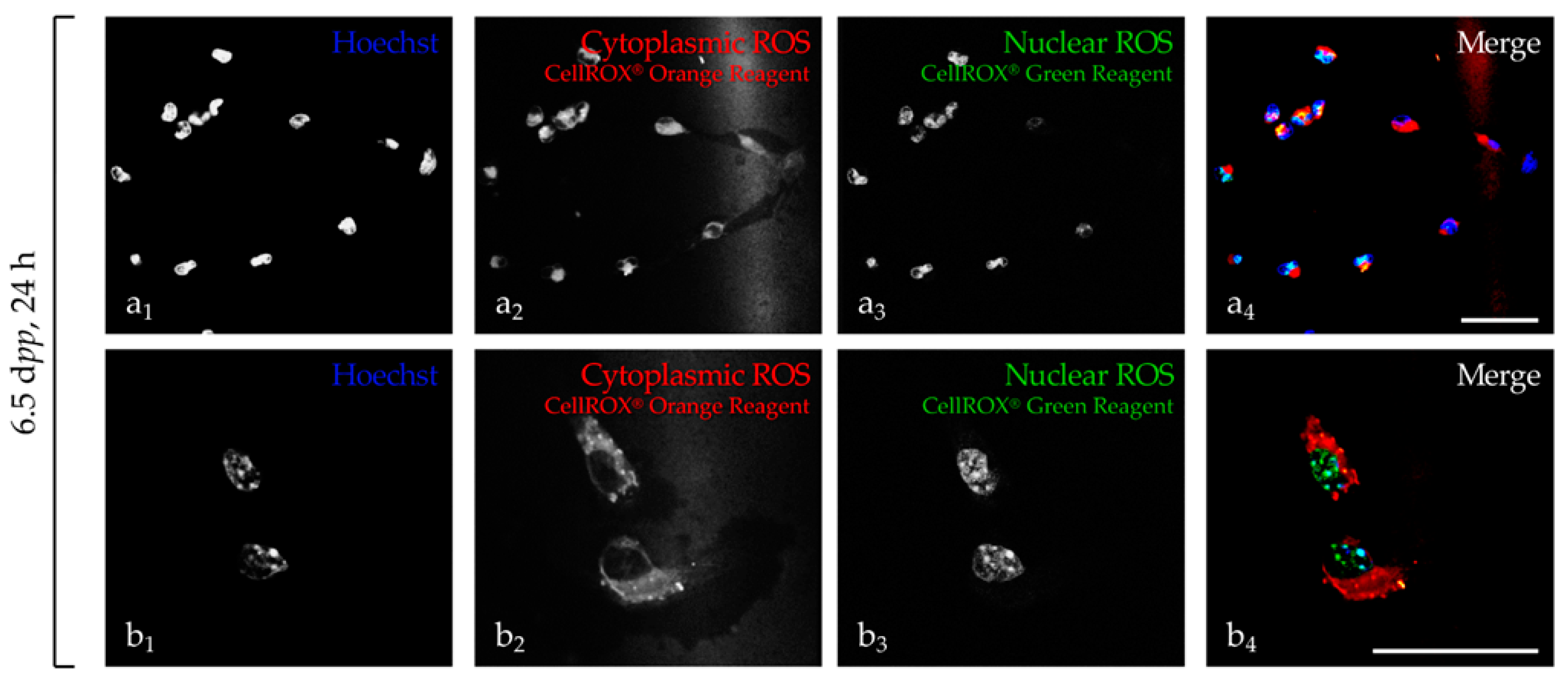
2.1. Detection of ROS in Cell Suspension and in Testicular Explants
2.1.1. Detection in Testicular Cell Suspension
2.1.2. Detection in Testicular Explants
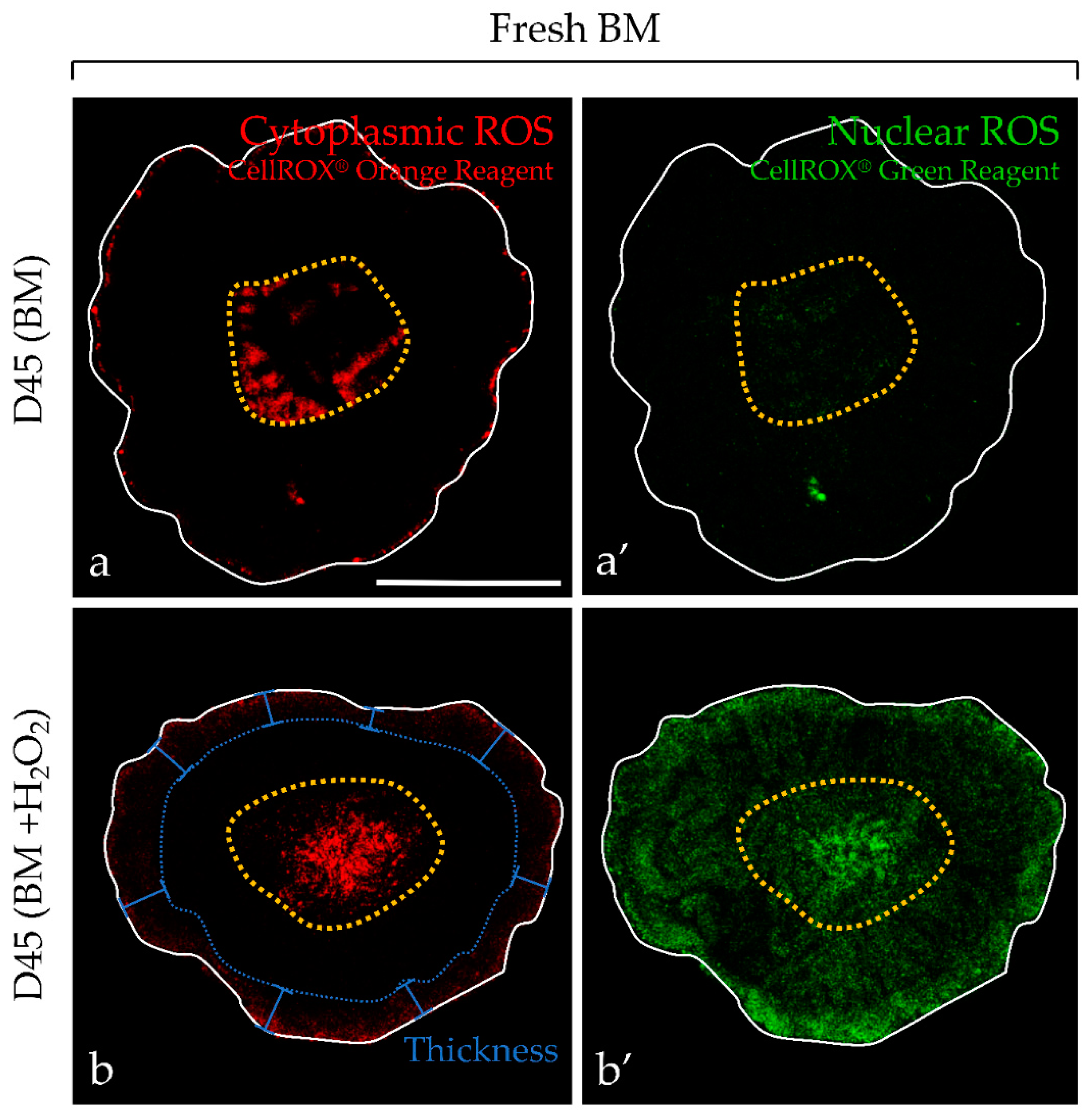
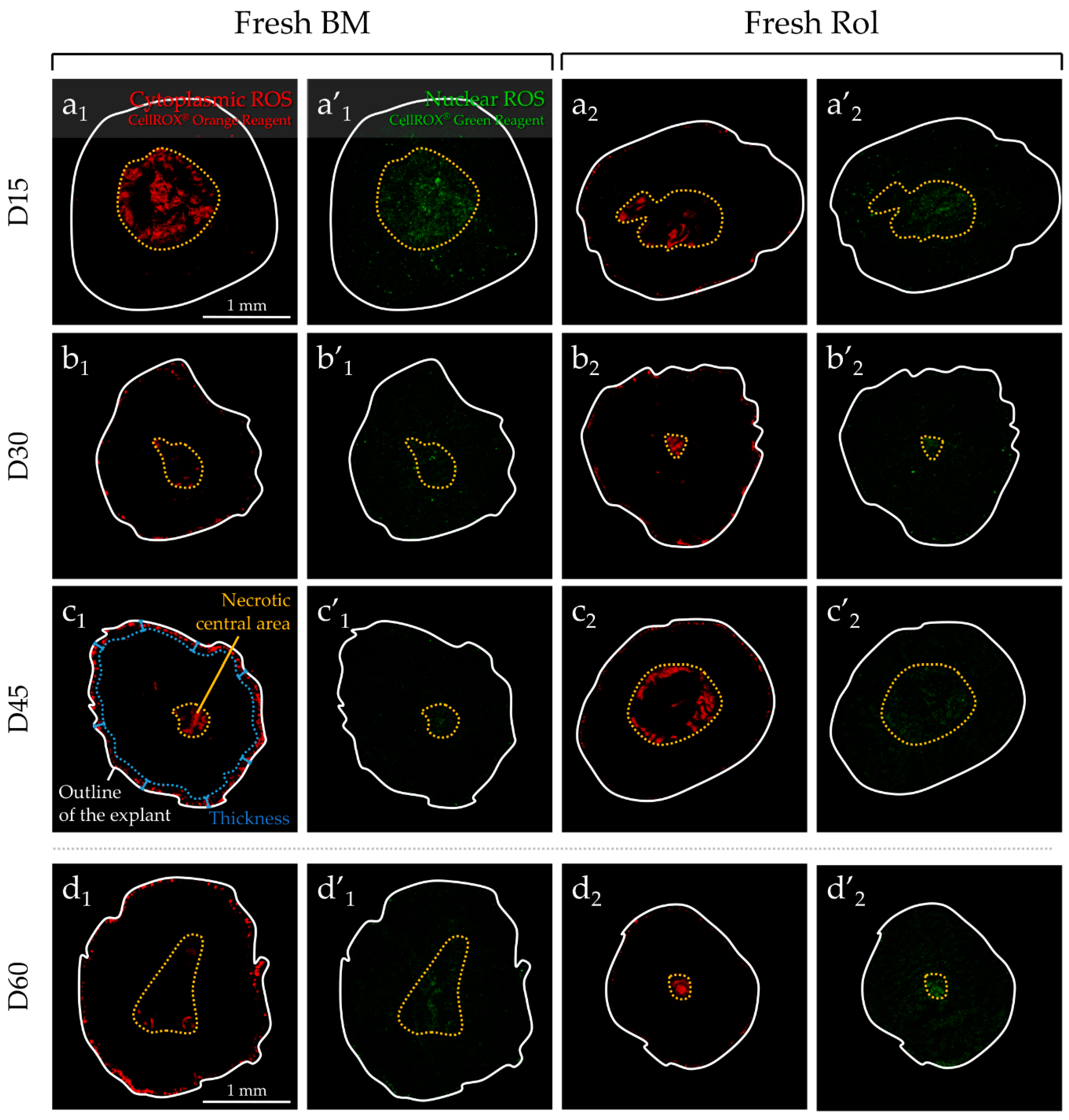
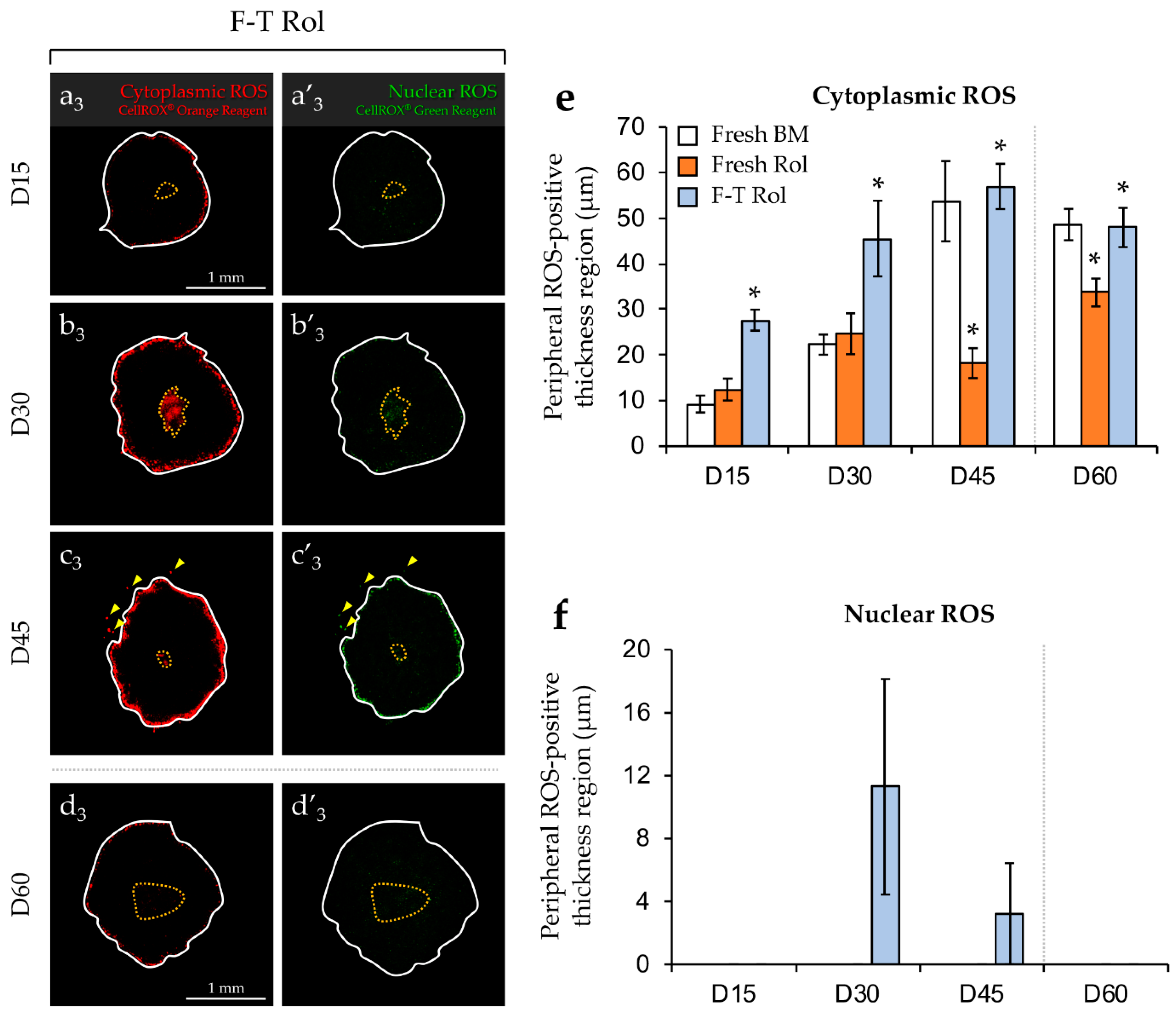
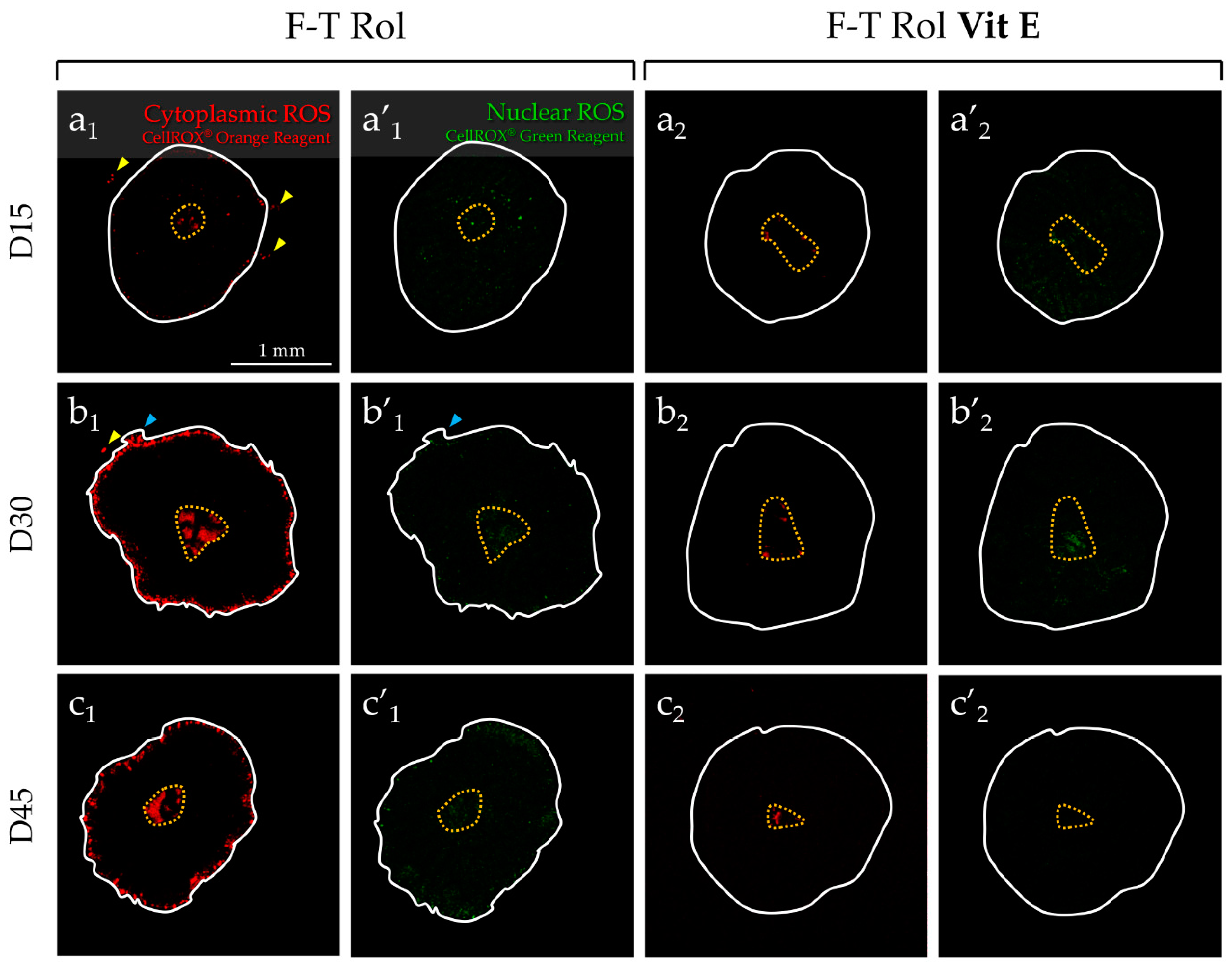
2.2. Analysis of ROS Generation in Fresh or F-T Mouse Prepubertal Testicular Tissue during In Vitro Maturation without and with an Antioxidant Supplementation
2.2.1. Evaluation of ROS without Antioxidant Supplementation and Long-Term Culture (60 days)
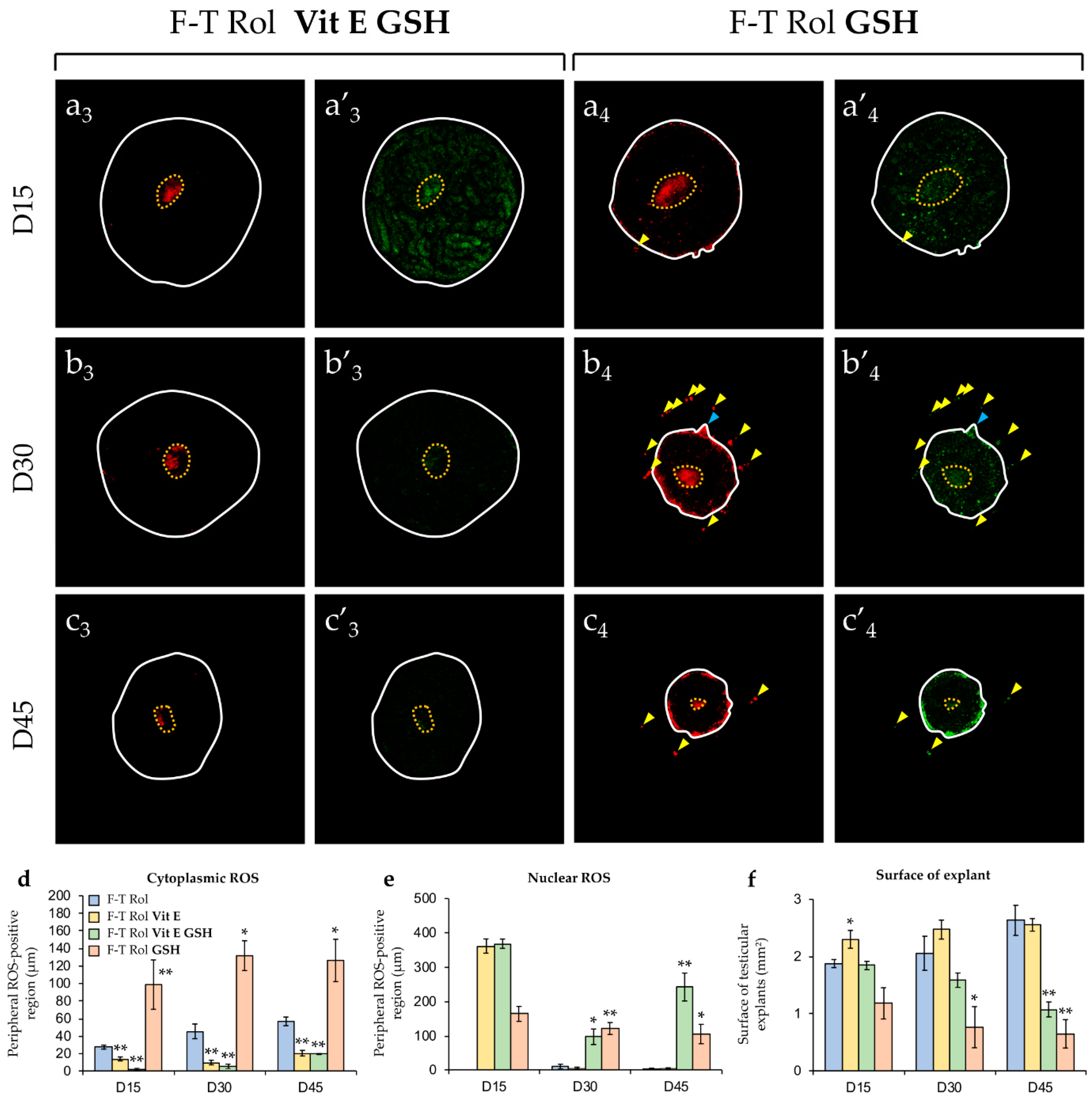
2.2.2. Evaluation of Antioxidant Supplementation
2.2.3. Growth of F-T Testicular Explants during In Vitro Culture with an Antioxidant Supplementation
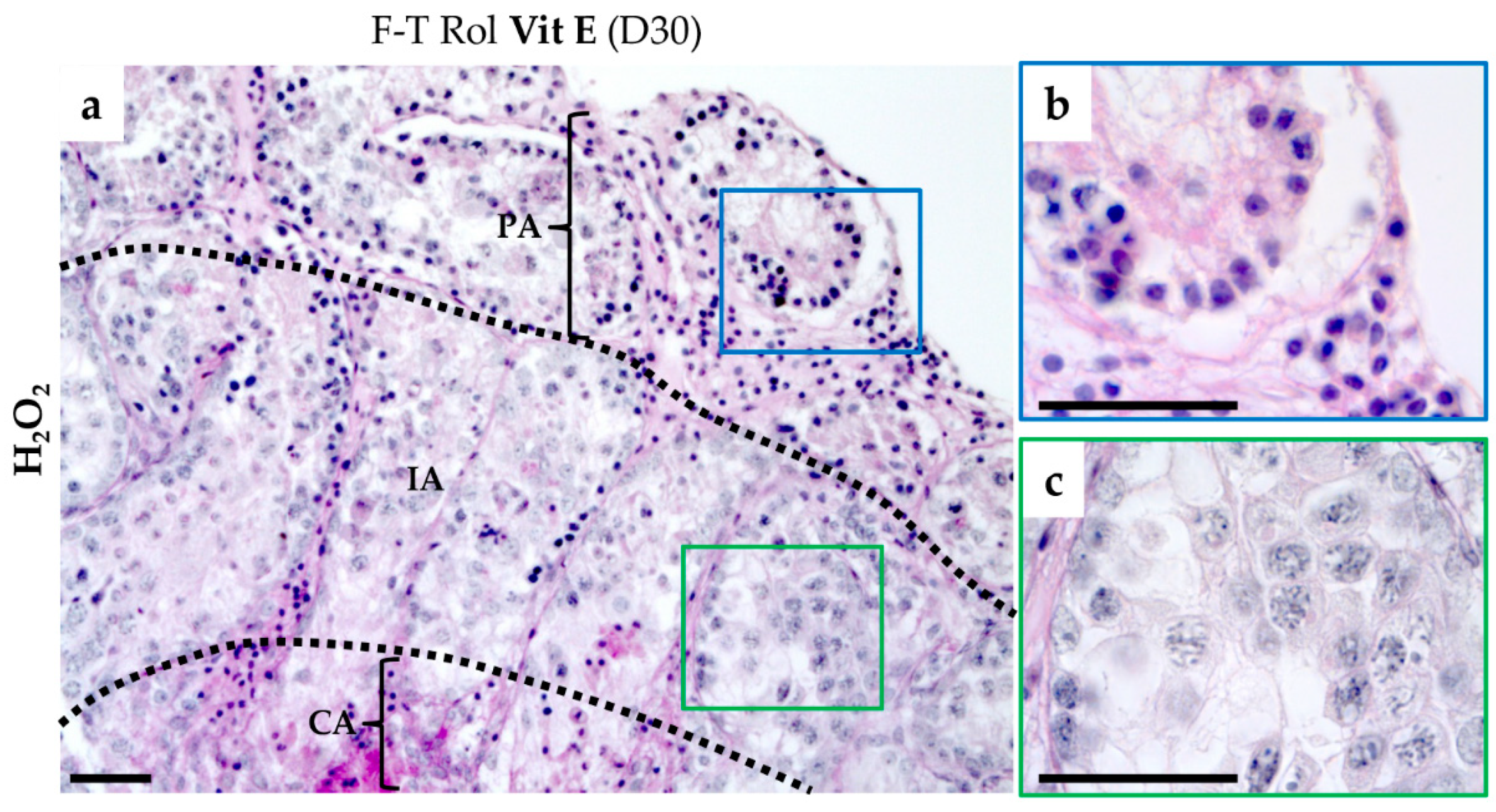
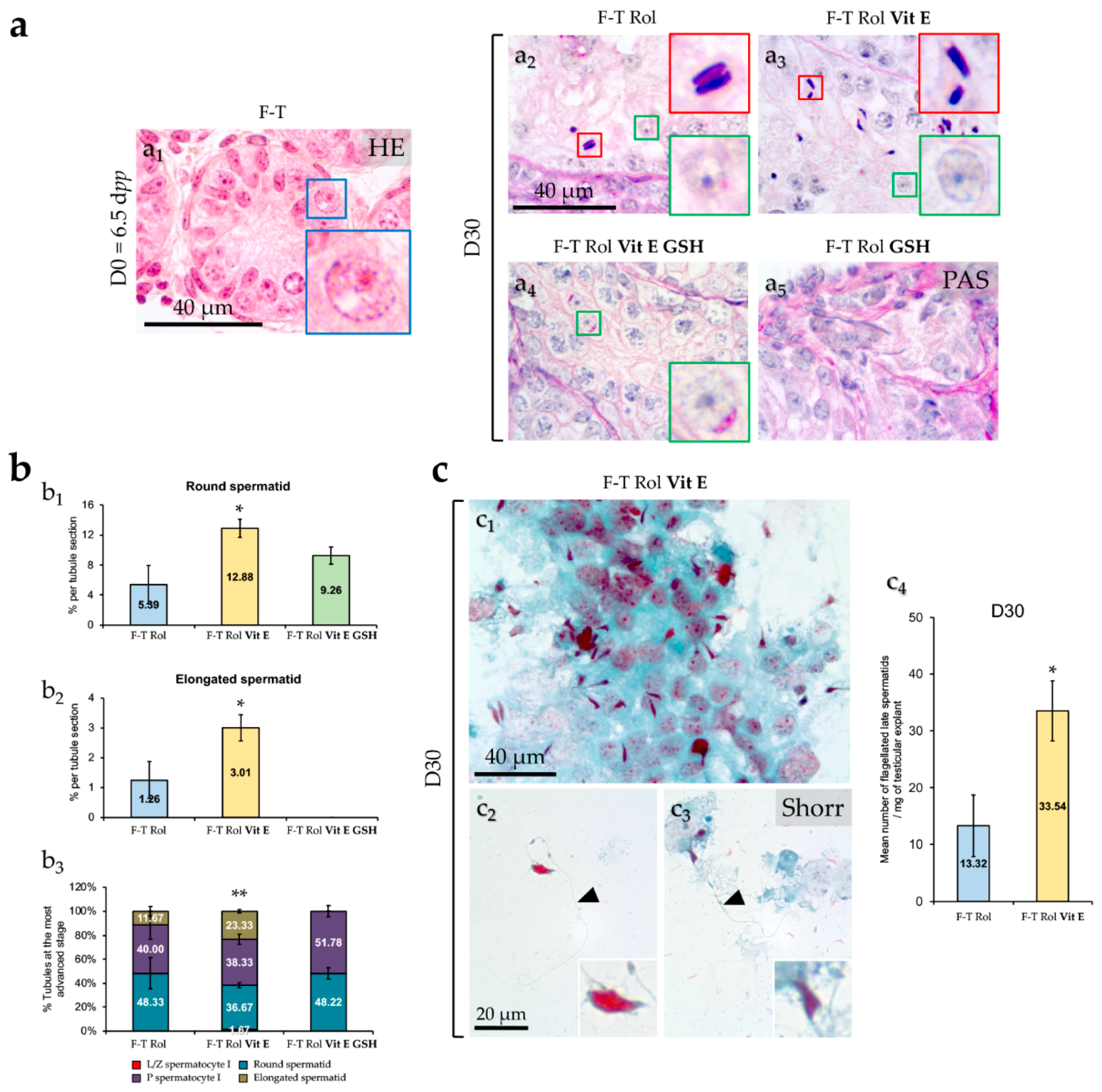
2.3. Spermatogenesis Evaluation
2.3.1. Testicular Germ Cells Differentiation
2.3.2. Identification and Enumeration of Spermatozoa
3. Discussion
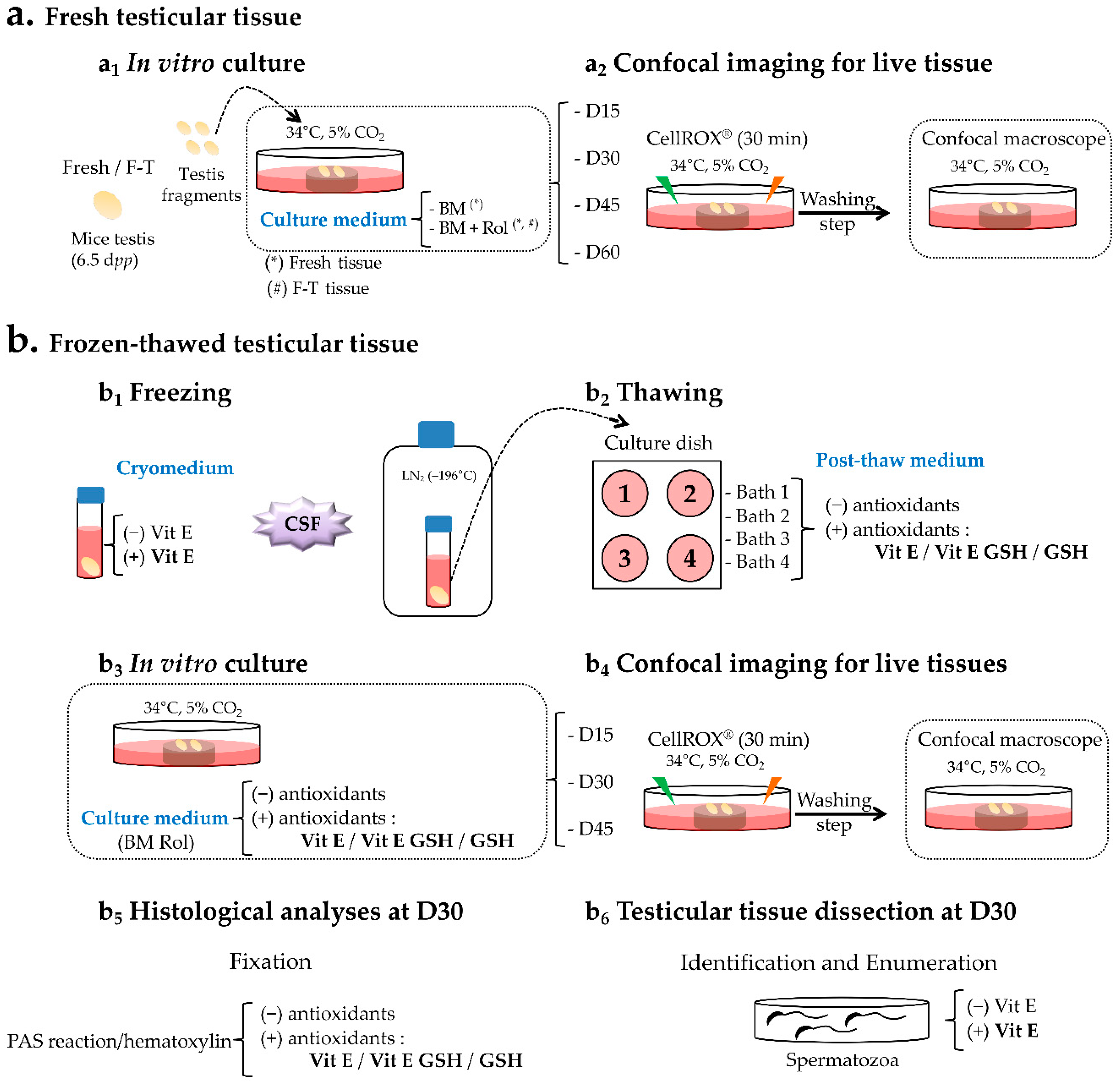
4. Materials and Methods
4.1. Mice and Testes Collection
4.2. Media and Reagents
4.3. Cryopreservation Procedure
4.4. Organ Culture
4.5. Confocal Live Imaging
4.5.1. ROS Measurement in Germinal Cells
4.5.2. ROS Measurement in Testicular Tissue Explants
4.6. Histological Analysis
4.7. Spermatozoa Enumeration
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BM | basal medium |
| CA | central area |
| CSF | controlled slow freezing |
| D | day |
| dpp | days post-partum |
| FCS | fetal calf serum |
| F-T | frozen-thawed |
| GSH | reduced glutathione |
| h | hour |
| HE | hematoxylin eosin |
| H2O2 | hydrogen peroxide |
| IA | intermediate area |
| KSR | KnockOut Serum Replacement |
| LN2 | liquid nitrogen |
| L/Z | leptotene/zygotene |
| min | minutes |
| P | pachytene |
| PA | peripheral area |
| PAS | periodic acid Schiff |
| Rol | retinol |
| ROS | reactive oxygen species |
| s.e.m. | standard error of the mean |
| SSC | spermatogonial stem cell |
| Vit E | vitamin E |
References
- Picton, H.M.; Wyns, C.; Anderson, R.A.; Goossens, E.; Jahnukainen, K.; Kliesch, S.; Mitchell, R.T.; Pennings, G.; Rives, N.; Tournaye, H.; et al. A European perspective on testicular tissue cryopreservation for fertility preservation in prepubertal and adolescent boys. Hum. Reprod. 2015, 30, 2463–2475. [Google Scholar] [CrossRef]
- Wyns, C.; Van Langendonckt, A.; Wese, F.-X.; Donnez, J.; Curaba, M. Long-term spermatogonial survival in cryopreserved and xenografted immature human testicular tissue. Hum. Reprod. 2008, 23, 2402–2414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onofre, J.; Baert, Y.; Faes, K.; Goossens, E. Cryopreservation of testicular tissue or testicular cell suspensions: A pivotal step in fertility preservation. Hum. Reprod. Update 2016, 22, 744–761. [Google Scholar] [CrossRef]
- Curaba, M.; Verleysen, M.; Amorim, C.A.; Dolmans, M.-M.; Van Langendonckt, A.; Hovatta, O.; Wyns, C.; Donnez, J. Cryopreservation of prepubertal mouse testicular tissue by vitrification. Fertil. Steril. 2011, 95, 1229–1234.e1. [Google Scholar] [CrossRef]
- Poels, J.; Van Langendonckt, A.; Many, M.-C.; Wese, F.-X.; Wyns, C. Vitrification preserves proliferation capacity in human spermatogonia. Hum. Reprod. 2013, 28, 578–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, A.; Allamaneni, S.S.R. Role of free radicals in female reproductive diseases and assisted reproduction. Reprod. Biomed. Online 2004, 9, 338–347. [Google Scholar] [CrossRef]
- Shi, Y.; Buffenstein, R.; Pulliam, D.A.; Van Remmen, H. Comparative studies of oxidative stress and mitochondrial function in aging. Integr. Comp. Biol. 2010, 50, 869–879. [Google Scholar] [CrossRef]
- Morimoto, H.; Iwata, K.; Ogonuki, N.; Inoue, K.; Atsuo, O.; Kanatsu-Shinohara, M.; Morimoto, T.; Yabe-Nishimura, C.; Shinohara, T. ROS are required for mouse spermatogonial stem cell self-renewal. Cell Stem Cell 2013, 12, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef]
- Saalu, L.C. The incriminating role of reactive oxygen species in idiopathic male infertility: An evidence based evaluation. Pak. J. Biol. Sci. 2010, 13, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Trocchia, S.; Abdel-Gawad, F.K.; Ciarcia, G. Roles of reactive oxygen species in the spermatogenesis regulation. Front. Endocrinol. 2014, 5, 56. [Google Scholar] [CrossRef] [PubMed]
- Di Santo, M.; Tarozzi, N.; Nadalini, M.; Borini, A. Human Sperm Cryopreservation: Update on Techniques, Effect on DNA Integrity, and Implications for ART. Adv. Urol. 2012, 2012, 854837. [Google Scholar] [CrossRef] [PubMed]
- Thomson, L.K.; Fleming, S.D.; Aitken, R.J.; De Iuliis, G.N.; Zieschang, J.-A.; Clark, A.M. Cryopreservation-induced human sperm DNA damage is predominantly mediated by oxidative stress rather than apoptosis. Hum. Reprod. 2009, 24, 2061–2070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.W.; Zhang, H.; Ikemoto, I.; Anderson, D.J.; Loughlin, K.R. Reactive oxygen species generation by seminal cells during cryopreservation. Urology 1997, 49, 921–925. [Google Scholar] [CrossRef]
- Rahimi, G.; Isachenko, E.; Sauer, H.; Isachenko, V.; Wartenberg, M.; Hescheler, J.; Mallmann, P.; Nawroth, F. Effect of different vitrification protocols for human ovarian tissue on reactive oxygen species and apoptosis. Reprod. Fertil. Dev. 2003, 15, 343–349. [Google Scholar] [CrossRef]
- Fabbri, R.; Vicenti, R.; Martino, N.A.; Dell’Aquila, M.E.; Pasquinelli, G.; Macciocca, M.; Magnani, V.; Paradisi, R.; Venturoli, S. Confocal laser scanning microscopy analysis of bioenergetic potential and oxidative stress in fresh and frozen-thawed human ovarian tissue from oncologic patients. Fertil. Steril. 2014, 101, 795–804.e1. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M.; Bedaiwy, M.A.; Banerjee, J.; Alvarez, J.G. Oxidative stress in an assisted reproductive techniques setting. Fertil. Steril. 2006, 86, 503–512. [Google Scholar] [CrossRef]
- Arkoun, B.; Dumont, L.; Milazzo, J.-P.; Way, A.; Bironneau, A.; Wils, J.; Macé, B.; Rives, N. Retinol improves in vitro differentiation of pre-pubertal mouse spermatogonial stem cells into sperm during the first wave of spermatogenesis. PLoS ONE 2015, 10, e0116660. [Google Scholar] [CrossRef]
- Dumont, L.; Oblette, A.; Rondanino, C.; Jumeau, F.; Bironneau, A.; Liot, D.; Duchesne, V.; Wils, J.; Rives, N. Vitamin A prevents round spermatid nuclear damage and promotes the production of motile sperm during in vitro maturation of vitrified pre-pubertal mouse testicular tissue. Mol. Hum. Reprod. 2016, 22, 819–832. [Google Scholar]
- Dumont, L.; Chalmel, F.; Oblette, A.; Berby, B.; Rives, A.; Duchesne, V.; Rondanino, C.; Rives, N. Evaluation of apoptotic- and autophagic-related protein expressions before and after IVM of fresh, slow-frozen and vitrified pre-pubertal mouse testicular tissue. Mol. Hum. Reprod. 2017, 23, 738–754. [Google Scholar] [CrossRef]
- Sato, T.; Katagiri, K.; Gohbara, A.; Inoue, K.; Ogonuki, N.; Ogura, A.; Kubota, Y.; Ogawa, T. In vitro production of functional sperm in cultured neonatal mouse testes. Nature 2011, 471, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Komeya, M.; Kimura, H.; Nakamura, H.; Yokonishi, T.; Sato, T.; Kojima, K.; Hayashi, K.; Katagiri, K.; Yamanaka, H.; Sanjo, H.; et al. Long-term ex vivo maintenance of testis tissues producing fertile sperm in a microfluidic device. Sci. Rep. 2016, 6, 21472. [Google Scholar] [CrossRef] [PubMed]
- Komeya, M.; Hayashi, K.; Nakamura, H.; Yamanaka, H.; Sanjo, H.; Kojima, K.; Sato, T.; Yao, M.; Kimura, H.; Fujii, T.; et al. Pumpless microfluidic system driven by hydrostatic pressure induces and maintains mouse spermatogenesis in vitro. Sci. Rep. 2017, 7, 15459. [Google Scholar] [CrossRef] [PubMed]
- Arkoun, B.; Dumont, L.; Milazzo, J.-P.; Rondanino, C.; Bironneau, A.; Wils, J.; Rives, N. Does soaking temperature during controlled slow freezing of pre-pubertal mouse testes influence course of in vitro spermatogenesis? Cell Tissue Res. 2016, 364, 661–674. [Google Scholar] [CrossRef]
- Dumont, L.; Arkoun, B.; Jumeau, F.; Milazzo, J.-P.; Bironneau, A.; Liot, D.; Wils, J.; Rondanino, C.; Rives, N. Assessment of the optimal vitrification protocol for pre-pubertal mice testes leading to successful in vitro production of flagellated spermatozoa. Andrology 2015, 3, 611–625. [Google Scholar] [CrossRef] [Green Version]
- Niki, E. Interaction of ascorbate and alpha-tocopherol. Ann. N. Y. Acad. Sci. 1987, 498, 186–199. [Google Scholar] [CrossRef]
- Taylor, K.; Roberts, P.; Sanders, K.; Burton, P. Effect of antioxidant supplementation of cryopreservation medium on post-thaw integrity of human spermatozoa. Reprod. Biomed. Online 2009, 18, 184–189. [Google Scholar] [CrossRef]
- Kalthur, G.; Raj, S.; Thiyagarajan, A.; Kumar, S.; Kumar, P.; Adiga, S.K. Vitamin E supplementation in semen-freezing medium improves the motility and protects sperm from freeze-thaw-induced DNA damage. Fertil. Steril. 2011, 95, 1149–1151. [Google Scholar] [CrossRef]
- Keshtgar, S.; Fanaei, H.; Bahmanpour, S.; Azad, F.; Ghannadi, A.; Kazeroni, M. In vitro effects of α-tocopherol on teratozoospermic semen samples. Andrologia 2012, 44, 721–727. [Google Scholar] [CrossRef]
- Hu, J.-H.; Zhao, X.-L.; Tian, W.-Q.; Zan, L.-S.; Li, Q.-W. Effects of vitamin E supplementation in the extender on frozen-thawed bovine semen preservation. Anim. Int. J. Anim. Biosci. 2011, 5, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Virk, G.; Ong, C.; du Plessis, S.S. Effect of oxidative stress on male reproduction. World J. Men’s Health 2014, 32, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Aliakbari, F.; Sedighi Gilani, M.A.; Yazdekhasti, H.; Koruji, M.; Asgari, H.R.; Baazm, M.; Izadyar, F.; Kharrazi Nejad, E.; Khanezad, M.; Abbasi, M. Effects of antioxidants, catalase and α-tocopherol on cell viability and oxidative stress variables in frozen-thawed mice spermatogonial stem cells. Artif. Cells Nanomed. Biotechnol. 2017, 45, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Aliakbari, F.; Gilani, M.A.S.; Amidi, F.; Baazm, M.; Korouji, M.; Izadyar, F.; Yazdekhasti, H.; Abbasi, M. Improving the Efficacy of Cryopreservation of Spermatogonia Stem Cells by Antioxidant Supplements. Cell. Reprogramming 2016, 18, 87–95. [Google Scholar] [CrossRef]
- Gadea, J.; Molla, M.; Sellés, E.; Marco, M.A.; Garcia-Vázquez, F.A.; Gardon, J.C. Reduced glutathione content in human sperm is decreased after cryopreservation: Effect of the addition of reduced glutathione to the freezing and thawing extenders. Cryobiology 2011, 62, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, J.F.; Chatterjee, S.; Sirard, M.A.; Gagnon, C. Levels of antioxidant defenses are decreased in bovine spermatozoa after a cycle of freezing and thawing. Mol. Reprod. Dev. 2000, 55, 282–288. [Google Scholar] [CrossRef]
- Gadea, J.; Sellés, E.; Marco, M.A.; Coy, P.; Matás, C.; Romar, R.; Ruiz, S. Decrease in glutathione content in boar sperm after cryopreservation. Effect of the addition of reduced glutathione to the freezing and thawing extenders. Theriogenology 2004, 62, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Stradaioli, G.; Noro, T.; Sylla, L.; Monaci, M. Decrease in glutathione (GSH) content in bovine sperm after cryopreservation: Comparison between two extenders. Theriogenology 2007, 67, 1249–1255. [Google Scholar] [CrossRef]
- Hong, Z.; Hailing, L.; Hui, M.; Guijie, Z. Effect of vitamin E supplementation on development of reproductive organs in Boer goat. Anim. Reprod. Sci. 2009, 113, 93–101. [Google Scholar] [CrossRef]
- Zhang, X.-G.; Wang, Y.-H.; Han, C.; Hu, S.; Wang, L.-Q.; Hu, J.-H. Effects of trehalose supplementation on cell viability and oxidative stress variables in frozen-thawed bovine calf testicular tissue. Cryobiology 2015, 70, 246–252. [Google Scholar] [CrossRef]
- Agarwal, A.; Durairajanayagam, D.; du Plessis, S.S. Utility of antioxidants during assisted reproductive techniques: An evidence based review. Reprod. Biol. Endocrinol. 2014, 12, 112. [Google Scholar] [CrossRef]
- Yamamoto, K.; Maruyama, K.; Himori, N.; Omodaka, K.; Yokoyama, Y.; Shiga, Y.; Morin, R.; Nakazawa, T. The novel Rho kinase (ROCK) inhibitor K-115: A new candidate drug for neuroprotective treatment in glaucoma. Investig. Ophthalmol. Vis. Sci. 2014, 55, 7126–7136. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, Y.; Maruyama, K.; Yamamoto, K.; Omodaka, K.; Yasuda, M.; Himori, N.; Ryu, M.; Nishiguchi, K.M.; Nakazawa, T. The role of calpain in an in vivo model of oxidative stress-induced retinal ganglion cell damage. Biochem. Biophys. Res. Commun. 2014, 451, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, Y.; Chen, H.; Dan, J.; Cheng, J.; Guo, S.; Sun, X.; Wang, W.; Ai, Y.; Li, S.; et al. The predominant pathway of apoptosis in THP-1 macrophage-derived foam cells induced by 5-aminolevulinic acid-mediated sonodynamic therapy is the mitochondria-caspase pathway despite the participation of endoplasmic reticulum stress. Cell. Physiol. Biochem. 2014, 33, 1789–1801. [Google Scholar] [CrossRef] [PubMed]
- DeLoughery, Z.; Luczak, M.W.; Zhitkovich, A. Monitoring Cr intermediates and reactive oxygen species with fluorescent probes during chromate reduction. Chem. Res. Toxicol. 2014, 27, 843–851. [Google Scholar] [CrossRef]
- Huang, Y.-Y.; Nagata, K.; Tedford, C.E.; McCarthy, T.; Hamblin, M.R. Low-level laser therapy (LLLT) reduces oxidative stress in primary cortical neurons in vitro. J. Biophotonics 2013, 6, 829–838. [Google Scholar] [CrossRef]
- Ahn, H.-Y.; Fairfull-Smith, K.E.; Morrow, B.J.; Lussini, V.; Kim, B.; Bondar, M.V.; Bottle, S.E.; Belfield, K.D. Two-photon fluorescence microscopy imaging of cellular oxidative stress using profluorescent nitroxides. J. Am. Chem. Soc. 2012, 134, 4721–4730. [Google Scholar] [CrossRef]
- Wood, J.W.; Bas, E.; Gupta, C.; Selman, Y.; Eshraghi, A.; Telischi, F.F.; Van De Water, T.R. Otoprotective properties of mannitol against gentamicin induced hair cell loss. Otol. Neurotol. 2014, 35, e187–e194. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Chatterjee, S.; Gagnon, C. Production of reactive oxygen species by spermatozoa undergoing cooling, freezing, and thawing. Mol. Reprod. Dev. 2001, 59, 451–458. [Google Scholar] [CrossRef]
- Lasso, J.L.; Noiles, E.E.; Alvarez, J.G.; Storey, B.T. Mechanism of superoxide dismutase loss from human sperm cells during cryopreservation. J. Androl. 1994, 15, 255–265. [Google Scholar]
- Price, P.; Goldsborough, M.; Tilkins, M. Embryonic Stem Cell Serum Replacement. WO98/30679, 16 July 1998. [Google Scholar]
- Beorlegui, N.; Cetica, P.; Trinchero, G.; Córdoba, M.; Beconi, M. Comparative study of functional and biochemical parameters in frozen bovine sperm. Andrologia 1997, 29, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Peña, F.J.; Johannisson, A.; Wallgren, M.; Rodriguez Martinez, H. Antioxidant supplementation in vitro improves boar sperm motility and mitochondrial membrane potential after cryopreservation of different fractions of the ejaculate. Anim. Reprod. Sci. 2003, 78, 85–98. [Google Scholar] [CrossRef]
- Breininger, E.; Beorlegui, N.B.; O’Flaherty, C.M.; Beconi, M.T. Alpha-tocopherol improves biochemical and dynamic parameters in cryopreserved boar semen. Theriogenology 2005, 63, 2126–2135. [Google Scholar] [CrossRef] [PubMed]
- Mulholland, C.A.; Benford, D.J. What is known about the safety of multivitamin-multimineral supplements for the generally healthy population? Theoretical basis for harm. Am. J. Clin. Nutr. 2007, 85, 318S–322S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pompella, A.; Visvikis, A.; Paolicchi, A.; De Tata, V.; Casini, A.F. The changing faces of glutathione, a cellular protagonist. Biochem. Pharmacol. 2003, 66, 1499–1503. [Google Scholar] [CrossRef]
- Johnson, F.C. The antioxidant vitamins. Crit. Rev. Food Sci. Nutr. 1979, 11, 217–309. [Google Scholar] [CrossRef]
- Yoganathan, T.; Eskild, W.; Hansson, V. Investigation of detoxification capacity of rat testicular germ cells and Sertoli cells. Free Radic. Biol. Med. 1989, 7, 355–359. [Google Scholar] [CrossRef]
- Takanami, Y.; Iwane, H.; Kawai, Y.; Shimomitsu, T. Vitamin E supplementation and endurance exercise: Are there benefits? Sports Med. 2000, 29, 73–83. [Google Scholar] [CrossRef]
- Oblette, A.; Rives, N.; Dumont, L.; Rives, A.; Verhaeghe, F.; Jumeau, F.; Rondanino, C. Assessment of sperm nuclear quality after in vitro maturation of fresh or frozen/thawed mouse pre-pubertal testes. Mol. Hum. Reprod. 2017, 23, 674–684. [Google Scholar] [CrossRef]
- Milazzo, J.P.; Vaudreuil, L.; Cauliez, B.; Gruel, E.; Massé, L.; Mousset-Siméon, N.; Macé, B.; Rives, N. Comparison of conditions for cryopreservation of testicular tissue from immature mice. Hum. Reprod. 2008, 23, 17–28. [Google Scholar] [CrossRef]
- Travers, A.; Arkoun, B.; Safsaf, A.; Milazzo, J.-P.; Absyte, A.; Bironneau, A.; Perdrix, A.; Sibert, L.; Macé, B.; Cauliez, B.; et al. Effects of vitamin A on in vitro maturation of pre-pubertal mouse spermatogonial stem cells. PLoS ONE 2013, 8, e82819. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | Testicular Tissue | Cryomedium | Post-Thaw Medium | Culture Medium |
|---|---|---|---|---|
| Fresh BM | Fresh | - | - | BM |
| Fresh Rol | Fresh | - | - | BM + Rol |
| F-T Rol | Frozen-thawed | No antioxidant | No antioxidant | BM + Rol |
| F-T Rol Vit E | Frozen-thawed | With Vit E | With Vit E | BM + Rol + Vit E |
| F-T Rol Vit E GSH | Frozen-thawed | With Vit E | With Vit E and GSH | BM + Rol + Vit E+GSH |
| F-T Rol GSH | Frozen-thawed | With GSH | With GSH | BM + Rol + GSH |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arkoun, B.; Galas, L.; Dumont, L.; Rives, A.; Saulnier, J.; Delessard, M.; Rondanino, C.; Rives, N. Vitamin E but Not GSH Decreases Reactive Oxygen Species Accumulation and Enhances Sperm Production during In Vitro Maturation of Frozen-Thawed Prepubertal Mouse Testicular Tissue. Int. J. Mol. Sci. 2019, 20, 5380. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215380
Arkoun B, Galas L, Dumont L, Rives A, Saulnier J, Delessard M, Rondanino C, Rives N. Vitamin E but Not GSH Decreases Reactive Oxygen Species Accumulation and Enhances Sperm Production during In Vitro Maturation of Frozen-Thawed Prepubertal Mouse Testicular Tissue. International Journal of Molecular Sciences. 2019; 20(21):5380. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215380
Chicago/Turabian StyleArkoun, Brahim, Ludovic Galas, Ludovic Dumont, Aurélie Rives, Justine Saulnier, Marion Delessard, Christine Rondanino, and Nathalie Rives. 2019. "Vitamin E but Not GSH Decreases Reactive Oxygen Species Accumulation and Enhances Sperm Production during In Vitro Maturation of Frozen-Thawed Prepubertal Mouse Testicular Tissue" International Journal of Molecular Sciences 20, no. 21: 5380. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20215380