Irrigation-Induced Changes in Chemical Composition and Quality of Seeds of Yellow Lupine (Lupinus luteus L.)

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Seed Yield
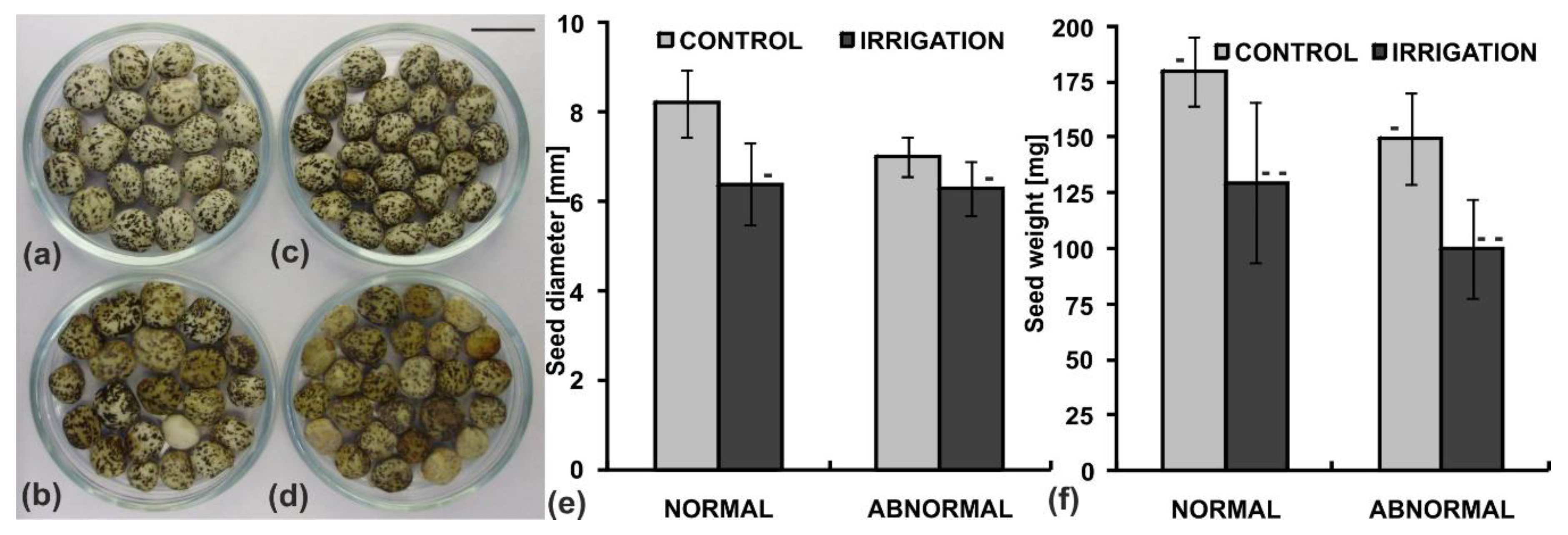
2.2. Seed Morphology
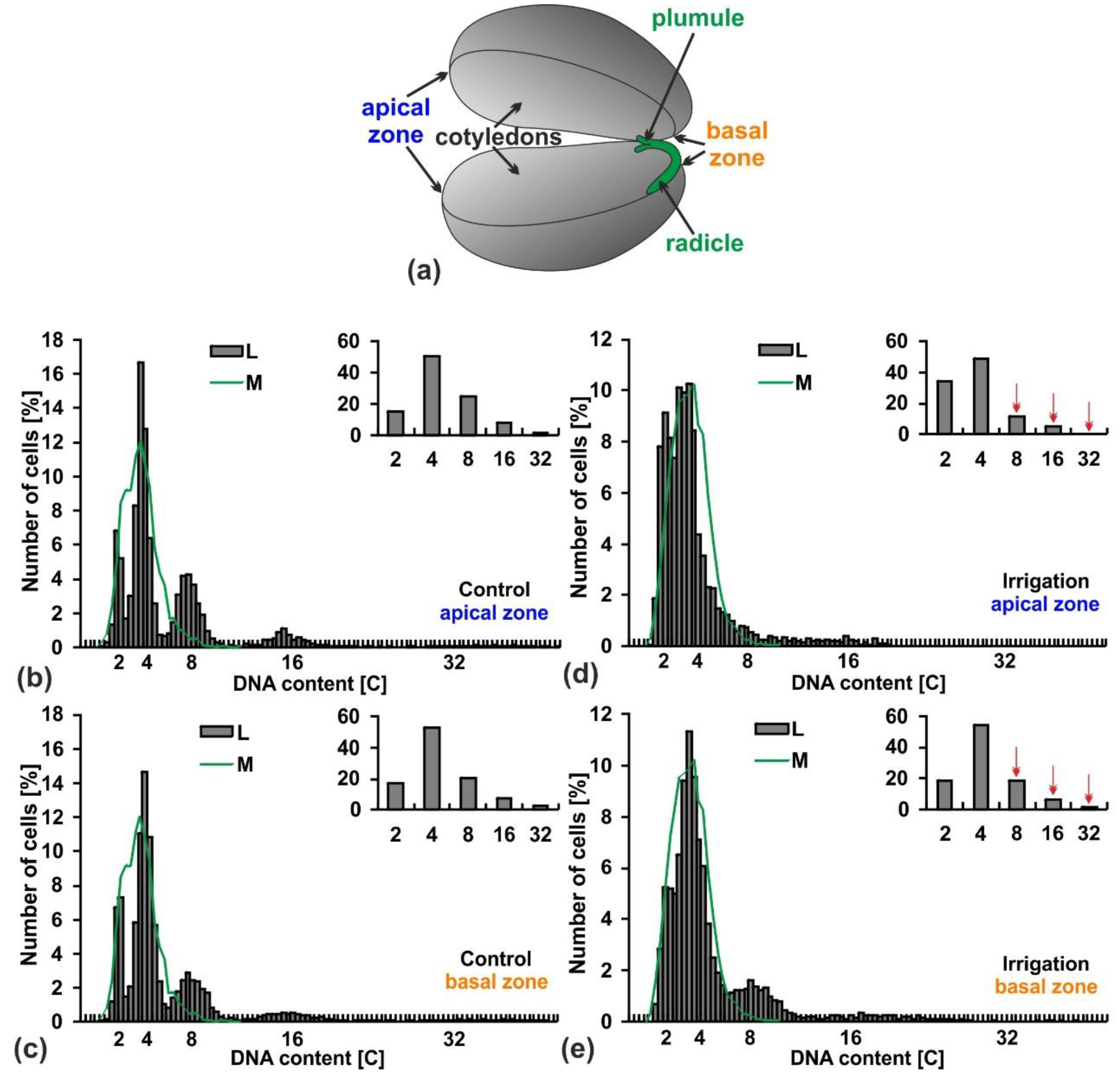
2.3. DNA Content
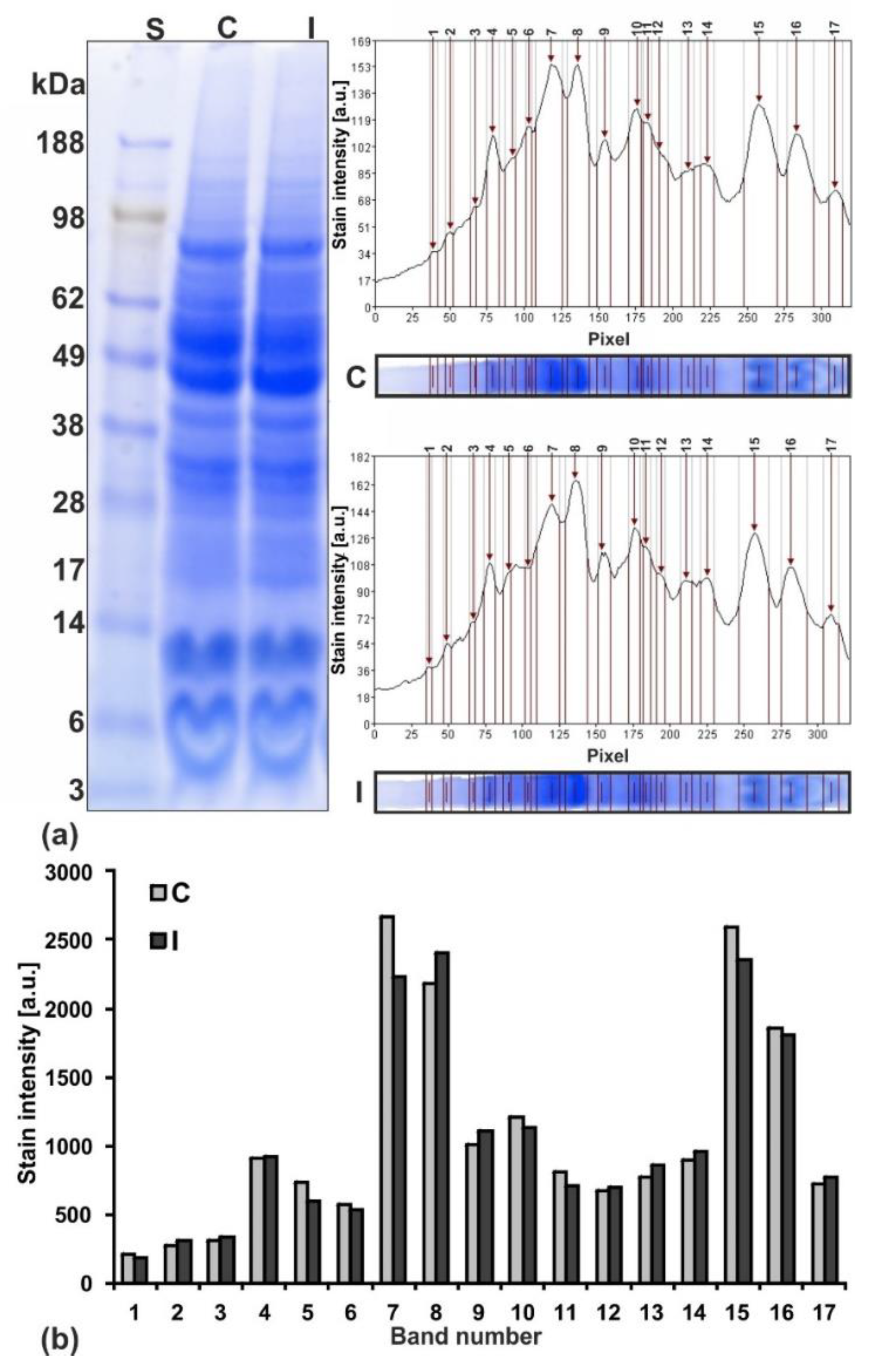
2.4. Protein Profiles
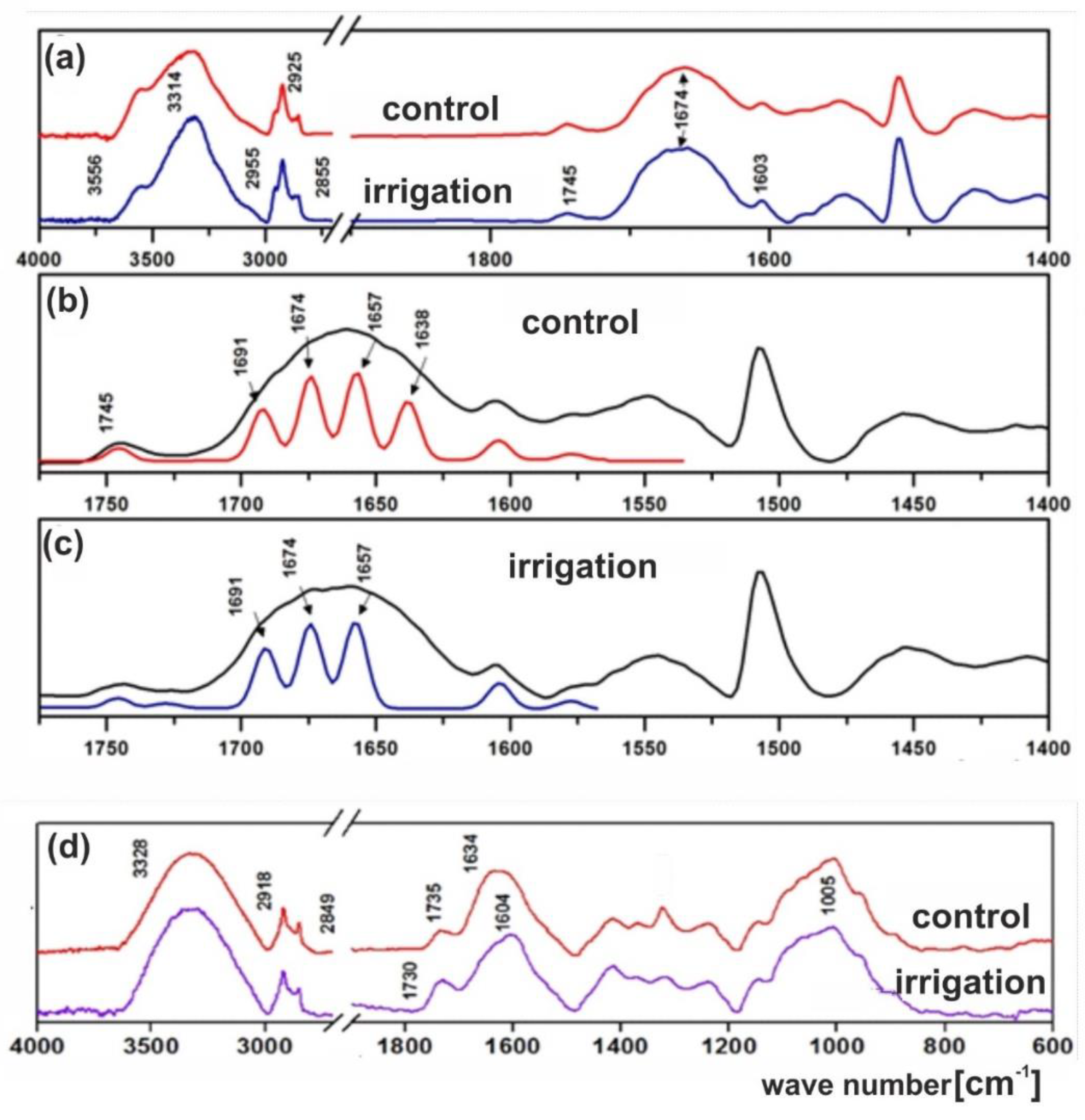
2.5. Fourier Transform–Infrared Spectroscopy (FT-IR) Analysis of Lupine Seeds
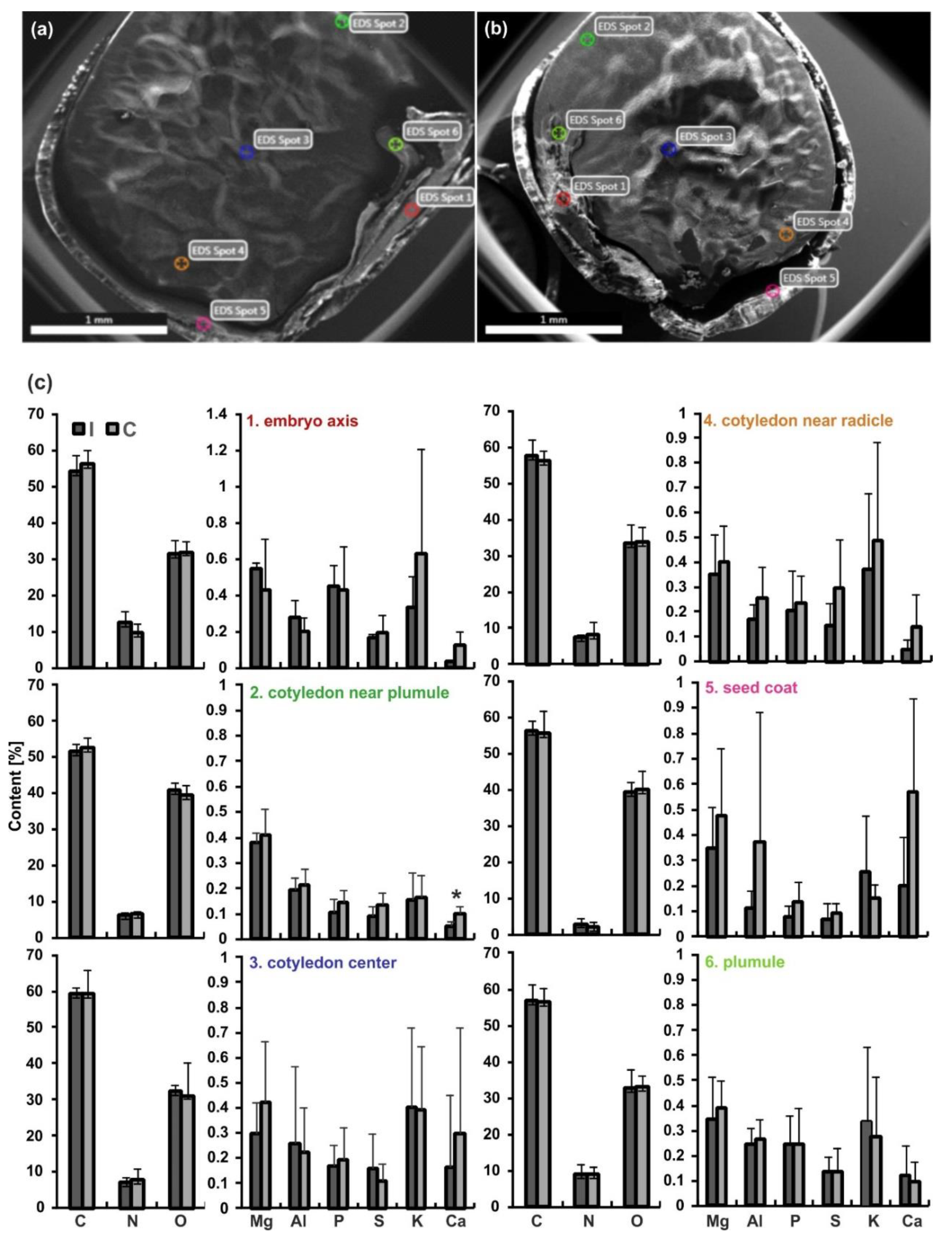
2.6. Analysis of Chemical Elements by the SEM/Energy Dispersive Spectroscopy (EDS) Technique
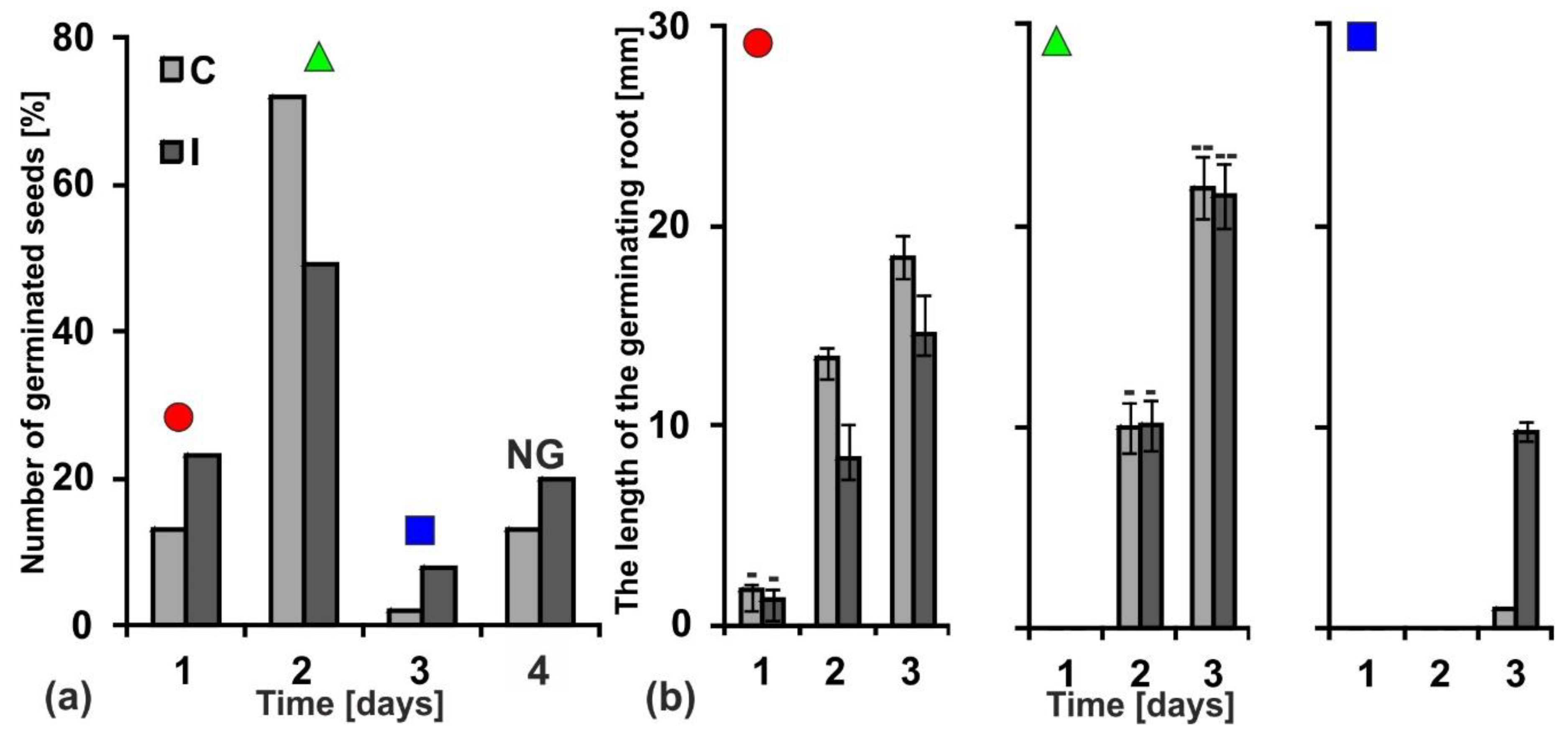
2.7. Germination and Seedlings’ Growth
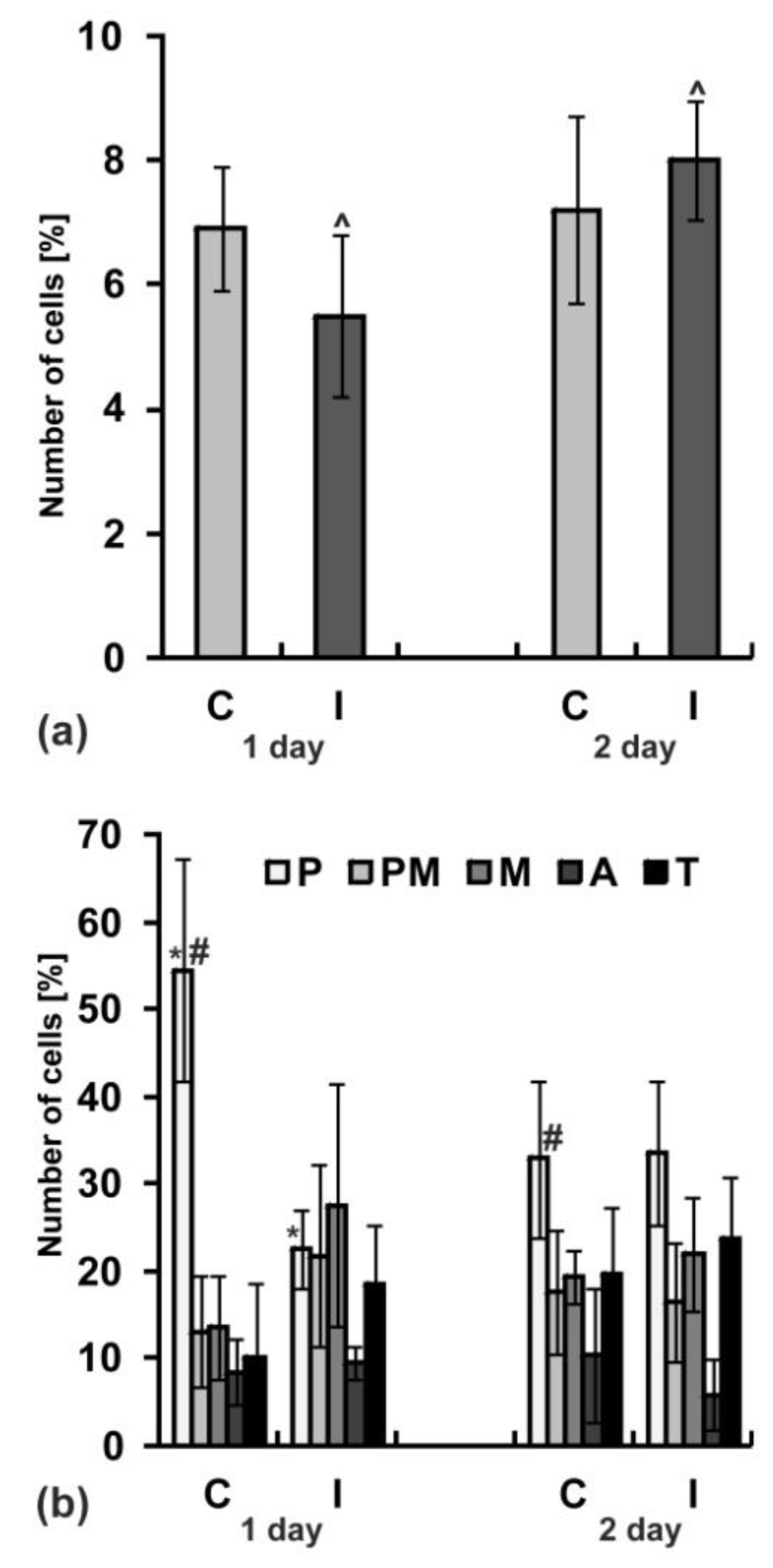
2.8. Mitotic Activity
2.9. Detection of Hydrogen Peroxide
3. Discussion
4. Materials and Methods
4.1. Plant Cultivation
4.2. Yield Assessment
4.3. Seed Germination for Cytological Study
4.4. Cytophotometry
4.5. Electrophoretic Separation of Proteins
4.6. FTIR Analysis of Lupine Seeds
4.7. SEM/EDS Microanalysis
4.8. Histochemical Localization of H2O2
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FTIR | Fourier transform infrared spectroscopy |
| SEM | Scanning electron microscope |
| EDS | Energy dispersive spectroscopy |
References
- Sujak, A.; Kotlarz, A.; Strobel, W. Compositional and nutritional evaluation of several lupin seeds. Food Chem. 2006, 98, 711–719. [Google Scholar] [CrossRef]
- Fernández-Pascual, M.; PueyoMaría, J.J.; Lucas, M.M. Singular features of the Bradyrhizobium-Lupinus symbiosis. Dyn. Soil Dyn. Plant 2007, 1, 1–16. [Google Scholar]
- Coba de la Peña, Y.; Pueyo, J.J. Legumes in the reclamation of marginal soils, from cultivar and inoculants selection to transgenic approaches. Agron. Sustain. Dev. 2012, 32, 65–91. [Google Scholar] [CrossRef]
- Lucas, M.M.; Stoddard, F.L.; Annicchiarico, P.; Frías, J.; Martínez-Villaluenga, C.; Sussmann, D.; Duranti, M.; Seger, A.; Zander, P.M.; Pueyo, J.J. The future of lupin as a protein crop in Europe. Front. Plant Sci. 2015, 6, 705. [Google Scholar] [CrossRef] [PubMed]
- Wilmowicz, E.; Kućko, A.; Burchardt, S.; Przywieczerski, T. Molecular and hormonal aspects of drought-triggered flower shedding in yellow lupine. Int. J Mol. Sci. 2019, 20, 3731. [Google Scholar] [CrossRef]
- Graham, P.H.; Vance, C.P. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef]
- Straková, E.; Suchý, P.; Večerek, V.; Šerman, V.; Mas, N.; Jůzl, M. Nutritional composition of seeds of genus Lupinus. Acta Vet. Brno 2006, 75, 489–493. [Google Scholar] [CrossRef]
- Ogura, T.; Ogihara, J.; Sunairi, M.; Takeishi, H.; Aizawa, T.; Olivos-Trujillo, M.R.; Maureira-Butler, I.J.; Salvo-Garrido, H.E. Proteomic characterization of seeds from yellow lupin (Lupinus luteus L.). Proteomics 2014, 14, 1543–1546. [Google Scholar] [CrossRef]
- Juzoń, K.; Czyczyło-Mysza, I.; Marcińska, I.; Dziurka, M.; Waligórski, P.; Skrzypek, E. Polyamines in yellow lupin (Lupinus luteus L.) tolerance to soil drought. Acta Physiol. Plant. 2017, 39, 202. [Google Scholar] [CrossRef]
- Bettzieche, A.; Brandsch, C.; Eder, K.; Stangl, G.I. Lupin protein acts hypocholesterolemic and increases milk fat content in lactating rats by influencing the expression of genes involved in cholesterol homeostasis and triglyceride synthesis. Mol. Nutr. Food Res. 2009, 53, 1134–1142. [Google Scholar] [CrossRef]
- Duranti, M.; Morazzoni, P. Nutraceutical properties of lupin seed proteins. A great potential still waiting for full exploitation. Agro Food Ind. Hi Technol. 2011, 22, 20–23. [Google Scholar]
- Walsh, C.J.; Guinane, C.M.; O’Toole, P.W.; Cotter, P.D. Beneficial modulation of the gut microbiota. FEBS Lett. 2014, 588, 4120–4130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strozycki, P.M.; Szczurek, A.; Lotocka, B.; Figlerowicz, M.; Legocki, A.B. Ferritins and nodulation in Lupinus luteus: Iron management in indeterminate type nodules. J. Exp. Bot. 2007, 58, 3145–3153. [Google Scholar] [CrossRef] [PubMed]
- Bieniaszewski, T.; Szwejkowski, Z.; Fordoński, G. Impact of temperature and rainfall distribution over 1989-1996 on the biometric and structural characteristics as well as on the ‘Juno’ yellow lupin yielding. EJPAU 2000, 3. Available online: http://www.ejpau.media.pl/volume3/issue2/agronomy/art-02.html (accessed on 5 November 2019).
- Faligowska, A.; Szukała, J. Influence of irrigation and soil tillage systems on vigour and sowing value of yellow lupine seeds. Sci. Nat. Technol. 2012, 2, 26. [Google Scholar]
- Podleśny, J.; Podleśna, A. Wpływ wysokiej temperatury w okresie kwitnienia na wzrost, rozwój i plonowanie łubinu żółtego. Acta Agrophyscia 2012, 19, 825–834. [Google Scholar]
- Pszczółkowska, A.; Olszewski, J.; Płodzień, K.; Kulik, T.; Fordoński, G.; Żuk-Gołaszwska, K. Effect of the water stress on the productivity of selected genotypes of pea (Pisum sativum L.) and yellow lupin (Lupinus luteus L.). Electron. J. Polish Agric. Univ, Agron. 2003, 6, 1. [Google Scholar]
- Gresta, F.; Wink, M.; Prins, U.; Abberton, M.; Capraro, J.; Scarafoni, A.; Hill, G. Lupins in european cropping system. In Legumes in Cropping System; Murphy-Bokern, D., Stoddard, F.L., Watson, C.A., Eds.; CABI Publishing: Wallingford, UK, 2017; pp. 88–108. [Google Scholar]
- Okamoto, K.; Sagata, N. Mechanism for inactivation of the mitotic inhibitory kinase Wee1 at M phase. Proc. Nat. Acad. Sci. USA 2007, 104, 3753–3758. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Basra, A.S.M. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Lisar, S.Y.S.; Motafakkerazad, R.; Hossain, M.M.; Rahman, I.M.M. Water Stress in Plants: Causes, Effects and Responses. In Water Stress; Rahman, I.M.M., Ed.; InTech: London, UK, 2012; pp. 1–16. [Google Scholar]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S.P. Response of plants to water stress. Front. Plant. Sci. 2014, 5, 1–8. [Google Scholar] [CrossRef]
- Shanker, A.K.; Maheswari, M.; Yadav, S.K.; Desai, S.; Bhanu, D.; Attal, N.B.; Venkateswarlu, B. Drought stress responses in crops. Funct. Integr. Genomics 2014, 14, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Sharma, K.D. Molecular and Morphophysiological Analysis of Drought Stress in Plants Summy. In Plant Growth; Rigobello, E., Ed.; InTech: London, UK, 2016; pp. 149–173. [Google Scholar]
- Kang, Y.; Khan, S.; Ma, X. Climate change impacts on crop yield, crop water productivity and food security - A review. Prog. Nat. Sci. 2009, 19, 1665–1674. [Google Scholar] [CrossRef]
- Morison, J.I.; Baker, N.; Mullineaux, P.; Davies, W. Improving water use in crop production. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 639–658. [Google Scholar] [CrossRef] [PubMed]
- Almer, C.; Laurent-Lucchetti, J.; Oechslin, M. Water scarcity and rioting: Disaggregated evidence from Sub-Saharan Africa. J. Environ. Econ. Manage. 2016, 86, 193–209. [Google Scholar] [CrossRef]
- Cox, F.R.; Sullivan, G.A.; Martin, C.K. Effect of calcium and irrigation treatments on peanut yield, grade and seed quality 1. Peanut Sci. 1976, 3, 81–85. [Google Scholar] [CrossRef]
- Holden, N.M.; Brereton, A.J. Adaptation of water and nitrogen management of spring barley and potato as a response to possible climate change in Ireland. Agric. Water Manag. 2006, 82, 297–317. [Google Scholar] [CrossRef]
- Zhao, H.L.; Cui, J.Y.; Zhou, R.L.; Zhang, T.H.; Zhao, X.Y.; Drake, S. Soil properties, crop productivity and irrigation effects on five croplands of Inner Mongolia. Soil Tillage Res. 2007, 93, 346–355. [Google Scholar] [CrossRef]
- Kassab, O.M.; Abo Ellil, A.A.; Abdallah, E.F.; Ibrahim, M.M. Performance of some chickpea cultivars under sprinkler irrigation treatments in sandy soil. Aust. J. Basic App. Sci. 2012, 6, 618–625. [Google Scholar]
- Levidow, L.; Zaccaria, D.; Maia, R.; Vivas, E.; Todorovic, M.; Scardigno, A. Improving water-efficient irrigation: Prospects and difficulties of innovative practices. Agric. Water Manag. 2014, 146, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Breen, A.N.; Richards, J.H. Irrigation and fertilization effects on seed number, size, germination and seedling growth: Implications for desert shrub establishment. Oecologia 2008, 157, 13–19. [Google Scholar] [CrossRef]
- Faligowska, A.; Panasiewicz, K.; Szymańska, G.; Bartos-Spychała, M. The seeds quality of yellow lupine depending on selected agrotechnical factors. Prog. Plant Prot. 2013, 53, 293–296. [Google Scholar] [Green Version]
- Faligowska, A.; Panasiewicz, K.; Szukała, J.; Koziara, W. Germination and vigour of narrow-leaved lupin seeds as the effect of irrigation of parent plants and cultivation in different soil tillage systems. Polish J. Agron. 2016, 24, 3–8. [Google Scholar]
- Tooke, P.B. Fourier self-deconvolution in IR spectroscopy. Trends Analyt. Chem. 1988, 7, 130–136. [Google Scholar] [CrossRef]
- Lamba, O.P.; Borchman, D.; Sinha, S.K.; Shah, J.; Renugopalakrishnan, V.; Yappert, M.C. Estimation of the secondary structure and conformation of bovine lens crystallins by infrared spectroscopy: Quantitative analysis and resolution by Fourier self-deconvolution and curve fit. Biochim. Biophys. Acta 1993, 1163, 113–123. [Google Scholar] [CrossRef]
- Ribeiro da Luz, B. Attenuated total reflectance spectroscopy of plant leaves: A tool for ecological and botanical studies. New Phytol. 2006, 172, 305–318. [Google Scholar] [CrossRef]
- Żarski, J.; Dudek, S.; Kuśmierek-Tomaszewska, R.; Rolbiecki, R.; Rolbiecki, S. Prognozowanie efektów nawadniania roślin na podstawie wybranych wskaźników suszy meteorologicznej i rolniczej. Rocznik Ochr. Środ. 2013, 15, 2185–2203. [Google Scholar]
- Davies, C.L.; Turner, D.W.; Dracup, M. Yellow lupin (Lupinus luteus) tolerates waterlogging better than narrow-leaved lupin (L. angustifolius). I. Schoot and root growth in a controlled environment. Aust. J. Agric. Res. 2000, 51, 701–709. [Google Scholar] [CrossRef]
- Dracup, M.; Turner, N.C.; Tang, C.; Reader, M.; Palta, J. Responses to abiotic stresses. Chapter 8. In Lupins as Crop Plants: Biology, Production and Utilization; Gladstones, J.S., Atkins, C.A., Hamblin, J., Eds.; CABI: Wallingford, UK, 1998; pp. 227–262. [Google Scholar]
- Płażek, A.; Dubert, F.; Kopeć, P.; Dziurka, M.; Kalandyk, A.; Pastuszak, J.; Wolko, B. Seed hydropriming and smoke water significantly improve low-temperature germination of Lupinus angustifolius L. Int. J. Mol. Sci. 2018, 19, 992. [Google Scholar] [CrossRef]
- Winnicki, K.; Ciereszko, I.; Leśniewska, J.; Dubis, A.T.; Basa, A.; Żabka, A.; Hołota, M.; Sobiech, Ł.; Faligowska, A.; Skrzypczak, G.; et al. Irrigation affects characteristics of Barrow-leaved lupin (Lupinus angustifolius L.). Planta 2019, 249, 1731–1746. [Google Scholar] [CrossRef]
- Ambika, S.; Manonmani, V.; Somasundaram, G. Review on effect of seed size on seedling vigour and seed yield. Res. J. Seed Sci. 2014, 7, 31–38. [Google Scholar] [CrossRef]
- Peksen, E.; Peksen, A.; Bozoglu, H.; Gulumser, A. Some seed traits and their relationships to seed germination and field emergence in pea (Pisum sativum L.). J. Agron. 2004, 3, 243–246. [Google Scholar]
- Rastegar, Z.; Kandi, M.A.S. The effect of salinity and seed size on seed reserve utilization and seedling growth of soybean (Glycin max). Int. J. Agron. Plant Prod. 2011, 2, 1–4. [Google Scholar]
- Farhoudi, R.; Motamedi, M. Effect of salt stress and seed size on germination and early seedling growth of safflower (Carthamus tinctorius L.). Seed Sci. Technol. 2010, 38, 73–78. [Google Scholar] [CrossRef]
- Bourdon, M.; Pirrello, J.; Cheniclet, C.; Coriton, O.; Bourge, M.; Brown, S.; Moïse, A.; Peypelut, M.; Rouyère, V.; Renaudin, J.P.; et al. Evidence for karyoplasmic homeostasis during endoreduplication and a ploidy-dependent increase in gene transcription during tomato fruit growth. Development 2012, 139, 3817–3826. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Umeda, M. Cytokinins promote onset of endoreplication by controlling cell cycle machinery. Plant Signal Behav. 2014, 9, e29396. [Google Scholar] [CrossRef] [Green Version]
- Scholes, D.R.; Paige, K.N. Plasticity in ploidy: A generalized response to stress. Trends Plant Sci. 2015, 20, 165–175. [Google Scholar] [CrossRef]
- Lee, H.O.; Davidson, J.M.; Duronio, R.J. Endoreplication: Polyploidy with purpose. Genes Develop. 2009, 23, 2461–2477. [Google Scholar] [CrossRef]
- Knake-Sobkowicz, S.; Marciniak, K. Cellular accumulation of protein bodies and changes in DNA ploidy level during seed development of Lathyrus tuberosus L. Acta Biol. Crac. Series Bot. 2005, 47, 147–157. [Google Scholar]
- Dante, R.A.; Larkins, B.A.; Sabelli, P.A. Cell cycle control and seed development. Front. Plant. Sci. 2014, 5, 1–14. [Google Scholar] [CrossRef]
- Leiva-Neto, J.T.; Grafi, G.; Sabelli, P.A.; Dante, R.A.; Woo, Y.; Maddock, S.; Gordon-Kamm, W.J.; Larkins, B.A. A dominant negative mutant of cyclin-dependent kinase A reduces endoreduplication but not cell size or gene expression in maize endosperm. Plant Cell 2004, 16, 1854–1869. [Google Scholar] [CrossRef]
- Joubès, J.; Chevalier, C. Endoredupication in higher plants. Plant Mol. Biol. 2000, 43, 735–745. [Google Scholar] [CrossRef] [PubMed]
- González-Sama, A.; Coba de la Peña, T.; Kevei, Z.; Mergaert, P.; Lucas, M.M.; de Felipe, M.R.; Kondorosi, E.; Pueyo, J.J. Nuclear DNA endoreduplication and expression of the mitotic inhibitor Ccs52 associated to determinate and lupinoid nodule organogenesis. Mol. Plant-Microbe Inter. J. 2006, 19, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Yeung, E.C.; Paek, K. Endoredupication in Phalaenopsis is affected by light quality from light-emitting diodes during somatic embryogenesis. Plant Biotechnol. Rep. 2010, 4, 303–309. [Google Scholar] [CrossRef]
- Chevalier, C.; Nafati, M.; Mathieu-Rivet, E.; Bourdon, M.; Frangne, N.; Cheniclet, C.; Renauldin, J.-P.; Gévaudant, F.; Hernould, M. Elucidating the functional role of endoreduplication in tomato fruit development. Ann. Bot. 2011, 107, 1159–1169. [Google Scholar] [CrossRef] [Green Version]
- Kondorosi, E.; Roudier, F.; Gendreau, E. Plant cell size control: Growing by ploidy? Curr. Opin. Plant Biol. 2000, 3, 488–492. [Google Scholar] [CrossRef]
- Breuer, C.; Braidwood, L.; Sugimoto, K. Endocycling in the path of plant development. Curr. Opin. Plant Biol. 2014, 17, 78–85. [Google Scholar] [CrossRef]
- Lqari, H.; Pedroche, J.; Girón-Calle, J.; Vioque, J.; Millán, F. Purification and partial characterization of storage proteins in Lupinus angustifolius seeds. Grasas y Aceites 2004, 55, 364–369. [Google Scholar]
- Duranti, M.; Consonni, A.; Magni, C.; Sessa, F.; Scarafoni, A. The major proteins of lupin seed: Characterisation and molecular properties for use as functional and nutraceutical ingredients. Trends Food Sci. Technol. 2008, 19, 624–633. [Google Scholar] [CrossRef]
- Foley, R.C.; Jimenez-Lopez, C.; Kamphuis, L.G.; Hane, J.K.; Melser, S.; Singh, K.B. Analysis of conglutin seed storage proteins across lupin species using transcriptomic, protein and comparative genomic approaches. BMC Plant Biol. 2015, 15, 106. [Google Scholar] [CrossRef]
- Jimenez-Lopez, J.C.; Melser, S.; DeBoer, K.; Thatcher, L.F.; Kamphuis, L.G.; Foley, R.C.; Singh, K.B. Narrow-leaved lupin (Lupinus angustifolius) β1- and β6-conglutin proteins exhibit antifungal activity, protecting plants against necrotrophic pathogen induced damage from Sclerotinia sclerotiorum and Phytophthora nicotianae. Front. Plant. Sci. 2016, 7, 1856. [Google Scholar] [CrossRef]
- Barciszewska-Pacak, M.; Milanowska, K.; Knop, K.; Bielewicz, D.; Nuc, P.; Plewka, P.; Pacak, A.M.; Vazquez, F.; Karłowski, W.; Jarmołowski, A.; et al. Arabidopsis microRNA expression regulation in a wide range of abiotic stress responses. Front Plant Sci. 2015, 6, 410. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, M.; Covarrubis, A.A. Late Embryogenesis Abundant (LEA) proteins in legumes. Front. Plant Sci. 2013, 4, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi. J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Heredia, A. Biophysical and biochemical characteristics of cutin, a plant barrier biopolymer. Bioch. Bioph. Acta 2003, 1620, 1–7. [Google Scholar] [CrossRef]
- Shao, S.; Meyer, C.J.; Ma, F.; Peterson, C.A.; Bernards, M.A. The outermost cuticle of soybean seeds: Chemical composition and function during imbibition. J. Exp. Bot. 2007, 58, 1071–1082. [Google Scholar] [CrossRef]
- Clua, A.A.; Gimenez, D.O. Environmental factors during seed development of narrow-leaved bird’s-foot-trefoil (Lotus tenuis) influences subsequent dormancy and germination. Grass Forage Sci. 2003, 58, 333–338. [Google Scholar] [CrossRef]
- Clua, A.; Fernandez, G.; Ferro, L.; Dietrich, M. Drought stress conditions during seed development of narrowleaf birdsfoot trefoil (Lotus glaber) influences seed production and subsequent dormancy and germination. Lotus Newsl. 2006, 36, 58–63. [Google Scholar]
- Ranal, M.A.; Santana, D.G. How and why to measure the germination process? Rev. Bras. Bot. 2006, 29, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, K.; Job, C.; Groot, S.P.C.; Puype, M.; Demol, H.; Vandekerckhove, J.; Job, D. Proteomic analysis of Arabidopsis seed germination and priming. Plant Physiol. 2001, 126, 835–848. [Google Scholar] [CrossRef]
- Osuna, D.; Prieto, P.; Aguilar, M. Control of seed germination and plant development by carbon and nitrogen availability. Front. Plant Sci. 2015, 6, 1023. [Google Scholar] [CrossRef]
- Tuteja, N.; Mahajan, S. Calcium signaling network in plants. Plant Signal Behav. 2007, 2, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Polit, J.T.; Ciereszko, I. In situ activities of hexokinase and fructokinase in relation tophosphorylation status of root meristem cells of Vicia faba during reactivation from sugar starvation. Physiol. Plant. 2009, 135, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Polit, J.T.; Ciereszko, I. Sucrose synthase activity and carbohydrates content in relation to phosphorylation status of Vicia faba root meristems during reactivation from sugar depletion. J. Plant Physiol. 2012, 169, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Kurusu, T.; Kimura, S.; Tada, Y.; Kaya, H.; Kuchitsu, K. Plant signaling networks involving reactive oxygen species and Ca2+. In Handbook on reactive oxygen species (ROS): Formation mechanisms, physiological roles and common harmful effects; Suzuki, M., Yamamoto, S., Eds.; Nova Science Publishers: Hauppauge, NY, USA, 2013; pp. 315–324. [Google Scholar]
- Passardi, F.; Penel, C.; Dunand, C. Performing the paradoxical: How plant peroxidases modify the cell wall. Trends Plant Sci. 2004, 9, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Dunand, C.; Crèvecoeur, M.; Penel, C. Distribution of superoxide and hydrogen peroxide in Arabidopsis root and their influence on root development: Possible interaction with peroxidases. New Physiol. 2007, 174, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xing, D.; Li, L.; Zhang, L. Rapid determination of seed vigor based on the level of superoxide generation during early imbibitions. Photochem. Photobiol. Sci. 2007, 6, 767–774. [Google Scholar] [CrossRef]
- Szopińska, D. Effects of hydrogen peroxide treatment on the germination, vigour and health of Zinnia elegans seeds. Folia Hort. 2014, 26, 19–29. [Google Scholar] [CrossRef]
- Maszewski, J.; Kaźmierczak, A.; Polit, J. Cell cycle agrest in antheridial extract-treated Root meristems of Allium cepa and Melandrium noctiflorum. Folia Histochem. Cytobiol. 1998, 36, 35–43. [Google Scholar]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin Phenol Reagen. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Dubis, E.N.; Dubis, A.T.; Popławski, J. Determination of the aromatic compounds in plant cuticular waxes using FT-IR spectroscopy. J. Mol. Struct. 2001, 596, 83–88. [Google Scholar] [CrossRef]
- He, H.; Veneklaas, E.J.; Kuo, J.; Lambers, H. Physiological and ecological significance of biomineralization in plants. Trends Plant Sci. 2014, 19, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Psaras, G.K.; Manetas, Y. Nickel localization in seeds of the metal hyperaccumulator Thlaspi pindicum Hausskn. Ann. Bot. 2001, 88, 513–516. [Google Scholar] [CrossRef]
- Thordal-Christensen, H.; Zhang, Z.; Wei, Y.; Collinge, D.B. Subcellular localization of H2O2 in plants: H2O2 accumulation in papillae and hypersensitive response during the barley-powdery mildew interaction. Plant J. 1997, 11, 1187–1194. [Google Scholar] [CrossRef]
- Żabka, A.; Polit, J.T.; Maszewski, J. DNA replication stress induces deregulation of the cell cycle events in root meristems of Allium cepa. Ann. Bot. 2012, 110, 1581–1591. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Stem | Branches | Plant | |
|---|---|---|---|
| Control | 3.68 | 0.38 | 4.06 |
| Irrigation | 4.14 | 0.94 | 5.08 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polit, J.T.; Ciereszko, I.; Dubis, A.T.; Leśniewska, J.; Basa, A.; Winnicki, K.; Żabka, A.; Audzei, M.; Sobiech, Ł.; Faligowska, A.; et al. Irrigation-Induced Changes in Chemical Composition and Quality of Seeds of Yellow Lupine (Lupinus luteus L.). Int. J. Mol. Sci. 2019, 20, 5521. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225521
Polit JT, Ciereszko I, Dubis AT, Leśniewska J, Basa A, Winnicki K, Żabka A, Audzei M, Sobiech Ł, Faligowska A, et al. Irrigation-Induced Changes in Chemical Composition and Quality of Seeds of Yellow Lupine (Lupinus luteus L.). International Journal of Molecular Sciences. 2019; 20(22):5521. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225521
Chicago/Turabian StylePolit, Justyna T., Iwona Ciereszko, Alina T. Dubis, Joanna Leśniewska, Anna Basa, Konrad Winnicki, Aneta Żabka, Marharyta Audzei, Łukasz Sobiech, Agnieszka Faligowska, and et al. 2019. "Irrigation-Induced Changes in Chemical Composition and Quality of Seeds of Yellow Lupine (Lupinus luteus L.)" International Journal of Molecular Sciences 20, no. 22: 5521. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225521