Long Noncoding RNA (lncRNA)-Mediated Competing Endogenous RNA Networks Provide Novel Potential Biomarkers and Therapeutic Targets for Colorectal Cancer



Abstract
:1. Introduction
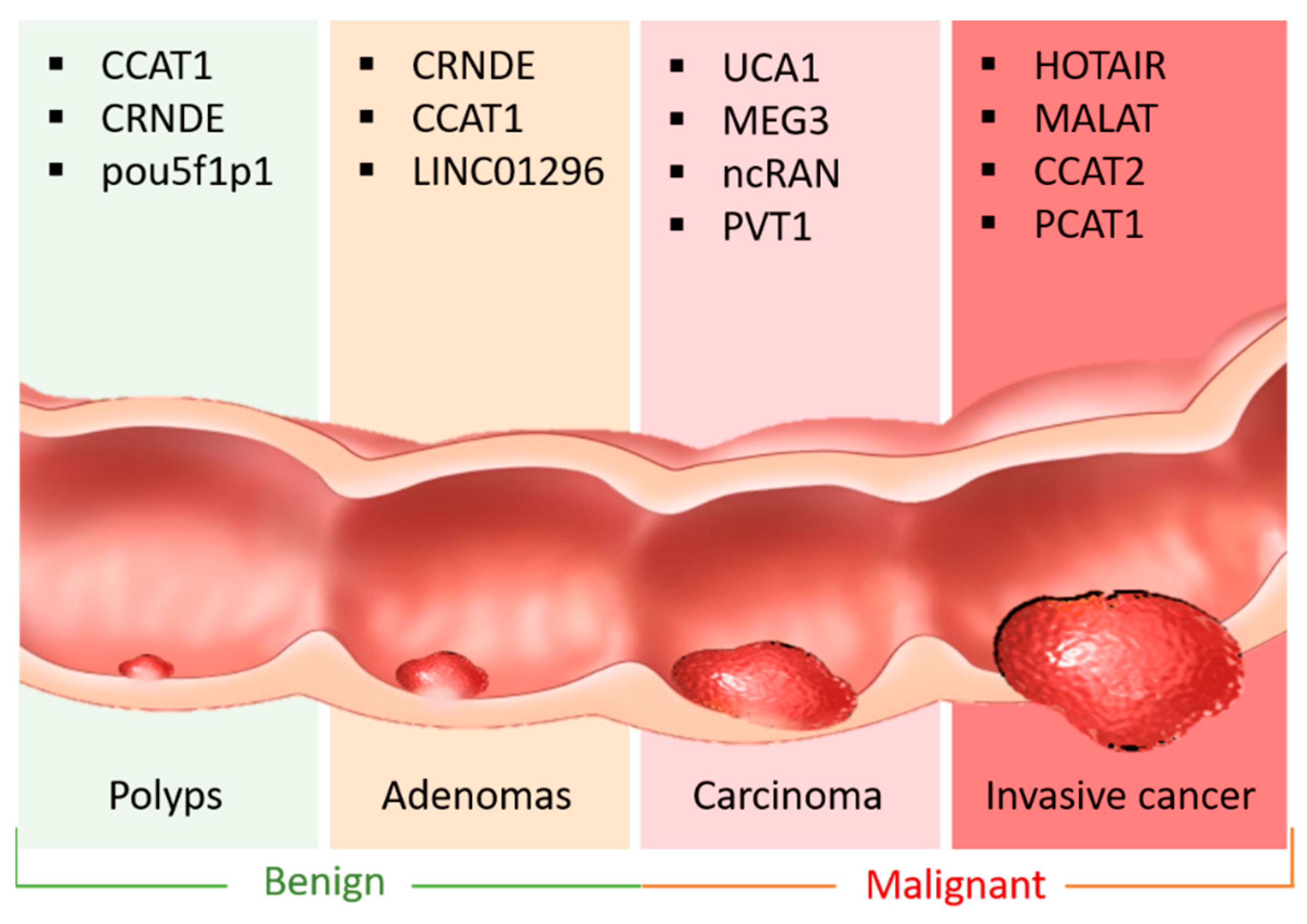
2. LncRNA in CRC
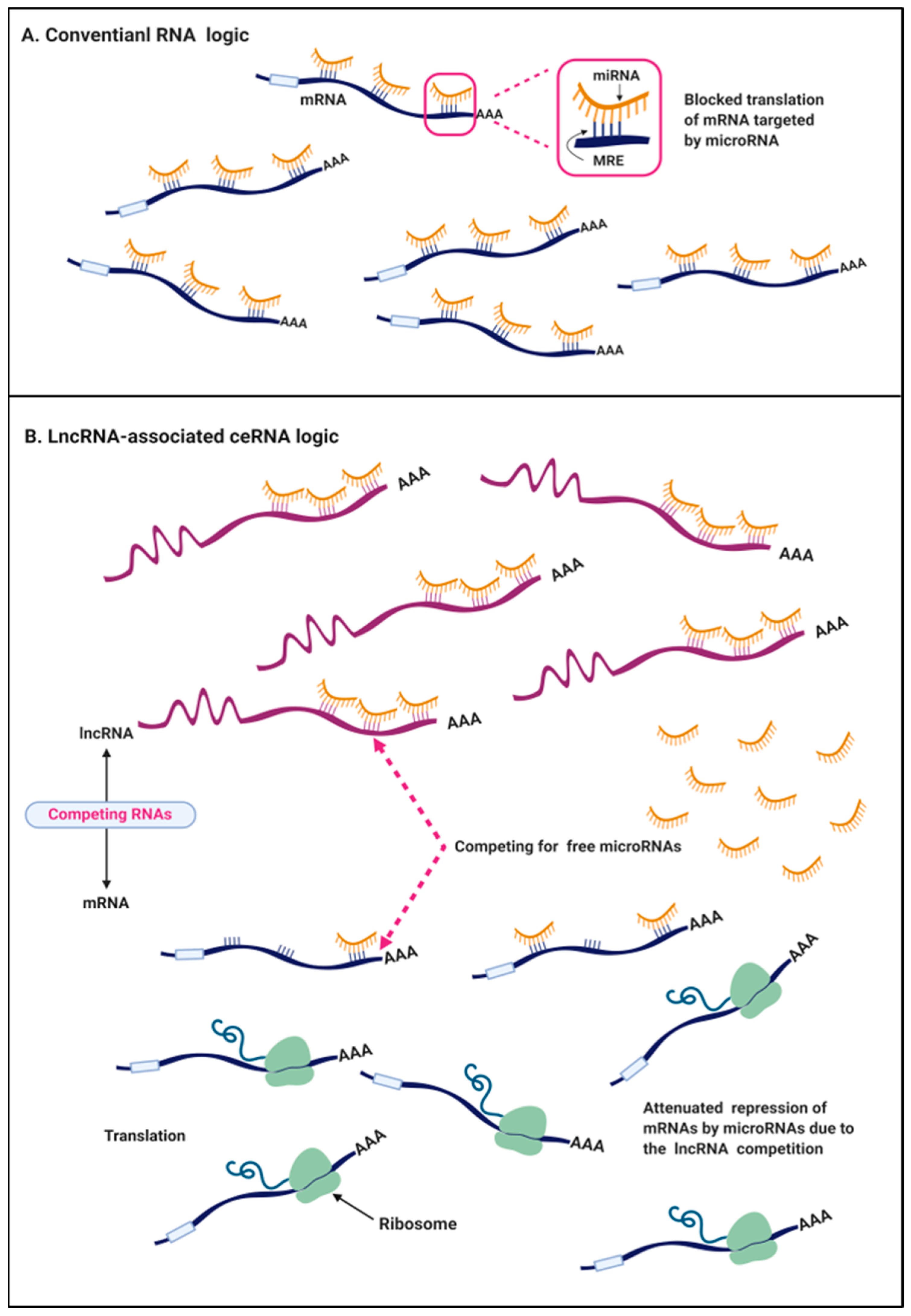
3. The ceRNA Hypothesis in Cancers
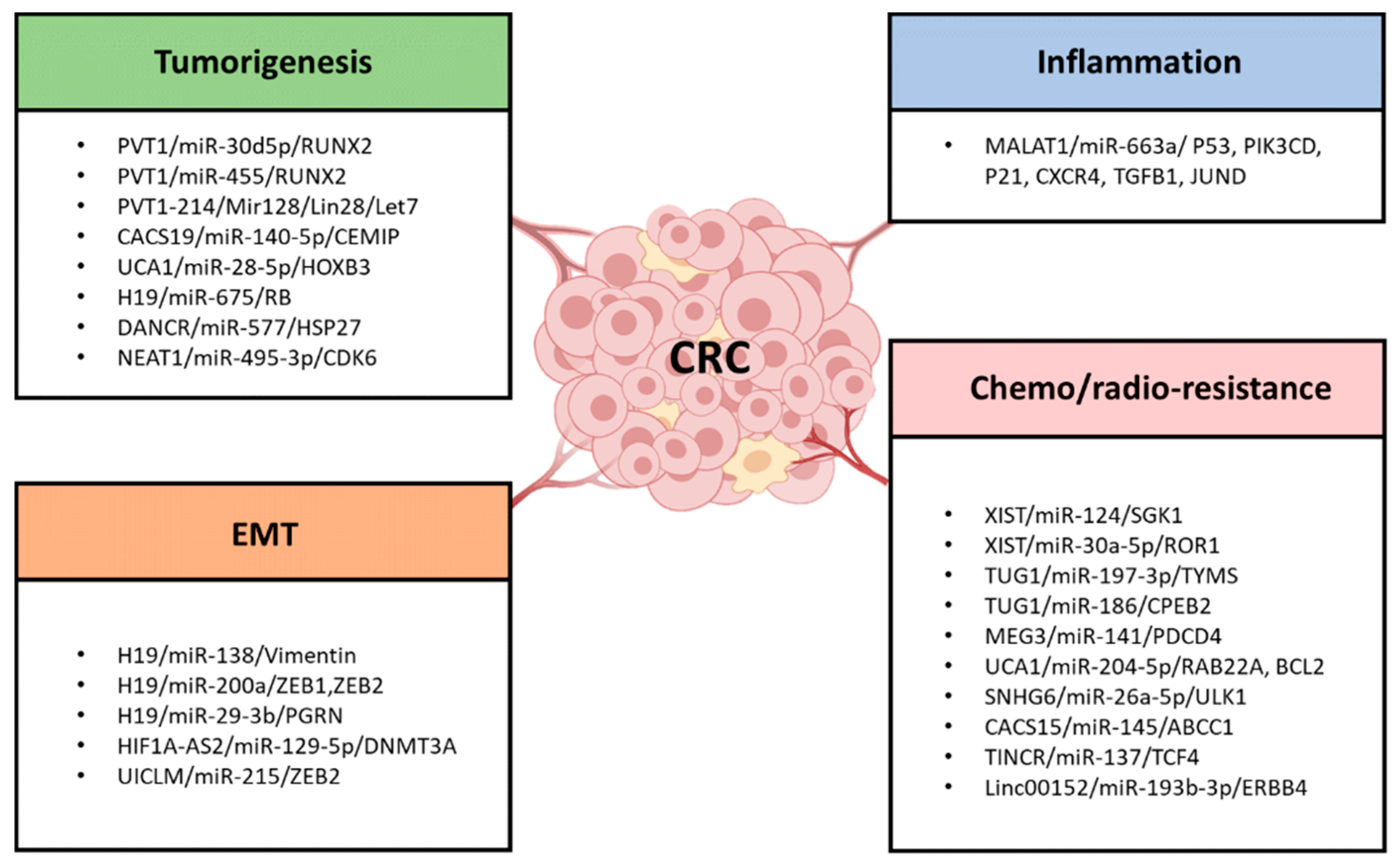
4. LncRNAs as ceRNA in CRC Tumorigenesis and Progression
4.1. PVT1/miR-30d-5p/RUNX2
4.2. PVT1/miR-455/RUNX2
4.3. PVT1-214/Mir128/Lin28/Let7
4.4. CACS19/miR-140-5p/CEMIP
4.5. UCA1/miR-28-5p/HOXB3
4.6. Other lncRNA/miRNA/mRNA Axis
5. LncRNAs as ceRNA in the EMT and Cell Stemness Formation in CRC
5.1. H19
5.1.1. H19/miR-138/Vimentin and H19/miR-200a/ZEB1, ZEB2
5.1.2. H19/miR-29-3b/PGRN
5.2. HIF1A-AS2/miR-129-5p/DNMT3A
5.3. UICLM/miR-215/ZEB2
6. LncRNA as ceRNA in the Inflammation Formation in CRC
7. LncRNA as ceRNA in Chemoresistance/Radioresistance of CRC
7.1. XIST
7.2. TUG1
7.3. MEG3
7.4. UCA1
7.5. Linc00152
7.6. SNHG6
7.7. CACS15
7.8. TINCR
8. Databases for the Prediction of lncRNA-Associated ceRNA Interactions
9. Conclusions and Future Outlook
Funding
Conflicts of Interest
Abbreviations
| ABCC1 | ATP Binding Cassette Subfamily C Member 1 |
| AKT | AKT Serine/Threonine Kinase 1 |
| BCAR4 | Breast Cancer Anti-Estrogen Resistance 4 |
| BCL2 | B-Cell CLL/Lymphoma 2 |
| CACS15 | Cancer Susceptibility 15 |
| CASC19 | Cancer Susceptibility 19 |
| CASC2 | Cancer Susceptibility 2 |
| CCAT1 | Colon Cancer Associated Transcript 1 |
| CCAT2 | Colon Cancer Associated Transcript 2 |
| CDC42 | Cell Division Cycle 42 |
| CDK1 | Cyclin-Dependent Kinase 1 |
| CDK6 | Cyclin-Dependent Kinase 6 |
| CDK9 | Cyclin-Dependent Kinase 9 |
| CEMIP | Cell Migration Inducing Hyaluronidase 1 |
| CPEB2 | Cytoplasmic Polyadenylation Element Binding Protein 2 |
| CRNDE | colorectal neoplasia differentially expressed |
| CXCR4 | C-X-C Motif Chemokine Receptor 4 |
| CYTOR | Long Intergenic Non-Protein Coding RNA 152 |
| DANCR | Differentiation antagonizing non-protein noding RNA |
| DNMT3A | DNA Methyltransferase 3 Alpha |
| ERBB2 | Erb-B2 Receptor Tyrosine Kinase 4 |
| EZH2 | Enhancer of Zeste 2 Polycomb Repressive Complex 2 |
| FOXD2-AS1 | FOXD2 Antisense RNA 1 |
| FOXD3-AS1 | FOXD3 Antisense RNA 1 |
| GACAT3 | Gastric Cancer Associated Transcript 3 |
| HAND2-AS1 | HAND2 Antisense RNA 1 |
| HAGLROS | HAGLR opposite strand lncRNA |
| HIF1A-AS2 | HIF1A Antisense RNA 2 |
| HOTAIR | HOX Transcript Antisense RNA |
| HSP27 | Heat Shock Protein 27 |
| HULC | Hepatocellular Carcinoma Associated Transcript 1 |
| KCNQ1OT1 | KCNQ1 opposite strand/antisense transcript 1 |
| KIAA1199 | Cell Migration Inducing Hyaluronidase 1 |
| KLF14 | Kruppel-Like Factor 14 |
| KRAS | K-Ras |
| LGR5 | Leucine Rich Repeat Containing G Protein-Coupled Receptor 5 |
| LIMK2 | LIM Domain Kinase 2 |
| LINC00460 | Long Intergenic Non-Protein Coding RNA 460 |
| LINC00668 | Long Intergenic Non-Protein Coding RNA 668 |
| LINC00858 | Long Intergenic Non-Protein Coding RNA 858 |
| LINC01234 | Long Intergenic Non-Protein Coding RNA 1234 |
| LINC01296 | Long Intergenic Non-Protein Coding RNA 1296 |
| LINC02418 | Long Intergenic Non-Protein Coding RNA 2418 |
| LSD1 | Lysine Demethylase 1A |
| MACC1 | MET Transcriptional Regulator MACC1 |
| MALAT1 | Metastasis Associated Lung Adenocarcinoma Transcript 1 |
| MBNL1-AS1 | MBNL1 Antisense RNA 1 |
| MEG3 | Maternally Expressed 3 |
| MELK | Maternal Embryonic Leucine Zipper Kinase |
| MIAT | Myocardial Infarction Associated Transcript |
| MIR17HG | MIR17 Host Gene |
| MNX1-AS1 | MNX1 Antisense RNA 1 |
| MYL9 | Myosin Light Chain 9 |
| NEAT1 | Nuclear Paraspeckle Assembly Transcript 1 |
| OECC | Overexpressed in Colorectal Cancer lncRNA |
| PART-1 | Prostate Androgen-Regulated Transcript 1 |
| PCAT1 | prostate cancer associated transcript 1 |
| PDCD4 | Programmed Cell Death 4 |
| PDCD4 | Programmed Cell Death 4 |
| PGRN | Progranulin |
| PIAS3 | Protein Inhibitor Of Activated STAT 3 |
| PIK3CD | Phosphatidylinositol-4,5-Bisphosphate 3-Kinase Catalytic Subunit Delta |
| POU3F3 | POU class 3 homeobox 3 |
| POU5F1P1 | POU class 5 homeobox 1B |
| PRC2 | Polycomb Repressive Complex 2 |
| PTEN | Phosphatase And Tensin Homolog |
| PTEN | Phosphatase And Tensin Homolog |
| PVT1 | Plasmacytoma Variant Translocation 1 |
| RAB22A | Ras-Related Protein Rab-22A |
| ROR | Long Intergenic Non-Protein Coding RNA, Regulator Of Reprogramming |
| ROR1 | Receptor Tyrosine Kinase Like Orphan Receptor 1 |
| RTKN | Rhotekin |
| RUNX2 | Runt-Related Transcription Factor 2 |
| SEC61A1 | SEC61 Translocon Alpha 1 Subunit |
| SHMT2 | Serine Hydroxymethyltransferase 2 |
| SIRT1 | Sirtuin 1 |
| SNHG15 | Small Nucleolar RNA Host Gene 15 |
| SNHG16 | Small Nucleolar RNA Host Gene 16 |
| SNRHG6 | Small Nucleolar RNA Host Gene 6 |
| SP1 | Sp1 Transcription Factor |
| STAT3 | Signal Transducer And Activator Of Transcription 3 |
| TCF4 | Transcription Factor 4 |
| TGF-a | Transforming Growth Factor Alpha |
| TGFB1 | Transforming Growth Factor Beta 1 |
| TINCR | Tissue Differentiation-Inducing Non-Protein Coding RNA |
| TOP2A | Topoisomerase (DNA) II Alpha |
| TP73-AS1 | lncRNA P73 antisense RNA 1T |
| TUG1 | Taurine Up-Regulated 1 |
| TUSC7 | Tumor Suppressor Candidate 7 |
| TYMS | Thymidylate Synthetase |
| UCA1 | Urothelial Cancer Associated 1 |
| UCC | A novel lincRNA termed upregulated in CRC |
| ULK1 | Unc-51 Like Autophagy Activating Kinase 1 |
| USP47 | Ubiquitin Specific Peptidase 47 |
| VEGFC | Vascular Endothelial Growth Factor C |
| XIST | X Inactive Specific Transcript |
| YWHAZ | Tyrosine 3-Monooxygenase/Tryptophan 5-Monooxygenase Activation Protein Zeta |
| ZDHHC8P1 | Zinc Finger DHHC-Type Containing 8 Pseudogene 1 |
| ZEB1 | Zinc Finger E-Box Binding Homeobox 1 |
| ZEB2 | Zinc Finger E-Box Binding Homeobox 2 |
| ZFAS1 | ZNFX1 Antisense RNA 1 |
| ZNF148 | Zinc Finger Protein 148 |
| ZNFX1-AS1 | ZNFX1 Antisense RNA 1 |
References
- Macrae, F.A. Colorectal Cancer: Epidemiology, Risk Factors, and Protective Factors. Available online: https://www.uptodate.com/contents/colorectal-cancer-epidemiology-risk-factors-and-protective-factors (accessed on 25 October 2019).
- Fearnhead, N.S.; Wilding, J.L.; Bodmer, W.F. Genetics of colorectal cancer: Hereditary aspects and overview of colorectal tumorigenesis. Br. Med. Bull. 2002, 64, 27–43. [Google Scholar] [CrossRef]
- Dong, Y.; Yu, T.; Ding, L.; Laurini, E.; Huang, Y.; Zhang, M.; Weng, Y.; Lin, S.; Chen, P.; Marson, D.; et al. A Dual Targeting Dendrimer-Mediated siRNA Delivery System for Effective Gene Silencing in Cancer Therapy. J. Am. Chem. Soc. 2018, 140, 16264–16274. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, A.; Miyata, K.; Kataoka, K. Recent progress in development of siRNA delivery vehicles for cancer therapy. Adv. Drug Deliv. Rev. 2016, 104, 61–77. [Google Scholar] [CrossRef]
- Katsushima, K.; Natsume, A.; Ohka, F.; Shinjo, K.; Hatanaka, A.; Ichimura, N.; Sato, S.; Takahashi, S.; Kimura, H.; Totoki, Y.; et al. Targeting the Notch-regulated non-coding RNA TUG1 for glioma treatment. Nat. Commun. 2016, 7, 13616. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xie, Z.; Xie, C.; Lu, W.; Gao, C.; Ren, H.; Ying, M.; Wei, X.; Gao, J.; Su, B.; et al. D-SP5 Peptide-Modified Highly Branched Polyethylenimine for Gene Therapy of Gastric Adenocarcinoma. Bioconj. Chem. 2015, 26, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lei, Y.; Xie, C.; Lu, W.; Wagner, E.; Xie, Z.; Gao, J.; Zhang, X.; Yan, Z.; Liu, M. Retro-inverso CendR peptide-mediated polyethyleneimine for intracranial glioblastoma-targeting gene therapy. Bioconj. Chem. 2014, 25, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lei, Y.; Xie, C.; Lu, W.; Yan, Z.; Gao, J.; Xie, Z.; Zhang, X.; Liu, M. Targeted gene delivery to glioblastoma using a C-end rule RGERPPR peptide-functionalised polyethylenimine complex. Int. J. Pharm. 2013, 458, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.J.; Tay, Y. Noncoding RNA: RNA Regulatory Networks in Cancer. Int. J. Mol. Sci. 2018, 19, 1310. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Orom, U.A.; Derrien, T.; Beringer, M.; Gumireddy, K.; Gardini, A.; Bussotti, G.; Lai, F.; Zytnicki, M.; Notredame, C.; Huang, Q.; et al. Long noncoding RNAs with enhancer-like function in human cells. Cell 2010, 143, 46–58. [Google Scholar] [CrossRef]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Pichler, M.; Chen, M.; Calin, G.A.; Ling, H. To Wnt or Lose: The Missing Non-Coding Linc in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 2003. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.C.; Zhu, X.; Qiu, J.J.; Xu, J.; Wei, Y. Involvement of long non-coding RNA in colorectal cancer: From benchtop to bedside (Review). Oncol. Lett. 2015, 9, 1039–1045. [Google Scholar] [CrossRef] [PubMed]
- Saus, E.; Brunet-Vega, A.; Iraola-Guzman, S.; Pegueroles, C.; Gabaldon, T.; Pericay, C. Long Non-Coding RNAs As Potential Novel Prognostic Biomarkers in Colorectal Cancer. Front. Genet. 2016, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Lieberman, J.; Lal, A. Desperately seeking microRNA targets. Nat. Struct. Mol. Biol. 2010, 17, 1169–1174. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef]
- Seitz, H. Redefining microRNA targets. Curr. Biol. 2009, 19, 870–873. [Google Scholar] [CrossRef]
- Selbach, M.; Schwanhausser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread changes in protein synthesis induced by microRNAs. Nature 2008, 455, 58–63. [Google Scholar] [CrossRef]
- Lee, D.Y.; Jeyapalan, Z.; Fang, L.; Yang, J.; Zhang, Y.; Yee, A.Y.; Li, M.; Du, W.W.; Shatseva, T.; Yang, B.B. Expression of versican 3’-untranslated region modulates endogenous microRNA functions. PLoS ONE 2010, 5, e13599. [Google Scholar] [CrossRef] [PubMed]
- Cazalla, D.; Yario, T.; Steitz, J.A. Down-regulation of a host microRNA by a Herpesvirus saimiri noncoding RNA. Science 2010, 328, 1563–1566. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, X.; Wu, H.; Ni, P.; Gu, Z.; Qiao, Y.; Chen, N.; Sun, F.; Fan, Q. CREB up-regulates long non-coding RNA, HULC expression through interaction with microRNA-372 in liver cancer. Nucl. Acids Res. 2010, 38, 5366–5383. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.Y.; Moriarity, B.S.; Gong, W.; Akiyama, R.; Tiwari, A.; Kawakami, H.; Ronning, P.; Reuland, B.; Guenther, K.; Beadnell, T.C.; et al. PVT1 dependence in cancer with MYC copy-number increase. Nature 2014, 512, 82–86. [Google Scholar] [CrossRef]
- Yu, X.; Zhao, J.; He, Y. Long non-coding RNA PVT1 functions as an oncogene in human colon cancer through miR-30d-5p/RUNX2 axis. J. B.U.ON Off. J. Balk. Union Oncol. 2018, 23, 48–54. [Google Scholar]
- Gokulnath, M.; Partridge, N.C.; Selvamurugan, N. Runx2, a target gene for activating transcription factor-3 in human breast cancer cells. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2015, 36, 1923–1931. [Google Scholar] [CrossRef]
- Tandon, M.; Chen, Z.; Pratap, J. Runx2 activates PI3K/Akt signaling via mTORC2 regulation in invasive breast cancer cells. Breast Cancer Res. 2014, 16, R16. [Google Scholar] [CrossRef]
- Qin, L.; Zhang, Y.; Lin, J.; Shentu, Y.; Xie, X. MicroRNA-455 regulates migration and invasion of human hepatocellular carcinoma by targeting Runx2. Oncol. Rep. 2016, 36, 3325–3332. [Google Scholar] [CrossRef]
- Wen, C.; Liu, X.; Ma, H.; Zhang, W.; Li, H. miR3383p suppresses tumor growth of ovarian epithelial carcinoma by targeting Runx2. Int. J. Oncol. 2015, 46, 2277–2285. [Google Scholar] [CrossRef]
- Chai, J.; Guo, D.; Ma, W.; Han, D.; Dong, W.; Guo, H.; Zhang, Y. A feedback loop consisting of RUNX2/LncRNA-PVT1/miR-455 is involved in the progression of colorectal cancer. Am. J. Cancer Res. 2018, 8, 538–550. [Google Scholar]
- He, F.; Song, Z.; Chen, H.; Chen, Z.; Yang, P.; Li, W.; Yang, Z.; Zhang, T.; Wang, F.; Wei, J.; et al. Long noncoding RNA PVT1-214 promotes proliferation and invasion of colorectal cancer by stabilizing Lin28 and interacting with miR-128. Oncogene 2019, 38, 164–179. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.D.; Lu, J.; Lin, Y.S.; Gao, C.; Qi, F. Functional role of long non-coding RNA CASC19/miR-140-5p/CEMIP axis in colorectal cancer progression in vitro. World J. Gastroenterol. 2019, 25, 1697–1714. [Google Scholar] [CrossRef] [PubMed]
- Banach, A.; Jiang, Y.P.; Roth, E.; Kuscu, C.; Cao, J.; Lin, R.Z. CEMIP upregulates BiP to promote breast cancer cell survival in hypoxia. Oncotarget 2019, 10, 4307–4320. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, W.; Xu, X.; Zhu, J.; Wu, Y.; Zhao, K.; He, S.; Li, M.; Wu, Y.; Zhang, S.; et al. Downregulation of long noncoding RNA UCA1 enhances the radiosensitivity and inhibits migration via suppression of epithelialmesenchymal transition in colorectal cancer cells. Oncol. Rep. 2018, 40, 1554–1564. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Yang, Y.N.; Yuan, H.H.; Zhang, T.T.; Sui, H.; Wei, X.L.; Liu, L.; Huang, P.; Zhang, W.J.; Bai, Y.X. UCA1, a long non-coding RNA up-regulated in colorectal cancer influences cell proliferation, apoptosis and cell cycle distribution. Pathology 2014, 46, 396–401. [Google Scholar] [CrossRef]
- Neve, B.; Jonckheere, N.; Vincent, A.; Van Seuningen, I. Epigenetic Regulation by lncRNAs: An Overview Focused on UCA1 in Colorectal Cancer. Cancers 2018, 10, 440. [Google Scholar] [CrossRef] [Green Version]
- Bian, Z.; Jin, L.; Zhang, J.; Yin, Y.; Quan, C.; Hu, Y.; Feng, Y.; Liu, H.; Fei, B.; Mao, Y.; et al. LncRNA-UCA1 enhances cell proliferation and 5-fluorouracil resistance in colorectal cancer by inhibiting miR-204-5p. Sci. Rep. 2016, 6, 23892. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.; Chen, M.; Shen, Z.; Wang, R.; Fang, X.; Song, B. LncRNA-UCA1 modulates progression of colon cancer through regulating the miR-28-5p/HOXB3 axis. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef]
- Li, X.; Wang, H.; Yao, B.; Xu, W.; Chen, J.; Zhou, X. lncRNA H19/miR-675 axis regulates cardiomyocyte apoptosis by targeting VDAC1 in diabetic cardiomyopathy. Sci. Rep. 2016, 6, 36340. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Lu, Z.; Wang, N.; Feng, J.; Zhang, J.; Luan, L.; Zhao, W.; Zeng, X. Long noncoding RNA DANCR promotes colorectal cancer proliferation and metastasis via miR-577 sponging. Exp. Mol. Med. 2018, 50, 57. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.H.; Li, J.; Liu, Y.; Liu, Q.Z.; Hao, L.Q.; Liu, L.J.; Zhang, W. ZNF148 modulates TOP2A expression and cell proliferation via ceRNA regulatory mechanism in colorectal cancer. Medicine 2017, 96, e5845. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Dang, J.; Song, A.; Cui, X.; Ma, Z.; Zhang, Z. NEAT1 promotes colon cancer progression through sponging miR-495-3p and activating CDK6 in vitro and in vivo. J. Cell. Physiol. 2019, 234, 19582–19591. [Google Scholar] [CrossRef] [PubMed]
- Ishiwata, T. Cancer stem cells and epithelial-mesenchymal transition: Novel therapeutic targets for cancer. Pathol. Int. 2016, 66, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Bates, R.C.; Pursell, B.M.; Mercurio, A.M. Epithelial-mesenchymal transition and colorectal cancer: Gaining insights into tumor progression using LIM 1863 cells. Cells Tissues Organs 2007, 185, 29–39. [Google Scholar] [CrossRef]
- Bates, R.C. Colorectal cancer progression: Integrin alphavbeta6 and the epithelial-mesenchymal transition (EMT). Cell Cycle (Georget. Tex.) 2005, 4, 1350–1352. [Google Scholar] [CrossRef] [Green Version]
- Yap, A.S. The morphogenetic role of cadherin cell adhesion molecules in human cancer: A thematic review. Cancer Investig. 1998, 16, 252–261. [Google Scholar] [CrossRef]
- Thoreson, M.A.; Reynolds, A.B. Altered expression of the catenin p120 in human cancer: Implications for tumor progression. Differ. Res. Biol. Divers. 2002, 70, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Mendez, M.G.; Kojima, S.; Goldman, R.D. Vimentin induces changes in cell shape, motility, and adhesion during the epithelial to mesenchymal transition. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010, 24, 1838–1851. [Google Scholar] [CrossRef] [Green Version]
- Skarkova, V.; Kralova, V.; Vitovcova, B.; Rudolf, E. Selected Aspects of Chemoresistance Mechanisms in Colorectal Carcinoma-A Focus on Epithelial-to-Mesenchymal Transition, Autophagy, and Apoptosis. Cells 2019, 8, 234. [Google Scholar] [CrossRef] [Green Version]
- Ikeguchi, M.; Taniguchi, T.; Makino, M.; Kaibara, N. Reduced E-cadherin expression and enlargement of cancer nuclei strongly correlate with hematogenic metastasis in colorectal adenocarcinoma. Scand. J. Gastroenterol. 2000, 35, 839–846. [Google Scholar] [CrossRef]
- Lugli, A.; Zlobec, I.; Minoo, P.; Baker, K.; Tornillo, L.; Terracciano, L.; Jass, J.R. Prognostic significance of the wnt signalling pathway molecules APC, beta-catenin and E-cadherin in colorectal cancer: A tissue microarray-based analysis. Histopathology 2007, 50, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Roca, F.; Mauro, L.V.; Morandi, A.; Bonadeo, F.; Vaccaro, C.; Quintana, G.O.; Specterman, S.; de Kier Joffe, E.B.; Pallotta, M.G.; Puricelli, L.I.; et al. Prognostic value of E-cadherin, beta-catenin, MMPs (7 and 9), and TIMPs (1 and 2) in patients with colorectal carcinoma. J. Surg. Oncol. 2006, 93, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Zhou, M.; Tian, B.; Wu, B.; Li, J. Expression of lncRNA-CCAT1, E-cadherin and N-cadherin in colorectal cancer and its clinical significance. Int. J. Clin. Exp. Med. 2015, 8, 3707–3715. [Google Scholar] [PubMed]
- Shan, Z.Z.; Yan, X.B.; Yan, L.L.; Tian, Y.; Meng, Q.C.; Qiu, W.W.; Zhang, Z.; Jin, Z.M. Overexpression of Tbx3 is correlated with Epithelial-Mesenchymal Transition phenotype and predicts poor prognosis of colorectal cancer. Am. J. Cancer Res. 2015, 5, 344–353. [Google Scholar]
- Ying, L.; Chen, Q.; Wang, Y.; Zhou, Z.; Huang, Y.; Qiu, F. Upregulated MALAT-1 contributes to bladder cancer cell migration by inducing epithelial-to-mesenchymal transition. Mol. BioSyst. 2012, 8, 2289–2294. [Google Scholar] [CrossRef]
- Bhan, A.; Mandal, S.S. LncRNA HOTAIR: A master regulator of chromatin dynamics and cancer. Biochim. Biophys. Acta 2015, 1856, 151–164. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.C.; Fu, W.M.; Wong, C.W.; Wang, Y.; Wang, W.M.; Hu, G.X.; Zhang, L.; Xiao, L.J.; Wan, D.C.; Zhang, J.F.; et al. The lncRNA H19 promotes epithelial to mesenchymal transition by functioning as miRNA sponges in colorectal cancer. Oncotarget 2015, 6, 22513–22525. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, X.; Li, X.; Li, X.; Chen, Z. LncRNA H19 promotes epithelial-mesenchymal transition (EMT) by targeting miR-484 in human lung cancer cells. J. Cell. Biochem. 2018, 119, 4447–4457. [Google Scholar] [CrossRef]
- Zhou, W.; Ye, X.L.; Xu, J.; Cao, M.G.; Fang, Z.Y.; Li, L.Y.; Guan, G.H.; Liu, Q.; Qian, Y.H.; Xie, D. The lncRNA H19 mediates breast cancer cell plasticity during EMT and MET plasticity by differentially sponging miR-200b/c and let-7b. Sci. Signal. 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Nong, K.; Zhu, H.; Wang, W.; Huang, X.; Yuan, Z.; Ai, K. H19 promotes pancreatic cancer metastasis by derepressing let-7’s suppression on its target HMGA2-mediated EMT. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2014, 35, 9163–9169. [Google Scholar] [CrossRef]
- Xu, J.; Lamouille, S.; Derynck, R. TGF-beta-induced epithelial to mesenchymal transition. Cell Res. 2009, 19, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Park, S.M.; Gaur, A.B.; Lengyel, E.; Peter, M.E. The miR-200 family determines the epithelial phenotype of cancer cells by targeting the E-cadherin repressors ZEB1 and ZEB2. Genes Dev. 2008, 22, 894–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregory, P.A.; Bert, A.G.; Paterson, E.L.; Barry, S.C.; Tsykin, A.; Farshid, G.; Vadas, M.A.; Khew-Goodall, Y.; Goodall, G.J. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat. Cell Biol. 2008, 10, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, C.; Chen, Z.; Jin, Y.; Wang, Y.; Kolokythas, A.; Dai, Y.; Zhou, X. MicroRNA-138 suppresses epithelial-mesenchymal transition in squamous cell carcinoma cell lines. Biochem. J. 2011, 440, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Long, L.; Huang, G.; Zhu, H.; Guo, Y.; Liu, Y.; Huo, J. Down-regulation of miR-138 promotes colorectal cancer metastasis via directly targeting TWIST2. J. Transl. Med. 2013, 11, 275. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Liu, D.; Feng, Z.; Mao, J.; Zhang, C.; Lu, Y.; Li, J.; Zhang, Q.; Li, Q.; Li, L. MicroRNA-138 modulates metastasis and EMT in breast cancer cells by targeting vimentin. Biomed. Pharmacother. 2016, 77, 135–141. [Google Scholar] [CrossRef]
- Jin, D.; Fang, Y.; Li, Z.; Chen, Z.; Xiang, J. Epithelial-mesenchymal transitionassociated microRNAs in colorectal cancer and drug-targeted therapies (Review). Oncol. Rep. 2015, 33, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Wang, X.; Tang, C.; Chen, X.; He, J. H19 promotes the migration and invasion of colon cancer by sponging miR-138 to upregulate the expression of HMGA1. Int. J. Oncol. 2017, 50, 1801–1809. [Google Scholar] [CrossRef] [Green Version]
- Fu, F.; Wang, T.; Wu, Z.; Feng, Y.; Wang, W.; Zhou, S.; Ma, X.; Wang, S. HMGA1 exacerbates tumor growth through regulating the cell cycle and accelerates migration/invasion via targeting miR-221/222 in cervical cancer. Cell Death Dis. 2018, 9, 594. [Google Scholar] [CrossRef]
- Ding, D.; Li, C.; Zhao, T.; Li, D.; Yang, L.; Zhang, B. LncRNA H19/miR-29b-3p/PGRN Axis Promoted Epithelial-Mesenchymal Transition of Colorectal Cancer Cells by Acting on Wnt Signaling. Mol. Cells 2018, 41, 423–435. [Google Scholar] [CrossRef]
- Chen, L.; Li, Q.; Wang, J.; Jin, S.; Zheng, H.; Lin, J.; He, F.; Zhang, H.; Ma, S.; Mei, J.; et al. MiR-29b-3p promotes chondrocyte apoptosis and facilitates the occurrence and development of osteoarthritis by targeting PGRN. J. Cell. Mol. Med. 2017, 21, 3347–3359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, T.; Yang, D.; Li, R.; Zhang, L.; Zhao, H.; Shen, Y.; Zhang, X.; Kong, B.; Wang, L. PGRN promotes migration and invasion of epithelial ovarian cancer cells through an epithelial mesenchymal transition program and the activation of cancer associated fibroblasts. Exp. Mol. Pathol. 2016, 100, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhuang, C.; Liu, Y.; Li, J.; Dai, F.; Xia, M.; Zhan, Y.; Lin, J.; Chen, Z.; He, A.; et al. Tetracycline-inducible shRNA targeting antisense long non-coding RNA HIF1A-AS2 represses the malignant phenotypes of bladder cancer. Cancer Lett. 2016, 376, 155–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Wang, M.; Mei, Z.; Cao, W.; Yang, Y.; Wang, Y.; Wen, A. lncRNAs HIF1A-AS2 facilitates the up-regulation of HIF-1alpha by sponging to miR-153-3p, whereby promoting angiogenesis in HUVECs in hypoxia. Biomed. Pharmacother. 2017, 96, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Mineo, M.; Ricklefs, F.; Rooj, A.K.; Lyons, S.M.; Ivanov, P.; Ansari, K.I.; Nakano, I.; Chiocca, E.A.; Godlewski, J.; Bronisz, A. The Long Non-coding RNA HIF1A-AS2 Facilitates the Maintenance of Mesenchymal Glioblastoma Stem-like Cells in Hypoxic Niches. Cell Rep. 2016, 15, 2500–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Shi, Z.; Yu, Z.; He, Z. LncRNA HIF1A-AS2 positively affects the progression and EMT formation of colorectal cancer through regulating miR-129-5p and DNMT3A. Biomed. Pharmacother. 2018, 98, 433–439. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, J. DNA methyltransferases and their roles in tumorigenesis. Biomark. Res. 2017, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.E.; Arends, M.J.; Silva, A.L.; Wyllie, A.H.; Greger, L.; Ito, Y.; Vowler, S.L.; Huang, T.H.; Tavare, S.; Murrell, A.; et al. Sequential DNA methylation changes are associated with DNMT3B overexpression in colorectal neoplastic progression. Gut 2011, 60, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Steine, E.J.; Barrasa, M.I.; Hockemeyer, D.; Pawlak, M.; Fu, D.; Reddy, S.; Bell, G.W.; Jaenisch, R. Deletion of the de novo DNA methyltransferase Dnmt3a promotes lung tumor progression. Proc. Natl. Acad. Sci. USA 2011, 108, 18061–18066. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Wu, Q.; Cheng, J.; Qiu, X.; Zhang, J.; Fan, H. Depletion of DNMT3A suppressed cell proliferation and restored PTEN in hepatocellular carcinoma cell. J. Biomed. Biotechnol. 2010, 2010, 737535. [Google Scholar] [CrossRef] [Green Version]
- Pathania, R.; Ramachandran, S.; Elangovan, S.; Padia, R.; Yang, P.; Cinghu, S.; Veeranan-Karmegam, R.; Arjunan, P.; Gnana-Prakasam, J.P.; Sadanand, F.; et al. DNMT1 is essential for mammary and cancer stem cell maintenance and tumorigenesis. Nat. Commun. 2015, 6, 6910. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.L.; Lu, Y.X.; Zhang, J.X.; Wei, X.L.; Wang, F.; Zeng, Z.L.; Pan, Z.Z.; Yuan, Y.F.; Wang, F.H.; Pelicano, H.; et al. Long non-coding RNA UICLM promotes colorectal cancer liver metastasis by acting as a ceRNA for microRNA-215 to regulate ZEB2 expression. Theranostics 2017, 7, 4836–4849. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, C.N.; Blanchard, J.F.; Kliewer, E.; Wajda, A. Cancer risk in patients with inflammatory bowel disease: A population-based study. Cancer 2001, 91, 854–862. [Google Scholar] [CrossRef]
- Zacharopoulou, E.; Gazouli, M.; Tzouvala, M.; Vezakis, A.; Karamanolis, G. The contribution of long non-coding RNAs in Inflammatory Bowel Diseases. Dig. Liver Dis. 2017, 49, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yao, J.; Xie, J.; Liu, Z.; Zhou, Y.; Pan, H.; Han, W. The role of autophagy in colitis-associated colorectal cancer. Sign. Transduct. Targ. Ther. 2018, 3, 31. [Google Scholar] [CrossRef]
- Yarani, R.; Mirza, A.H.; Kaur, S.; Pociot, F. The emerging role of lncRNAs in inflammatory bowel disease. Exp. Mol. Med. 2018, 50, 161. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.Q.; Huang, M.L.; Xu, A.T.; Zhao, D.; Ran, Z.H.; Shen, J. LncRNA DQ786243 affects Treg related CREB and Foxp3 expression in Crohn’s disease. J. Biomed. Sci. 2013, 20, 87. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Han, X.; Wittfeldt, A.; Sun, J.; Liu, C.; Wang, X.; Gan, L.M.; Cao, H.; Liang, Z. Long non-coding RNA ANRIL regulates inflammatory responses as a novel component of NF-kappaB pathway. RNA Biol. 2016, 13, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Huang, Y.; Dong, F.; Kwon, J.H. Ulcerative Colitis-Associated Long Noncoding RNA, BC012900, Regulates Intestinal Epithelial Cell Apoptosis. Inflamm. Bowel Dis. 2016, 22, 782–795. [Google Scholar] [CrossRef]
- Yang, L.; Wang, H.; Shen, Q.; Feng, L.; Jin, H. Long non-coding RNAs involved in autophagy regulation. Cell Death Dis. 2017, 8, e3073. [Google Scholar] [CrossRef] [Green Version]
- Bermudez, M.; Aguilar-Medina, M.; Lizarraga-Verdugo, E.; Avendano-Felix, M.; Silva-Benitez, E.; Lopez-Camarillo, C.; Ramos-Payan, R. LncRNAs as Regulators of Autophagy and Drug Resistance in Colorectal Cancer. Front. Oncol. 2019, 9, 1008. [Google Scholar] [CrossRef] [PubMed]
- Qian, M.; Fang, X.; Wang, X. Autophagy and inflammation. Clin. Transl. Med. 2017, 6, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, C.; Li, D.; Yang, L.; Jin, J.; Zhang, B. lncRNA KCNQ1OT1 enhances the chemoresistance of oxaliplatin in colon cancer by targeting the miR-34a/ATG4B pathway. OncoTarg. Ther. 2019, 12, 2649–2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, T.D.; Xu, J.H.; Yu, T.; Li, J.Y.; Zhao, L.N.; Ouyang, H.; Luo, S.; Lu, X.J.; Huang, C.Z.; Lan, Q.S.; et al. Knockdown of linc-POU3F3 suppresses the proliferation, apoptosis, and migration resistance of colorectal cancer. Oncotarget 2016, 7, 961–975. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Tan, K.; Huang, H. Long noncoding RNA HAGLROS regulates apoptosis and autophagy in colorectal cancer cells via sponging miR-100 to target ATG5 expression. J. Cell. Biochem. 2019, 120, 3922–3933. [Google Scholar] [CrossRef]
- Yu, Z.; Li, N.; Jiang, K.; Zhang, N.; Yao, L.L. MiR-100 up-regulation enhanced cell autophagy and apoptosis induced by cisplatin in osteosarcoma by targeting mTOR. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5867–5873. [Google Scholar] [CrossRef]
- Ge, Y.Y.; Shi, Q.; Zheng, Z.Y.; Gong, J.; Zeng, C.; Yang, J.; Zhuang, S.M. MicroRNA-100 promotes the autophagy of hepatocellular carcinoma cells by inhibiting the expression of mTOR and IGF-1R. Oncotarget 2014, 5, 6218–6228. [Google Scholar] [CrossRef]
- Liu, C.; Ji, L.; Song, X. Long non coding RNA UCA1 contributes to the autophagy and survival of colorectal cancer cells via sponging miR-185-5p to up-regulate the WISP2/β-catenin pathway. RSC Adv. 2019, 9, 14160–14166. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.; Du, Y.; Ma, Y.; Gu, L.; Zhou, J.; Deng, D. MALAT1-miR663a negative feedback loop in colon cancer cell functions through direct miRNA-lncRNA binding. Cell Death Dis. 2018, 9, 857. [Google Scholar] [CrossRef]
- Yang, H.; Liang, N.; Wang, M.; Fei, Y.; Sun, J.; Li, Z.; Xu, Y.; Guo, C.; Cao, Z.; Li, S.; et al. Long noncoding RNA MALAT-1 is a novel inflammatory regulator in human systemic lupus erythematosus. Oncotarget 2017, 8, 77400–77406. [Google Scholar] [CrossRef]
- Michaille, J.J.; Piurowski, V.; Rigot, B.; Kelani, H.; Fortman, E.C.; Tili, E. MiR-663, a MicroRNA Linked with Inflammation and Cancer That Is under the Influence of Resveratrol. Medicines 2018, 5, 74. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.C. The molecular mechanisms of chemoresistance in cancers. Oncotarget 2017, 8, 59950–59964. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, X.; Wang, H.; Wang, L.; Liu, T.; Du, L.; Yang, Y.; Wang, C. MALAT1 Is Associated with Poor Response to Oxaliplatin-Based Chemotherapy in Colorectal Cancer Patients and Promotes Chemoresistance through EZH2. Mol. Cancer Ther. 2017, 16, 739–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Liu, H.; Yu, J.; Yu, H. Chemoresistance to doxorubicin induces epithelial-mesenchymal transition via upregulation of transforming growth factor beta signaling in HCT116 colon cancer cells. Mol. Med. Rep. 2015, 12, 192–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.; Zhang, R.; Yang, D.; Li, J.; Yan, X.; Jin, K.; Li, W.; Liu, X.; Zhao, J.; Shang, W.; et al. Knockdown of Long Non-Coding RNA XIST Inhibited Doxorubicin Resistance in Colorectal Cancer by Upregulation of miR-124 and Downregulation of SGK1. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 51, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Talarico, C.; Dattilo, V.; D’Antona, L.; Menniti, M.; Bianco, C.; Ortuso, F.; Alcaro, S.; Schenone, S.; Perrotti, N.; Amato, R. SGK1, the New Player in the Game of Resistance: Chemo-Radio Molecular Target and Strategy for Inhibition. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2016, 39, 1863–1876. [Google Scholar] [CrossRef] [PubMed]
- Amato, R.; Menniti, M.; Agosti, V.; Boito, R.; Costa, N.; Bond, H.M.; Barbieri, V.; Tagliaferri, P.; Venuta, S.; Perrotti, N. IL-2 signals through Sgk1 and inhibits proliferation and apoptosis in kidney cancer cells. J. Mol. Med. 2007, 85, 707–721. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, Z.; Yu, Q.; Shen, J.; He, W.; Zhou, D.; Yu, Q.; Fan, J.; Gao, S.; Duan, L. Atractylenolide II reverses the influence of lncRNA XIST/miR-30a-5p/ROR1 axis on chemo-resistance of colorectal cancer cells. J. Cell. Mol. Med. 2019, 23, 3151–3165. [Google Scholar] [CrossRef]
- Cui, B.; Zhang, S.; Chen, L.; Yu, J.; Widhopf, G.F., 2nd; Fecteau, J.F.; Rassenti, L.Z.; Kipps, T.J. Targeting ROR1 inhibits epithelial-mesenchymal transition and metastasis. Cancer Res. 2013, 73, 3649–3660. [Google Scholar] [CrossRef]
- Zhou, J.K.; Zheng, Y.Z.; Liu, X.S.; Gou, Q.; Ma, R.; Guo, C.L.; Croce, C.M.; Liu, L.; Peng, Y. ROR1 expression as a biomarker for predicting prognosis in patients with colorectal cancer. Oncotarget 2017, 8, 32864–32872. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.Y.; Guan, S.H.; Tang, R.N.; Tao, S.J.; Guo, D.A. Simultaneous determination of atractylenolide II and atractylenolide III by liquid chromatography-tandem mass spectrometry in rat plasma and its application in a pharmacokinetic study after oral administration of Atractylodes Macrocephala Rhizoma extract. Biomed. Chromatogr. BMC 2012, 26, 1386–1392. [Google Scholar] [CrossRef] [PubMed]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Hu, H.; Wang, Y.; Huang, Q.; Huang, R.; Chen, Y.; Ma, T.; Qiao, T.; Zhang, Q.; Wu, H.; et al. Long non-coding RNA TUG1 mediates 5-fluorouracil resistance by acting as a ceRNA of miR-197-3p in colorectal cancer. J. Cancer 2019, 10, 4603–4613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertz, R.; Li, M.C.; Spencer, D.B. Effect of methotrexate therapy upon choriocarcinoma and chorioadenoma. Proc. Soc. Exp. Biol. Med. 1956, 93, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Bertino, J.R.; Goker, E.; Gorlick, R.; Li, W.W.; Banerjee, D. Resistance Mechanisms to Methotrexate in Tumors. Oncologist 1996, 1, 223–226. [Google Scholar] [CrossRef]
- Li, C.; Gao, Y.; Li, Y.; Ding, D. TUG1 mediates methotrexate resistance in colorectal cancer via miR-186/CPEB2 axis. Biochem. Biophys. Res. Commun. 2017, 491, 552–557. [Google Scholar] [CrossRef]
- Martinez-Balibrea, E.; Martinez-Cardus, A.; Gines, A.; Ruiz de Porras, V.; Moutinho, C.; Layos, L.; Manzano, J.L.; Buges, C.; Bystrup, S.; Esteller, M.; et al. Tumor-Related Molecular Mechanisms of Oxaliplatin Resistance. Mol. Cancer ther. 2015, 14, 1767–1776. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Shang, J.; Zhang, Y.; Liu, S.; Peng, Y.; Zhou, Z.; Pan, H.; Wang, X.; Chen, L.; Zhao, Q. MEG3 is a prognostic factor for CRC and promotes chemosensitivity by enhancing oxaliplatin-induced cell apoptosis. Oncol. Rep. 2017, 38, 1383–1392. [Google Scholar] [CrossRef]
- Wang, H.; Li, H.; Zhang, L.; Yang, D. Overexpression of MEG3 sensitizes colorectal cancer cells to oxaliplatin through regulation of miR-141/PDCD4 axis. Biomed. Pharmacother. 2018, 106, 1607–1615. [Google Scholar] [CrossRef]
- Afonja, O.; Juste, D.; Das, S.; Matsuhashi, S.; Samuels, H.H. Induction of PDCD4 tumor suppressor gene expression by RAR agonists, antiestrogen and HER-2/neu antagonist in breast cancer cells. Evidence for a role in apoptosis. Oncogene 2004, 23, 8135–8145. [Google Scholar] [CrossRef] [Green Version]
- Shi, G.H.; Ye, D.W.; Yao, X.D.; Zhang, S.L.; Dai, B.; Zhang, H.L.; Shen, Y.J.; Zhu, Y.; Zhu, Y.P.; Xiao, W.J.; et al. Involvement of microRNA-21 in mediating chemo-resistance to docetaxel in androgen-independent prostate cancer PC3 cells. Acta Pharmacol. Sin. 2010, 31, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Shiota, M.; Izumi, H.; Tanimoto, A.; Takahashi, M.; Miyamoto, N.; Kashiwagi, E.; Kidani, A.; Hirano, G.; Masubuchi, D.; Fukunaka, Y.; et al. Programmed cell death protein 4 down-regulates Y-box binding protein-1 expression via a direct interaction with Twist1 to suppress cancer cell growth. Cancer Res. 2009, 69, 3148–3156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, B.; Yu, X.; Guo, X.; Fan, X.; Yang, Z.; Wu, P.; Yuan, Z.; Deng, Y.; Wang, J.; Chen, D.; et al. Increased urothelial cancer associated 1 is associated with tumor proliferation and metastasis and predicts poor prognosis in colorectal cancer. Int. J. Oncol. 2015, 47, 1329–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Shen, B.; Tan, M.; Mu, X.; Qin, Y.; Zhang, F.; Liu, Y. Long non-coding RNA UCA1 increases chemoresistance of bladder cancer cells by regulating Wnt signaling. FEBS J. 2014, 281, 1750–1758. [Google Scholar] [CrossRef]
- Yin, Y.; Zhang, B.; Wang, W.; Fei, B.; Quan, C.; Zhang, J.; Song, M.; Bian, Z.; Wang, Q.; Ni, S.; et al. miR-204-5p inhibits proliferation and invasion and enhances chemotherapeutic sensitivity of colorectal cancer cells by downregulating RAB22A. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2014, 20, 6187–6199. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; He, W.; Lu, Y.; Wang, Y. Long non-coding RNA LINC00152 promotes tumorigenesis via sponging miR-193b-3p in osteosarcoma. Oncol. Lett. 2019, 18, 3630–3636. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Tao, W. Long Noncoding RNA LINC00152 Facilitates the Leukemogenesis of Acute Myeloid Leukemia by Promoting CDK9 Through miR-193a. DNA Cell Biol. 2019, 38, 236–242. [Google Scholar] [CrossRef]
- Yue, B.; Cai, D.; Liu, C.; Fang, C.; Yan, D. Linc00152 Functions as a Competing Endogenous RNA to Confer Oxaliplatin Resistance and Holds Prognostic Values in Colon Cancer. Mol. Ther. J. Am. Soc. Gene Ther. 2016, 24, 2064–2077. [Google Scholar] [CrossRef] [Green Version]
- Mamoori, A.; Wahab, R.; Islam, F.; Lee, K.; Vider, J.; Lu, C.T.; Gopalan, V.; Lam, A.K. Clinical and biological significance of miR-193a-3p targeted KRAS in colorectal cancer pathogenesis. Hum. Pathol. 2018, 71, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.S.; Bernard, J.K.; Demory Beckler, M.; Almohazey, D.; Washington, M.K.; Smith, J.J.; Frey, M.R. ERBB4 is over-expressed in human colon cancer and enhances cellular transformation. Carcinogenesis 2015, 36, 710–718. [Google Scholar] [CrossRef] [Green Version]
- Bian, Z.; Zhang, J.; Li, M.; Feng, Y.; Yao, S.; Song, M.; Qi, X.; Fei, B.; Yin, Y.; Hua, D.; et al. Long non-coding RNA LINC00152 promotes cell proliferation, metastasis, and confers 5-FU resistance in colorectal cancer by inhibiting miR-139-5p. Oncogenesis 2017, 6, 395. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lan, Z.; He, J.; Lai, Q.; Yao, X.; Li, Q.; Liu, Y.; Lai, H.; Gu, C.; Yan, Q.; et al. LncRNA SNHG6 promotes chemoresistance through ULK1-induced autophagy by sponging miR-26a-5p in colorectal cancer cells. Cancer Cell Int. 2019, 19, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonioli, M.; Di Rienzo, M.; Piacentini, M.; Fimia, G.M. Emerging Mechanisms in Initiating and Terminating Autophagy. Trends Biochem. Sci. 2017, 42, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.M.M.; Towers, C.G.; Thorburn, A. Targeting autophagy in cancer. Nat. Rev. Cancer 2017, 17, 528–542. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H.; Young, L.N. Mechanisms of Autophagy Initiation. Annu. Rev. Biochem. 2017, 86, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548–563.e516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Han, D.; Yuan, Z.; Hu, H.; Zhao, Z.; Yang, R.; Jin, Y.; Zou, C.; Chen, Y.; Wang, G.; et al. Long non-coding RNA H19 confers 5-Fu resistance in colorectal cancer by promoting SIRT1-mediated autophagy. Cell Death Dis. 2018, 9, 1149. [Google Scholar] [CrossRef]
- Gao, R.; Fang, C.; Xu, J.; Tan, H.; Li, P.; Ma, L. LncRNA CACS15 contributes to oxaliplatin resistance in colorectal cancer by positively regulating ABCC1 through sponging miR-145. Arch. Biochem. Biophys. 2019, 663, 183–191. [Google Scholar] [CrossRef]
- Ji, D.; Zhan, T.; Li, M.; Yao, Y.; Jia, J.; Yi, H.; Qiao, M.; Xia, J.; Zhang, Z.; Ding, H.; et al. Enhancement of Sensitivity to Chemo/Radiation Therapy by Using miR-15b against DCLK1 in Colorectal Cancer. Stem Cell Rep. 2018, 11, 1506–1522. [Google Scholar] [CrossRef] [Green Version]
- Kang, Z.; Jifu, E.; Guo, K.; Ma, X.; Zhang, Y.; Yu, E. Knockdown of long non-coding RNA TINCR decreases radioresistance in colorectal cancer cells. Pathol. Res. Pract. 2019, 152622. [Google Scholar] [CrossRef]
- Ben-Porath, I.; Thomson, M.W.; Carey, V.J.; Ge, R.; Bell, G.W.; Regev, A.; Weinberg, R.A. An embryonic stem cell-like gene expression signature in poorly differentiated aggressive human tumors. Nat. Genet. 2008, 40, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Li, J.H.; Liu, S.; Zhou, H.; Qu, L.H.; Yang, J.H. starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucl. Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paraskevopoulou, M.D.; Georgakilas, G.; Kostoulas, N.; Reczko, M.; Maragkakis, M.; Dalamagas, T.M.; Hatzigeorgiou, A.G. DIANA-LncBase: Experimentally verified and computationally predicted microRNA targets on long non-coding RNAs. Nucl. Acids Res. 2013, 41, D239–D245. [Google Scholar] [CrossRef] [PubMed]
- Furio-Tari, P.; Tarazona, S.; Gabaldon, T.; Enright, A.J.; Conesa, A. spongeScan: A web for detecting microRNA binding elements in lncRNA sequences. Nucl. Acids Res. 2016, 44, W176–W180. [Google Scholar] [CrossRef]
- Das, S.; Ghosal, S.; Sen, R.; Chakrabarti, J. lnCeDB: Database of human long noncoding RNA acting as competing endogenous RNA. PLoS ONE 2014, 9, e98965. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Ping, Y.; Zhao, H.; Ning, S.; Xia, P.; Wang, W.; Wan, L.; Li, J.; Zhang, L.; Yu, L.; et al. LncNetP, a systematical lncRNA prioritization approach based on ceRNA and disease phenotype association assumptions. Oncotarget 2017, 8, 114603–114612. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, A.; Cui, Y. SomamiR 2.0: A database of cancer somatic mutations altering microRNA-ceRNA interactions. Nucl. Acids Res. 2016, 44, D1005–D1010. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zhi, H.; Zhang, Y.; Liu, Y.; Zhang, J.; Gao, Y.; Guo, M.; Ning, S.; Li, X. miRSponge: A manually curated database for experimentally supported miRNA sponges and ceRNAs. Database J. Biol. Databases Curation 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.; Meng, X.; Li, X.; Illing, N.; Ingle, R.A.; Wang, J.; Chen, M. PceRBase: A database of plant competing endogenous RNA. Nucl. Acids Res. 2017, 45, D1009–D1014. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Yan, Z.; Li, Y.; Sun, Z. Linc2GO: A human LincRNA function annotation resource based on ceRNA hypothesis. Bioinformatics 2013, 29, 2221–2222. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, S.; Zhou, X.; Chen, Z.; Wang, M.; Zheng, X.; Xie, M. LncRNA BCAR4, targeting to miR-665/STAT3 signaling, maintains cancer stem cells stemness and promotes tumorigenicity in colorectal cancer. Cancer Cell Int. 2019, 19, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, N.; Huang, T.; Guo, H.; Yang, J.; Li, M.; Chen, Z.; Zhang, Y. LncRNA CASC15 promotes colon cancer cell proliferation and metastasis by regulating the miR4310/LGR5/Wnt/betacatenin signaling pathway. Mol. Med. Rep. 2018, 18, 2269–2276. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Wu, X.; Li, S.; Xu, X.; Zhu, H.; Chen, X. The long noncoding RNA CASC2 functions as a competing endogenous RNA by sponging miR-18a in colorectal cancer. Sci. Rep. 2016, 6, 26524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Nangia-Makker, P.; Farhana, L.; Majumdar, A.P.N. A novel mechanism of lncRNA and miRNA interaction: CCAT2 regulates miR-145 expression by suppressing its maturation process in colon cancer cells. Mol. Cancer 2017, 16, 155. [Google Scholar] [CrossRef]
- Li, M.; Wang, Q.; Xue, F.; Wu, Y. lncRNA-CYTOR Works as an Oncogene Through the CYTOR/miR-3679-5p/MACC1 Axis in Colorectal Cancer. DNA Cell Biol. 2019, 38, 572–582. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zou, Y.; Hu, G.; Lin, C.; Guo, Y.; Gao, K.; Wu, M. Facilitating colorectal cancer cell metastasis by targeted binding of long non-coding RNA ENSG00000231881 with miR-133b via VEGFC signaling pathway. Biochem. Biophys. Res. Commun. 2019, 509, 1–7. [Google Scholar] [CrossRef]
- Yue, B.; Sun, B.; Liu, C.; Zhao, S.; Zhang, D.; Yu, F.; Yan, D. Long non-coding RNA Fer-1-like protein 4 suppresses oncogenesis and exhibits prognostic value by associating with miR-106a-5p in colon cancer. Cancer Sci. 2015, 106, 1323–1332. [Google Scholar] [CrossRef]
- Zhu, Y.; Qiao, L.; Zhou, Y.; Ma, N.; Wang, C.; Zhou, J. Long non-coding RNA FOXD2-AS1 contributes to colorectal cancer proliferation through its interaction with microRNA-185-5p. Cancer Sci. 2018, 109, 2235–2242. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Shi, M.; Meng, W.; Wang, Y.; Hui, P.; Ma, J. Long noncoding RNA FOXD3-AS1 promotes colon adenocarcinoma progression and functions as a competing endogenous RNA to regulate SIRT1 by sponging miR-135a-5p. J. Cell. Physiol. 2019, 234, 21889–21902. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, L.; Miao, Y.; Xing, R. Novel long noncoding RNA GACAT3 promotes colorectal cancer cell proliferation, invasion, and migration through miR-149. OncoTarg. Ther. 2018, 11, 1543–1552. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wang, H.J.; Meng, T.; Lei, C.; Yang, X.H.; Wang, Q.S.; Jin, B.; Zhu, J.F. lncRNA GAS5 Inhibits Cell Migration and Invasion and Promotes Autophagy by Targeting miR-222-3p via the GAS5/PTEN-Signaling Pathway in CRC. Mol. Ther. Nucl. Acids 2019, 17, 644–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Lin, J.; Zhang, H.; Zhu, F.; Xie, R. LncRNA HAND2-AS1 sponging miR-1275 suppresses colorectal cancer progression by upregulating KLF14. Biochem. Biophys. Res. Commun. 2018, 503, 1848–1853. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.L.; Zhao, X.J.; Wei, C.C.; Wu, J.W. LncRNA HOTAIR promotes colon cancer development by down-regulating miRNA-34a. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5752–5761. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wei, M.-H.; Lu, J.-G.; Bi, C.-Y. Long non-coding RNA HULC interacts with miR-613 to regulate colon cancer growth and metastasis through targeting RTKN. Biomed. Pharmacother. 2019, 109, 2035–2042. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, X.; Li, Q.; Zhang, Y. lncRNA LINC00460 promoted colorectal cancer cells metastasis via miR-939-5p sponging. Cancer Manag. Res. 2019, 11, 1779–1789. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Yue, Y.; Wang, J.; Li, W.; Sun, M.; Gu, C.; Zeng, L. LINC00668 promotes tumorigenesis and progression through sponging miR-188–5p and regulating USP47 in colorectal cancer. Eur. J. Pharmacol. 2019, 858, 172464. [Google Scholar] [CrossRef]
- Sha, Q.K.; Chen, L.; Xi, J.Z.; Song, H. Long non-coding RNA LINC00858 promotes cells proliferation, migration and invasion by acting as a ceRNA of miR-22-3p in colorectal cancer. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1057–1066. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Zhang, Y.; Chen, Y.; Bai, Y.; Zhang, Y. Long noncoding RNA LINC01234 promotes serine hydroxymethyltransferase 2 expression and proliferation by competitively binding miR-642a-5p in colon cancer. Cell Death Dis. 2019, 10, 137. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Zhang, M.; Wang, C.; Ning, X. Long Noncoding RNA LINC01296 Harbors miR-21a to Regulate Colon Carcinoma Proliferation and Invasion. Oncol. Res. 2019, 27, 541–549. [Google Scholar] [CrossRef]
- Zhao, Y.; Du, T.; Du, L.; Li, P.; Li, J.; Duan, W.; Wang, Y.; Wang, C. Long noncoding RNA LINC02418 regulates MELK expression by acting as a ceRNA and may serve as a diagnostic marker for colorectal cancer. Cell Death Dis. 2019, 10, 568. [Google Scholar] [CrossRef]
- Zhu, K.; Wang, Y.; Liu, L.; Li, S.; Yu, W. Long non-coding RNA MBNL1-AS1 regulates proliferation, migration, and invasion of cancer stem cells in colon cancer by interacting with MYL9 via sponging microRNA-412-3p. Clin. Res. Hepatol. Gastroenterol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, H.; Cai, H.; Hong, Y.; Li, Y.; Su, D.; Fan, Z. Long non-coding RNA MIAT promotes growth and metastasis of colorectal cancer cells through regulation of miR-132/Derlin-1 pathway. Cancer Cell Int. 2018, 18, 59. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Liu, B.; Han, W.; Zhao, D. LncRNA-MIR17HG mediated upregulation of miR-17 and miR-18a promotes colon cancer progression via activating Wnt/β-catenin signaling. Transl. Cancer Res. 2019, 8, 1097–1108. [Google Scholar] [CrossRef]
- Ye, Y.; Gu, B.; Wang, Y.; Shen, S.; Huang, W. E2F1-mediated MNX1-AS1-miR-218-5p-SEC61A1 feedback loop contributes to the progression of colon adenocarcinoma. J. Cell. Biochem. 2019, 120, 6145–6153. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wen, C.; Zhuansun, Y.; Huang, L.; Chen, W.; Yang, X.; Liu, H. A novel long noncoding RNA OECC promotes colorectal cancer development and is negatively regulated by miR-143-3p. Biochem. Biophys. Res. Commun. 2018, 503, 2949–2955. [Google Scholar] [CrossRef]
- Hu, Y.; Ma, Z.; He, Y.; Liu, W.; Su, Y.; Tang, Z. PART-1 functions as a competitive endogenous RNA for promoting tumor progression by sponging miR-143 in colorectal cancer. Biochem. Biophys. Res. Commun. 2017, 490, 317–323. [Google Scholar] [CrossRef]
- Zhou, P.; Sun, L.; Liu, D.; Liu, C.; Sun, L. Long Non-Coding RNA lincRNA-ROR Promotes the Progression of Colon Cancer and Holds Prognostic Value by Associating with miR-145. Pathol. Oncol. Res. POR 2016, 22, 733–740. [Google Scholar] [CrossRef]
- Sun, X.; Bai, Y.; Yang, C.; Hu, S.; Hou, Z.; Wang, G. Long noncoding RNA SNHG15 enhances the development of colorectal carcinoma via functioning as a ceRNA through miR-141/SIRT1/Wnt/beta-catenin axis. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2536–2544. [Google Scholar] [CrossRef] [Green Version]
- Ke, D.; Wang, Q.; Ke, S.; Zou, L.; Wang, Q. Long-Non Coding RNA SNHG16 Supports Colon Cancer Cell Growth by Modulating miR-302a-3p/AKT Axis. Pathol. Oncol. Res. POR 2019. [Google Scholar] [CrossRef]
- Jia, Z.; Peng, J.; Yang, Z.; Chen, J.; Liu, L.; Luo, D.; He, P. Long non-coding RNA TP73AS1 promotes colorectal cancer proliferation by acting as a ceRNA for miR103 to regulate PTEN expression. Gene 2019, 685, 222–229. [Google Scholar] [CrossRef]
- Cai, Y.; Yan, P.; Zhang, G.; Yang, W.; Wang, H.; Cheng, X. Long non-coding RNA TP73-AS1 sponges miR-194 to promote colorectal cancer cell proliferation, migration and invasion via up-regulating TGFalpha. Cancer Biomark. Sect. Dis. Mark. 2018, 23, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Hu, J.; Wang, G.; Yang, Z.; Zhao, C.; Zhang, X.; Wang, J. LncRNA TUG1 promoted KIAA1199 expression via miR-600 to accelerate cell metastasis and epithelial-mesenchymal transition in colorectal cancer. J. Exp. Clin. Cancer Res. CR 2018, 37, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Zhang, R.; Zhao, J. The Novel Long Noncoding RNA TUSC7 Inhibits Proliferation by Sponging MiR-211 in Colorectal Cancer. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 41, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.T.; Chen, W.Y.; Gu, Z.Q.; Zhuang, Y.Y.; Li, C.Q.; Wang, L.Y.; Peng, J.F.; Zhu, Z.; Luo, X.; Li, Y.H.; et al. The novel long intergenic noncoding RNA UCC promotes colorectal cancer progression by sponging miR-143. Cell Death Dis. 2017, 8, e2778. [Google Scholar] [CrossRef]
- Wu, X.; He, X.; Li, S.; Xu, X.; Chen, X.; Zhu, H. Long Non-Coding RNA ucoo2kmd.1 Regulates CD44-Dependent Cell Growth by Competing for miR-211-3p in Colorectal Cancer. PLoS ONE 2016, 11, e0151287. [Google Scholar] [CrossRef]
- Li, C.; Liu, T.; Zhang, Y.; Li, Q.; Jin, L.K. LncRNA-ZDHHC8P1 promotes the progression and metastasis of colorectal cancer by targeting miR-34a. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1476–1486. [Google Scholar] [CrossRef]
- Mo, D.; Liu, W.; Li, Y.; Cui, W. Long Non-coding RNA Zinc Finger Antisense 1 (ZFAS1) Regulates Proliferation, Migration, Invasion, and Apoptosis by Targeting MiR-7-5p in Colorectal Cancer. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 5150–5158. [Google Scholar] [CrossRef]
- Thorenoor, N.; Faltejskova-Vychytilova, P.; Hombach, S.; Mlcochova, J.; Kretz, M.; Svoboda, M.; Slaby, O. Long non-coding RNA ZFAS1 interacts with CDK1 and is involved in p53-dependent cell cycle control and apoptosis in colorectal cancer. Oncotarget 2016, 7, 622–637. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Hong, X.; Ba, L.; He, X.; Xiong, Y.; Ding, Q.; Yang, S.; Peng, G. Long non-coding RNA ZNFX1-AS1 promotes the tumor progression and metastasis of colorectal cancer by acting as a competing endogenous RNA of miR-144 to regulate EZH2 expression. Cell Death Dis. 2019, 10, 150. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| LncRNA | Chromosome Location | Competitor mRNA | Shared miRNA | ceRNA Network | ceRNA Role | Related CRC Hallmark | Ref. |
|---|---|---|---|---|---|---|---|
| BCAR4 | 16p13.13 | STAT3 | miR-665 | BCAR4/miR-655/STAT3 | Oncogenic | Proliferation, migration | [152] |
| CACS15 | 6p22.3 | LGR5 | miR-4310 | CACS15/miR-4310/LGR5 | Oncogenic | Proliferation, invasion, TNM stage, metastasis | [153] |
| CASC19 | 8q24.21 | CEMIP | miR-140-5p | CASC19/miR-140-5p/CEMIP | Oncogenic | Proliferation, invasion, migration, apoptosis, EMT | [33] |
| CASC2 | 10q26.11 | PIAS3 | miR-18a | CASC2/miR-18a/PIAS3/STAT3 | Tumor suppressive | Proliferation, tumor growth, G0/G1-S phase transition | [154] |
| CCAT2 | 8q24.21 | miR-145 | CCAT2/miR-145/miR-21 | Oncogenic | CSC proliferation and differentiation | [155] | |
| CYTOR | 2p11.2 | MACC1 | miR-3679-5p | CYTOR/miR-3679-5p/MACC1 | Oncogenic | TNM stage, perineural and venous invasions | [156] |
| ENSG00000-231881 | 6 | VEGFC | miR-133b | ENSG00000231881/miR-133b/VEGFC | Oncogenic | Metastasis | [157] |
| FER1L4 | 20q11.22 | miR-106a-5p | FER1L4/miR-106a-5p | Tumor suppressive | Proliferation, cell cycle | [158] | |
| FOXD2-AS1 | 1p33 | CDC42 | miR-185-5p | FOXD2-AS1/miR-185-5p/CDC42 | Oncogenic | Proliferation, migration, invasion | [159] |
| FOXD3-AS1 | 1p31.3 | SIRT1 | miR-135a-5p | FOXD3-AS1/miR-135a-5p/SIRT1 | Oncogenic | Proliferation, migration, invasion, cell cycle, apoptosis | [160] |
| GACAT3 | 2p24.3 | SP1, STAT3 | miR-149 | GACAT3/miR-149/SP1/STAT3 | Oncogenic | Proliferation, invasion, migration | [161] |
| GAS5 | 1q25.1 | PTEN | miR-222-3p | GAS5/miR-222-3p/PTEN | Tumor suppressive | Proliferation, migration, apoptosis | [162] |
| H19 | 11p15.5 | Vimentin, ZEB1, ZEB2 | miR-138, miR-200a | H19/miR-138/Vimentin, H19/miR-200a/ZEB1, H19/miR-200a/ZEB2 | Oncogenic | EMT progression | [58] |
| HAND2-AS1 | 4q34.1 | KLF14 | miR-1275 | HAND2-AS1/miR-1275/KLF14 | Tumor suppressive | Proliferation, invasion | [163] |
| HOTAIR | 12q13.13 | miR-34a | HOTAIR/miR-34a | Oncogenic | Metastasis | [164] | |
| HULC | 6p24.3 | RTKN | miR-613 | HULC/miR-613/RTKN | Oncogenic | Proliferation, metastasis | [165] |
| LINC00460 | 13q33.2 | LIMK2 | miR-939-5p | LINC00460/LIMK2/miR-939-5p | Oncogenic | Metastasis | [166] |
| LINC00668 | 18p11.31 | USP47 | miR-188–5p | LINC000668/miR-188-5p/USP47 | Oncogenic | Proliferation, metastasis | [167] |
| LINC00858 | 10q23.1 | YWHAZ | miR-22-3p | LINC00858/miR-22-3p/YWHAZ | Oncogenic | Proliferation, migration, invasion | [168] |
| LINC01234 | 12q24.13 | SHMT2 | miR-642a-5p | LINC01234/miR-642a-5p/SHMT2 | Oncogenic | Proliferation | [169] |
| LINC01296 | 14q11.2 | PDCD4 | miR-21a | LINC01296/miR-21a/PDCD4 | Oncogenic | Proliferation | [170] |
| LINC02418 | 12q24.33 | MELK | miR-1273g-3p | LINC02418/miR-1273g-3p/MELK | Oncogenic | Proliferation, apoptosis | [171] |
| MALAT1 | 11q13.1 | p53 | miR-663a | MALAT1/miR-663a/p53 | Oncogenic | Proliferation, migration, invasion, apoptosis | [100] |
| MBNL1-AS1 | 3q25.1 | MYL9 | miR-412-3p | MBNL-AS1/miR-412-3p/MYL9 | Tumor suppressive | Proliferation, invasion | [172] |
| MIAT | 22q12.1 | Derlin-1 | miR-132 | MIAT/miR-132/Derlin-1 | Oncogenic | Tumor growth, metastasis | [173] |
| MIR17HG | 13q31.3 | Wnt, β-catenin | miR-17, miR-18a | MIR17HG-miR-17/18a-Wnt/ β-catenin | Oncogenic | Lymph node metastasis, TNM stage | [174] |
| MNX1-AS1 | 7q36.3 | SEC61A1 | miR-218-5p | MNX1-AS1/miR-218-5p/ SEC61A1 | Oncogenic | progression of colon adenocarcinoma | [175] |
| NEAT1 | 11q13.1 | CDK6 | miR-495-3p | NEAT1/miR-495-3p/CDK6 | Oncogenic | Proliferation, migration, invasion | [43] |
| OECC | 8q24 | NF-κB, p38MAPK | miR-143-3p | OECC/miR-143-3p/NF-κB/ p38 MAPK | Oncogenic | Proliferation, apoptosis, migration | [176] |
| PART-1 | 5q12.1 | DNMT3A | miR-143 | PART-1/miR-143/DNMT3A | Oncogenic | Proliferation, metastasis | [177] |
| PVT1 | 8q24.21 | RUNX2 | miR-30d-5p | PVT1/miR-30d-5p/RUNX2 | Oncogenic | Proliferation, metastasis | [26] |
| PVT1 | 8q24.21 | RUNX2 | miR-455 | PVT1/miR-455/RUNX2 | Oncogenic | Proliferation, migration, invasion, apoptosis | [31] |
| PVT1-214 | 8q24.21 | Lin28 | miR-128 | PVT1-214/miR-128/Lin28/let-7 | Oncogenic | Proliferation, invasion | [32] |
| ROR | 18q21.31 | miR-145 | ROR/miR-145 | Oncogenic | Proliferation, migration, invasion | [178] | |
| SNHG15 | 7p13 | SIRT1 | miR-141 | SNHG15/miR-141/SIRT1 | Oncogenic | Proliferation, apoptosis | [179] |
| SNHG16 | 17q25.1 | AKT | miR-302a-3p | SNHG16/miR-302a-3p/AKT | Oncogenic | Proliferation | [180] |
| TP73-AS1 | 1p36.32 | PTEN | miR-103 | TP73-AS1/miR-103/PTEN | Tumor suppressive | Proliferation | [181] |
| TP73-AS1 | 1p36.32 | TGF-a | miR-194 | TP73-AS1/miR-194/TGF-a | Oncogenic | Proliferation, migration, invasion | [182] |
| TUG1 | 22q12.2 | KIAA1199 | miR-600 | TUG1/miR-600/KIAA1199 | Oncogenic | Metastasis, EMT | [183] |
| TUSC7 | 3q13.31 | CDK6 | miR-211-3p | TUSC7/miR-211-3p/CDK6 | Tumor suppressive | Proliferation | [184] |
| UCA1 | 19p13.12 | HOXB3 | miR-28-5p | UCA1/miR-28-5p/HOXB3 | Oncogenic | Proliferation, migration | [39] |
| UCC | 7p15.2 | KRAS | miR-143 | UCC/miR-143/KRAS | Oncogenic | Cell growth, invasion | [185] |
| ucoo2kmd.1 | 17q11.2 | CD44 | miR-211-3p | ucoo2lmd.1/miR-211-3p/CD44 | Oncogenic | Proliferation | [186] |
| ZDHHC8P1 | 22q11.23 | miR-34a | ZDHHC8P1/miRNA-34a | Oncogenic | Proliferation, metastasis | [187] | |
| ZFAS1 | 20q13.13 | miR-7-5p | ZFAS1/miR-7-5p | Oncogenic | Proliferation, migration, invasion, apoptosis | [188] | |
| ZFAS1 | 20q13.13 | CDK1/ cyclinB1, p53 | miR-590-3p | ZFAS1/miR-590-3p | Oncogenic | Apoptosis, p53 dependent cell cycle control | [189] |
| ZNFX1-AS1 | 20q13.13 | EZH2 | miR-144 | ZNFX1-AS1/miR-144/EZH2 | Oncogenic | Proliferation, migration, invasion, metastasis | [190] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Cho, K.B.; Li, Y.; Tao, G.; Xie, Z.; Guo, B. Long Noncoding RNA (lncRNA)-Mediated Competing Endogenous RNA Networks Provide Novel Potential Biomarkers and Therapeutic Targets for Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 5758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225758
Wang L, Cho KB, Li Y, Tao G, Xie Z, Guo B. Long Noncoding RNA (lncRNA)-Mediated Competing Endogenous RNA Networks Provide Novel Potential Biomarkers and Therapeutic Targets for Colorectal Cancer. International Journal of Molecular Sciences. 2019; 20(22):5758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225758
Chicago/Turabian StyleWang, Liye, Kwang Bog Cho, Yan Li, Gabriel Tao, Zuoxu Xie, and Bin Guo. 2019. "Long Noncoding RNA (lncRNA)-Mediated Competing Endogenous RNA Networks Provide Novel Potential Biomarkers and Therapeutic Targets for Colorectal Cancer" International Journal of Molecular Sciences 20, no. 22: 5758. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225758