Alpha-Lipoic Acid Ameliorates Radiation-Induced Lacrimal Gland Injury through NFAT5-Dependent Signaling

 , , , , and
, , , , and 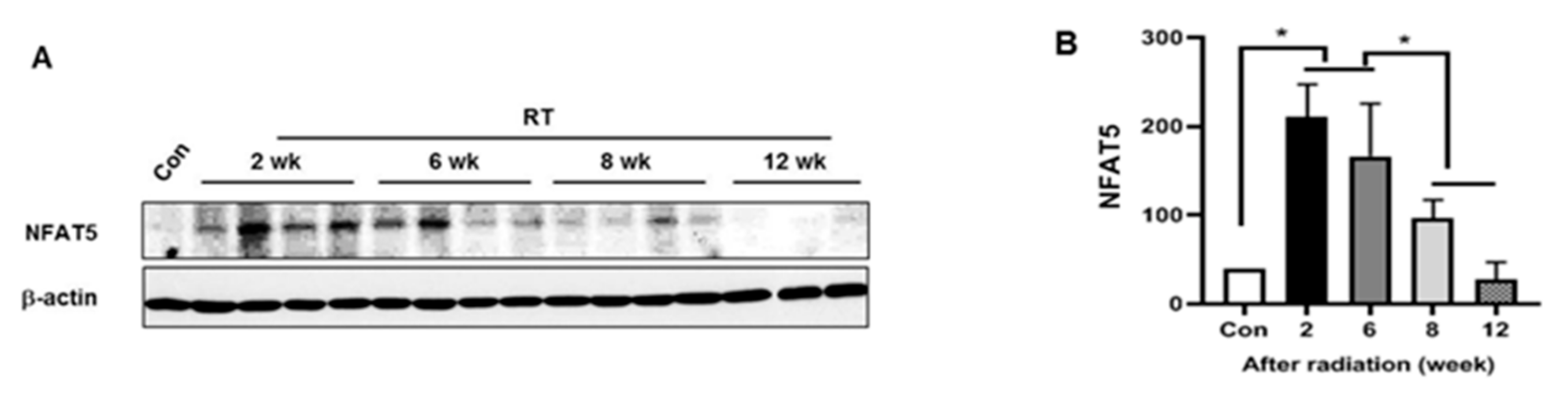{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. NFAT5 Expression during the LG Injury Progression to Radiation
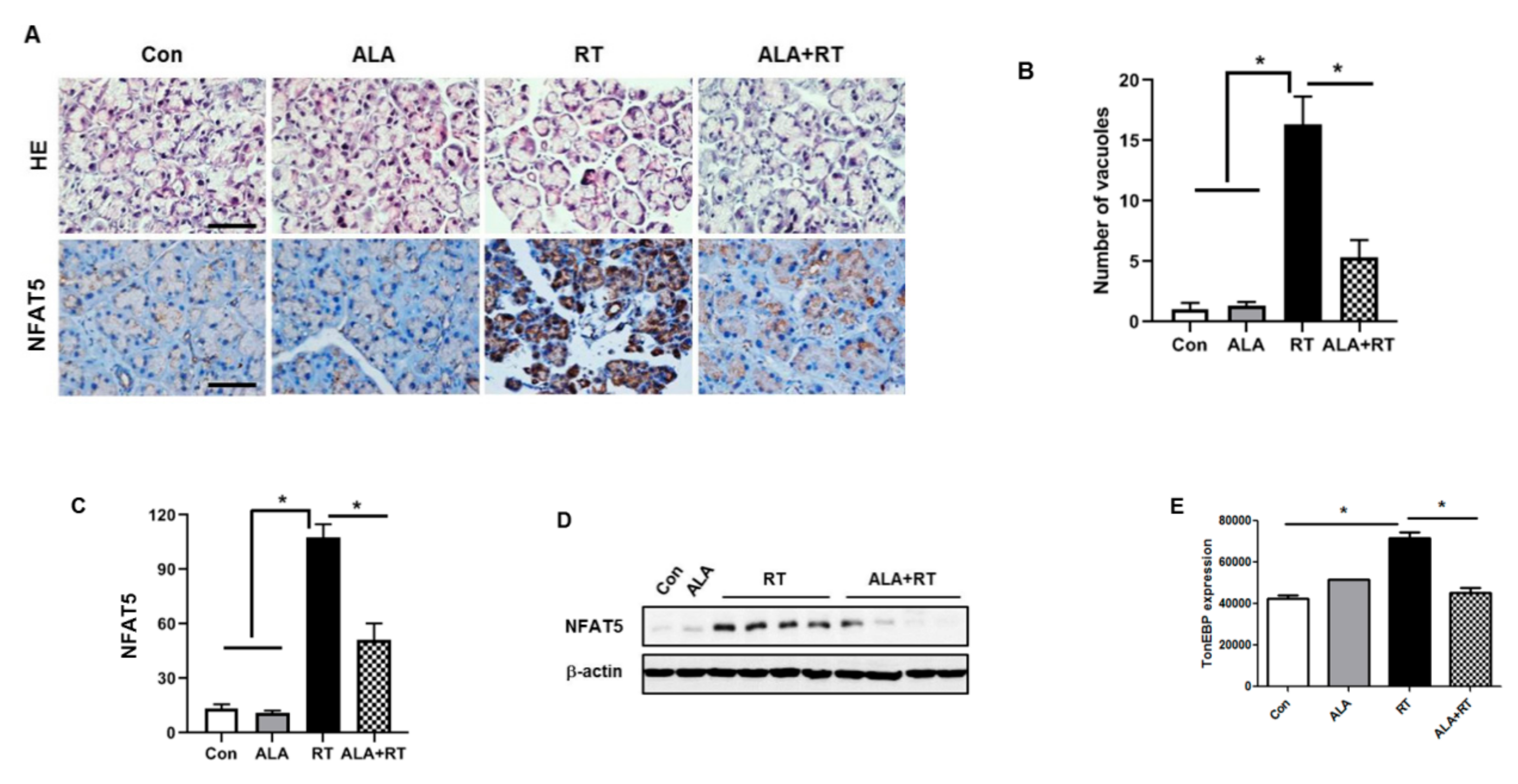
2.2. Effect of ALA on Radiation-Induced NFAT5 Expression in the LG
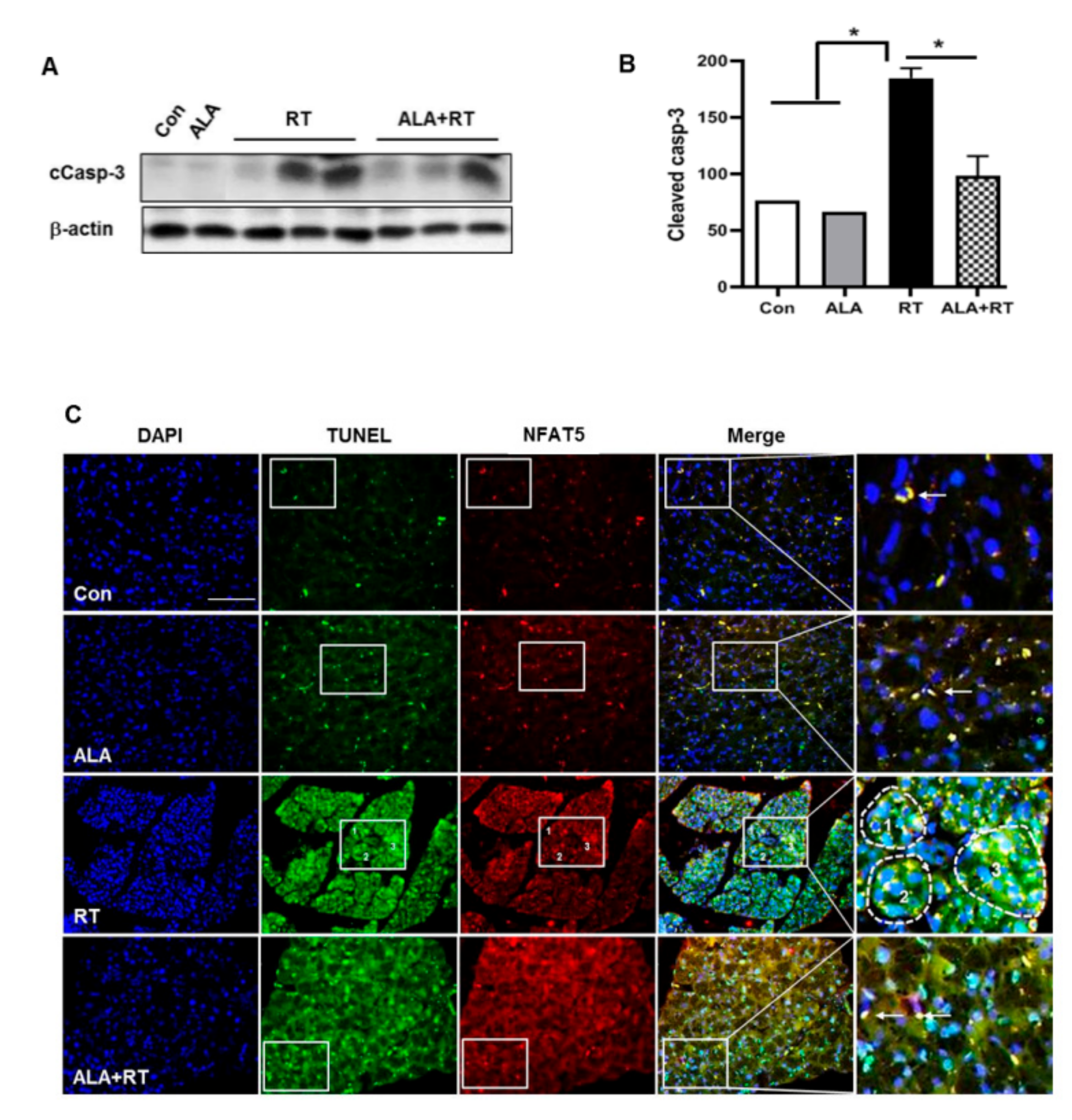
2.3. Effect of ALA on Radiation-Induced Apoptosis in the LG
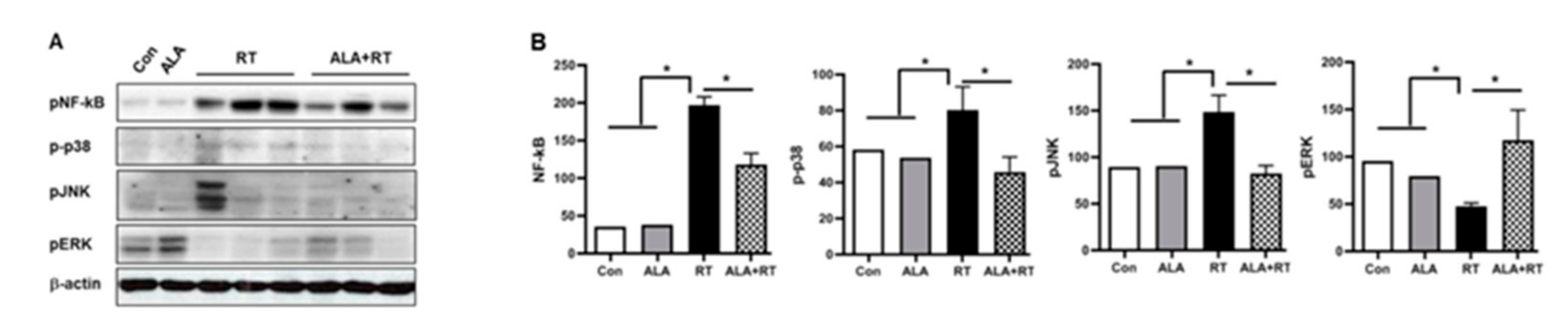
2.4. Suppression of NFAT5-Dependent Signaling Pathway by ALA Treatment in the LG
2.5. Protective Effects of ALA in Radiation-Induced LG Injury
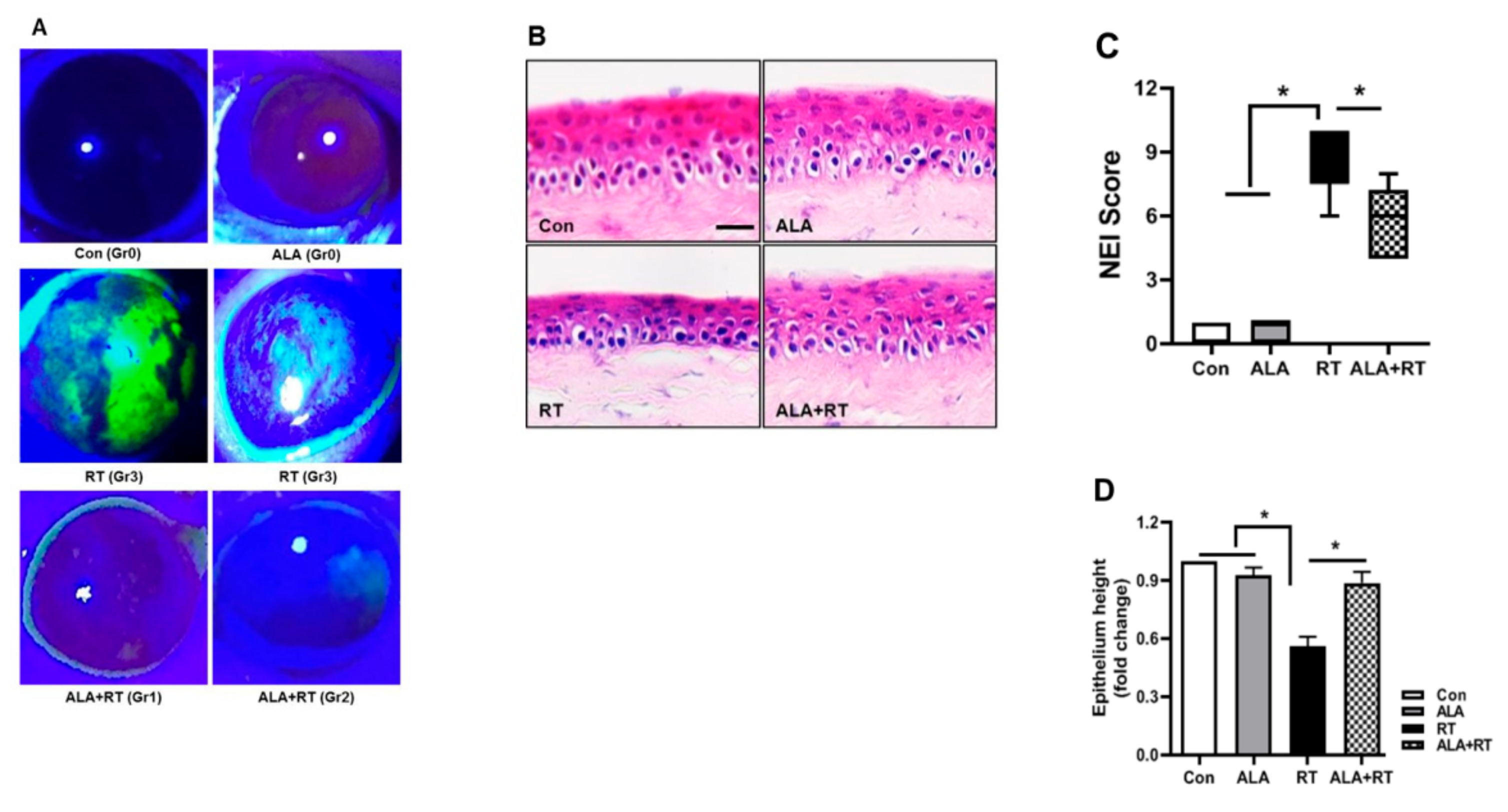
2.6. Effect of ALA on Corneal Fluorescein Staining and Integrity of the Corneal Epithelium
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Radiation Exposure
4.3. Immunoblotting
4.4. Double Immunofluorescence Staining
4.5. Immunohistochemistry
4.6. Corneal Fluorescein Staining
4.7. Histology for Corneal Epithelium
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Guchelaar, H.J.; Vermes, A.; Meerwaldt, J.H. Radiation-induced xerostomia: Pathophysiology, clinical course and supportive treatment. Support Care Cancer 1997, 5, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Fuks, Z.; Glatstein, E.; Marsa, G.W.; Bagshaw, M.A.; Kaplan, H.S. Long-term effects on external radiation on the pituitary and thyroid glands. Cancer 1976, 37, 1152–1161. [Google Scholar] [CrossRef]
- Wang, K.; Tobillo, R.; Mavroidis, P.; Pappafotis, R.; Pearlstein, K.A.; Moon, D.H.; Mahbooba, Z.M.; Deal, A.M.; Holmes, J.A.; Sheets, N.C.; et al. Prospective assessment of patient-reported dry eye syndrome after whole brain radiation. Int. J. Radiat. Oncol. Biol. Phys. 2019, 105, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Nanda, T.; Wu, C.C.; Campbell, A.A.; Bathras, R.M.; Jani, A.; Kazim, M.; Wang, T.J.C. Risk of dry eye syndrome in patients treated with whole-brain radiotherapy. Med. Dosim. 2017, 42, 357–362. [Google Scholar] [CrossRef]
- Thariat, J.; Maschi, C.; Lanteri, S.; Peyrichon, M.L.; Baillif, S.; Herault, J.; Salleron, J.; Caujolle, J.P. Dry Eye Syndrome After Proton Therapy of Ocular Melanomas. Int. J. Radiat. Oncol. Biol. Phys. 2017, 98, 142–151. [Google Scholar] [CrossRef]
- Bhandare, N.; Moiseenko, V.; Song, W.Y.; Morris, C.G.; Bhatti, M.T. Mendenhall WM. Severe dry eye syndrome after radiotherapy for head-and-neck tumors. Int. J. Radiat. Oncol. Biol. Phys. 2012, 82, 1501–1508. [Google Scholar] [CrossRef]
- Miljanovic, B.; Dana, R.; Sullivan, D.A.; Schaumberg, D.A. Impact of dry eye syndrome on vision-related quality of life. Am. J. Ophthalmol. 2007, 143, 409–415. [Google Scholar] [CrossRef]
- Labbe, A.; Wang, Y.X.; Jie, Y.; Baudouin, C.; Jonas, J.B.; Xu, L. Dry eye disease, dry eye symptoms and depression: the Beijing Eye Study. Br. J. Ophthalmol. 2013, 97, 1399–1403. [Google Scholar] [CrossRef]
- Halterman, J.A.; Kwon, H.M.; Wamhoff, B.R. Tonicity-independent regulation of the osmosensitive transcription factor TonEBP (NFAT5). Am. J. Physiol. Cell Physiol. 2012, 302, C1–C8. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, M.; Im, Y.S.; Choi, W.; Byeon, S.H.; Lee, H.K. NFAT5 induction and its role in hyperosmolar stressed human limbal epithelial cells. Invest. Ophthalmol. Vis. Sci. 2008, 49, 1827–1835. [Google Scholar] [CrossRef]
- Joon, H.; Lee Kim, J.W.; Im, Y.S.; Seong, G.J.; Lee, H.K. Cyclosporine A Induces Nerve Growth Factor Expression Via Activation of MAPK p38 and NFAT5. Cornea 2011, 30, S19–S24. [Google Scholar]
- Ramakrishnan, N.; Wolfe, W.W.; Catravas, G.N. Radioprotection of hematopoietic tissues in mice by lipoic acid. Radiat Res. 1992, 130, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, K.M.; Jung, M.H.; Jung, J.H.; Kang, K.M.; Jeong, B.K.; Kim, J.P.; Park, J.J.; Woo, H.H. Protective effects of alpha lipoic acid on radiation-induced salivary gland injury in rats. Oncotarget 2016, 7, 29143–29153. [Google Scholar] [CrossRef] [PubMed]
- Bilska, A.; Wlodek, L. Lipoic acid-the drug of the future? Pharm. Rep. 2005, 57, 570–577. [Google Scholar]
- Warcoin, E.; Baudouin, C.; Gard, C.; Brignole-Baudouin, F. In Vitro Inhibition of NFAT5-Mediated Induction of CCL2 in Hyperosmotic Conditions by Cyclosporine and Dexamethasone on Human HeLa-Modified Conjunctiva-Derived Cells. PLoS ONE 2016, 11, e0159983. [Google Scholar] [CrossRef] [PubMed]
- Hollborn, M.; Fischer, S.; Kuhrt, H.; Wiedemann, P.; Bringmann, A.; Kohen, L. Osmotic regulation of NFAT5 expression in RPE cells: The involvement of purinergic receptor signaling. Mol. Vis. 2017, 23, 116–130. [Google Scholar]
- Jung, J.H.; Jung, J.; Kim, S.K.; Woo, S.H.; Kang, K.M.; Jeong, B.K.; Jung, M.H.; Kim, J.Y.; Hahm, J.R. Alpha lipoic acid attenuates radiation-induced thyroid injury in Rats. PLoS ONE 2014, 9, e112253. [Google Scholar] [CrossRef]
- Jeong, B.K.; Song, J.H.; Jeong, H.; Choi, H.S.; Jung, J.H.; Hahm, J.R.; Woo, S.H.; Jung, M.H.; Choi, B.-H.; Kim, J.H.; et al. Effect of alpha-lipoic acid on radiation-induced small intestine injury in mice. Oncotarget 2016, 7, 15105–15117. [Google Scholar] [CrossRef]
- Kim, J.H.; Jung, M.H.; Kim, J.P.; Kim, H.J.; Jung, J.H.; Hahm, J.R.; Kang, K.M.; Jeong, B.K.; Woo, S.H. Alpha lipoic acid attenuates radiation-induced oral mucositis in rats. Oncotarget 2017, 8, 72739–72747. [Google Scholar] [CrossRef]
- Chung, I.; Hah, Y.S.; Ju, S.; Kim, J.H.; Yoo, W.S.; Cho, H.Y.; Yoo, J.M.; Seo, S.W.; Choi, W.S.; Kim, S.J. Ultraviolet B Radiation Stimulates the Interaction between Nuclear Factor of Activated T Cells 5 (NFAT5) and Nuclear Factor-Kappa B (NF-kappaB) in Human Lens Epithelial Cells. Curr. Eye Res. 2017, 42, 987–994. [Google Scholar] [CrossRef]
- Lee, N.; Kim, D.; Kim, W.U. Role of NFAT5 in the Immune System and Pathogenesis of Autoimmune Diseases. Front. Immunol. 2019, 10, 270. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Ferraris, J.D.; Cai, Q.; Agarwal, A.; Burg, M.B. Increased reactive oxygen species contribute to high NaCl-induced activation of the osmoregulatory transcription factor TonEBP/OREBP. Am. J. Physiol. Renal Physiol. 2005, 289, F377–F385. [Google Scholar] [CrossRef] [PubMed]
- Roth, I.; Leroy, V.; Kwon, H.M.; Martin, P.Y.; Feraille, E.; Hasler, U. Osmoprotective transcription factor NFAT5/TonEBP modulates nuclear factor-kappaB activity. Mol. Biol. Cell 2010, 21, 3459–3474. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Matsuzawa, A.; Nishitoh, H.; Ichijo, H. Roles of MAPKKK ASK1 in stress-induced cell death. Cell Struct. Funct. 2003, 28, 23–29. [Google Scholar] [CrossRef]
- Chang, L.; Karin, M. Mammalian MAP kinase signalling cascades. Nature 2001, 410, 37–40. [Google Scholar] [CrossRef]
- Hirai, S.; Ishida, N.; Watanabe, K.; Mita, S. Leakage of aquaporin 5 in the tear of dacryoadenitis mice. Invest. Ophthalmol. Vis. Sci. 2000, 41, 2432–2437. [Google Scholar]
- Batth, S.S.; Sreeraman, R.; Dienes, E.; Beckett, L.A.; Daly, M.E.; Cui, J.; Mathai, M.; Purdy, J.A.; Chen, A.M. Clinical-dosimetric relationship between lacrimal gland dose and ocular toxicity after intensity-modulated radiotherapy for sinonasal tumours. Br. J. Radiol. 2013, 86, 20130459. [Google Scholar] [CrossRef]
- Craig, J.P.; Nichols, K.K.; Akpek, E.K.; Caffery, B.; Dua, H.S.; Joo, C.K.; Liu, Z.; Nelson, J.D.; Nichols, J.J.; Tsubota, K.; et al. TFOS DEWS II Definition and Classification Report. Ocul. Surf. 2017, 15, 276–283. [Google Scholar] [CrossRef]
- De Herbelet, S.; Vlieghere, E.; de Goncalves, A.; Paepe, B.; Schmidt, K.; Nys, E.; Weynants, L.; Weis, J.; Peer, G.V.; Vandesompele, J.; et al. Localization and Expression of Nuclear Factor of Activated T-Cells 5 in Myoblasts Exposed to Pro-inflammatory Cytokines or Hyperosmolar Stress and in Biopsies from Myositis Patients. Front Physiol. 2018, 9, 126. [Google Scholar] [CrossRef]
- Sawazaki, R.; Ishihara, T.; Usui, S.; Hayashi, E.; Tahara, K.; Hoshino, T.; Higuchi, A.; Nakamura, S.; Tsubota, K.; Mizushima, T. Diclofenac protects cultured human corneal epithelial cells against hyperosmolarity and ameliorates corneal surface damage in a rat model of dry eye. Invest. Ophthalmol Vis. Sci. 2014, 55, 2547–2556. [Google Scholar] [CrossRef]
- Min, C.; Eddy, S.F.; Sherr, D.H.; Sonenshein, G.E. NF-kappaB and epithelial to mesenchymal transition of cancer. J. Cell Biochem. 2008, 104, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Van Waes, C. Nuclear factor-kappaB in development, prevention, and therapy of cancer. Clin. Cancer Res. 2007, 13, 1076–1082. [Google Scholar] [CrossRef] [PubMed]
- Waetzig, V.; Czeloth, K.; Hidding, U.; Mielke, K.; Kanzow, M.; Brecht, S.; Hanisch, U.K. c-Jun N-terminal kinases (JNKs) mediate pro-inflammatory actions of microglia. Glia 2005, 50, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.B.; Schrader, J.W.; Kim, S.U. p38 map kinase regulates TNF-alpha production in human astrocytes and microglia by multiple mechanisms. Cytokine 2000, 12, 874–880. [Google Scholar] [CrossRef] [PubMed]
- Andrade, A.S.; Salomon, T.B.; Behling, C.S.; Mahl, C.D.; Hackenhaar, F.S.; Putti, J.; Benfato, M.S. Alpha-lipoic acid restores tear production in an animal model of dry eye. Exp. Eye Res. 2014, 120, 1–9. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Yoo, W.-S.; Jung, J.H.; Jeong, B.K.; Woo, S.H.; Kim, J.H.; Kim, S.J. Alpha-Lipoic Acid Ameliorates Radiation-Induced Lacrimal Gland Injury through NFAT5-Dependent Signaling. Int. J. Mol. Sci. 2019, 20, 5691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225691
Kim H, Yoo W-S, Jung JH, Jeong BK, Woo SH, Kim JH, Kim SJ. Alpha-Lipoic Acid Ameliorates Radiation-Induced Lacrimal Gland Injury through NFAT5-Dependent Signaling. International Journal of Molecular Sciences. 2019; 20(22):5691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225691
Chicago/Turabian StyleKim, Hyuna, Woong-Sun Yoo, Jung Hwa Jung, Bae Kwon Jeong, Seung Hoon Woo, Jin Hyun Kim, and Seong Jae Kim. 2019. "Alpha-Lipoic Acid Ameliorates Radiation-Induced Lacrimal Gland Injury through NFAT5-Dependent Signaling" International Journal of Molecular Sciences 20, no. 22: 5691. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225691