Transcriptome Profiling Unravels a Vital Role of Pectin and Pectinase in Anther Dehiscence in Chrysanthemum

 ,
,
Abstract
:1. Introduction
2. Results
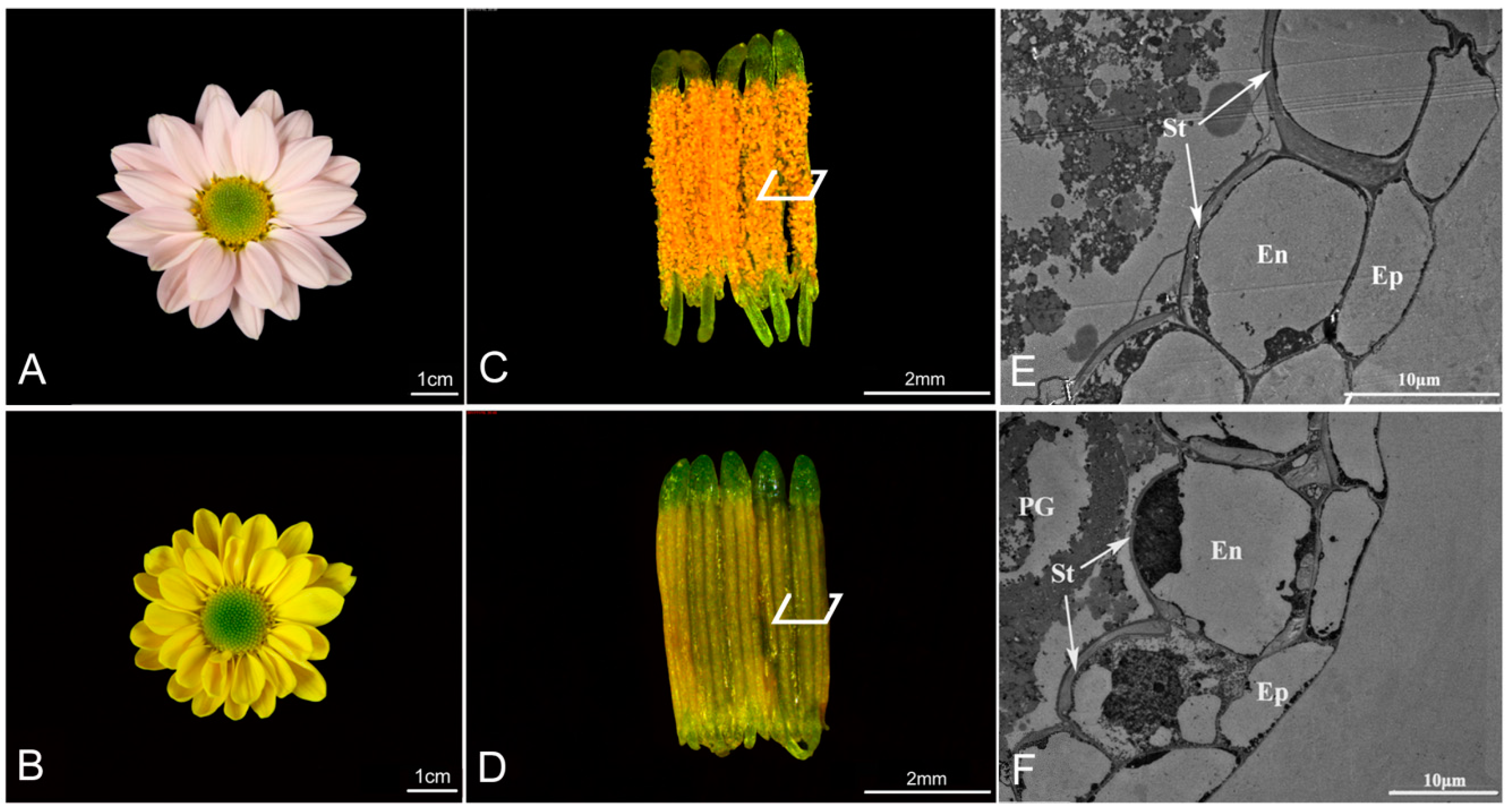
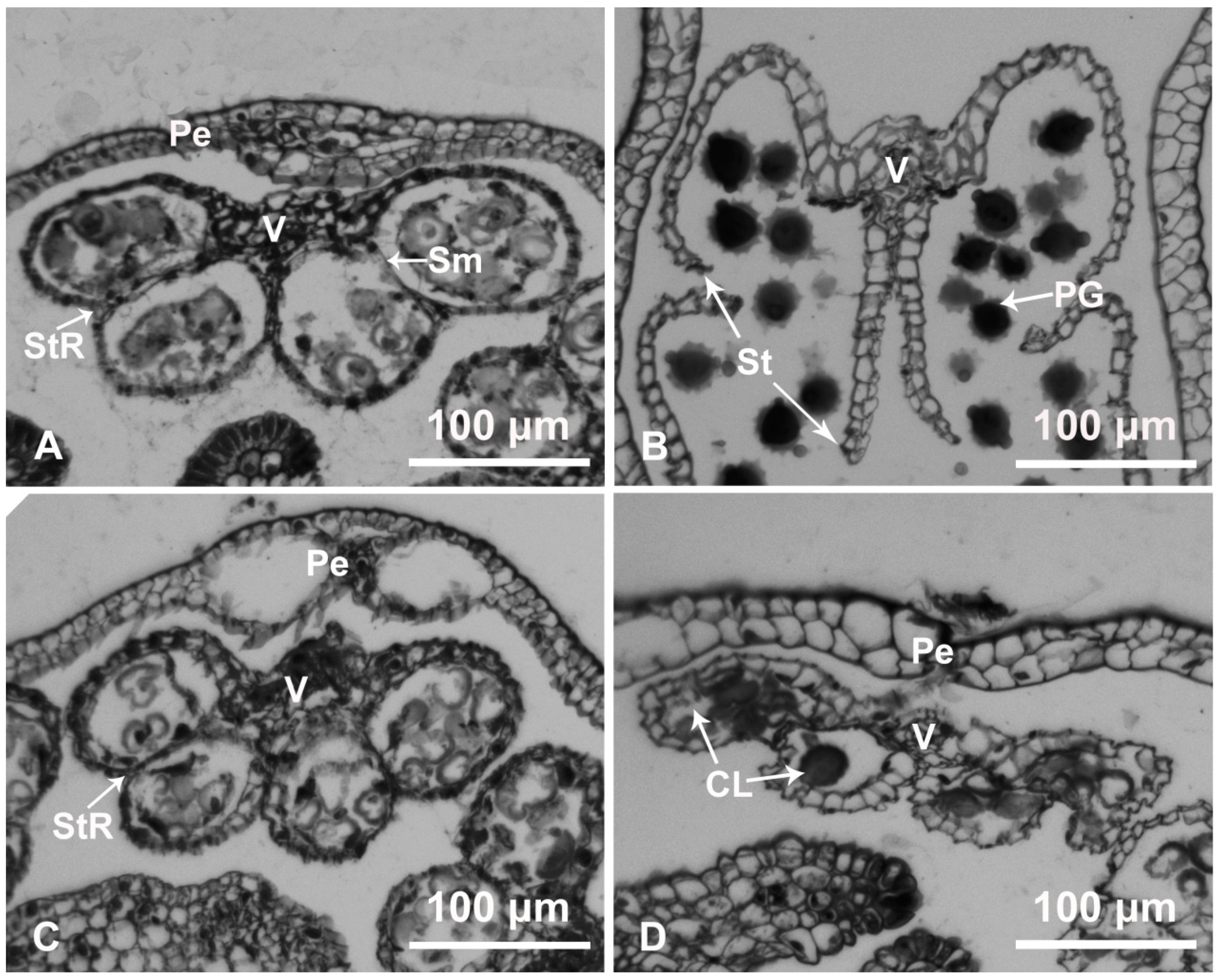
2.1. Anther Development and Dehiscence in Chrysanthemum
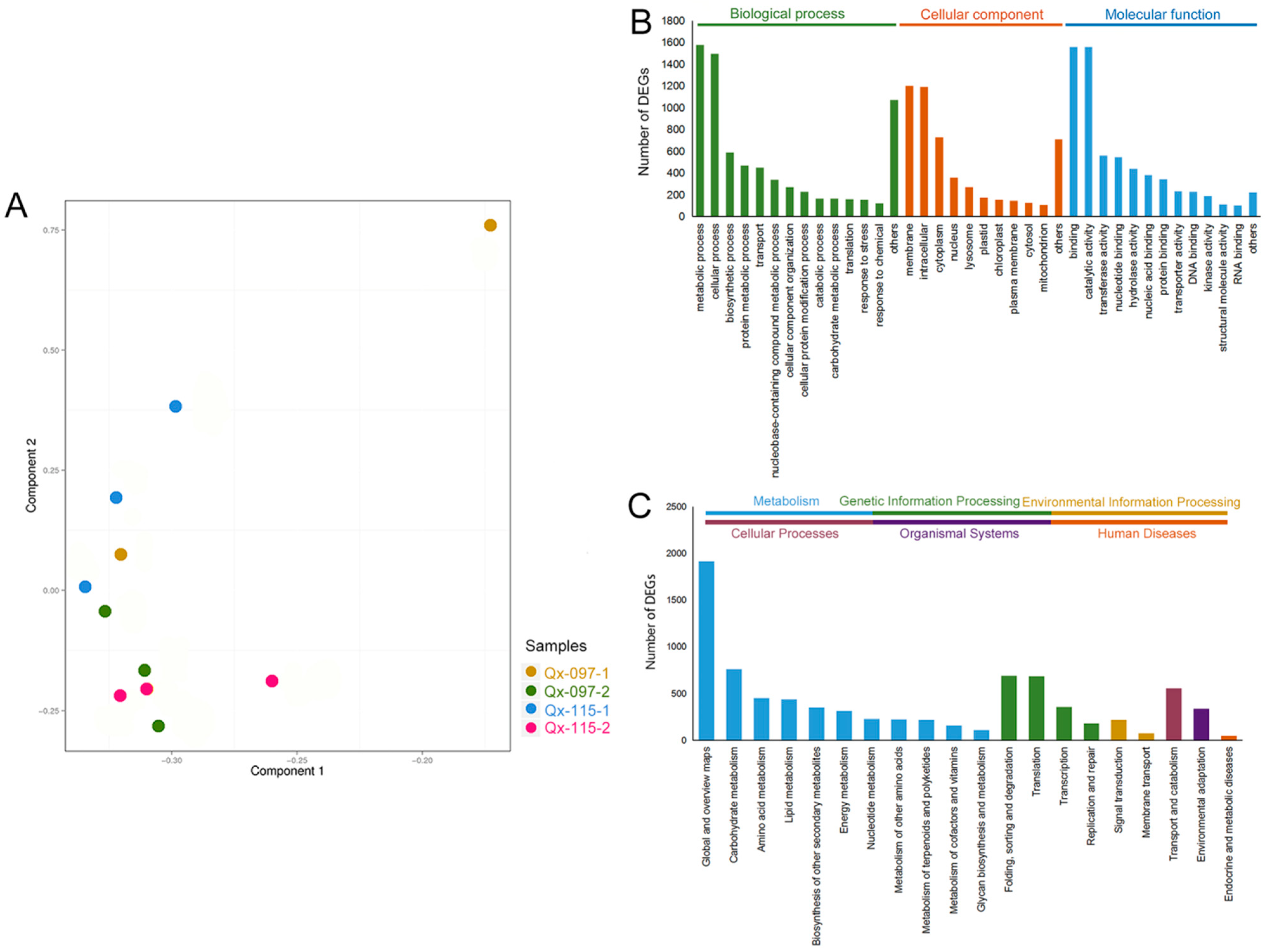
2.2. RNA-Seq Analysis and Read Assembly
2.3. Gene Annotation and Functional Classification
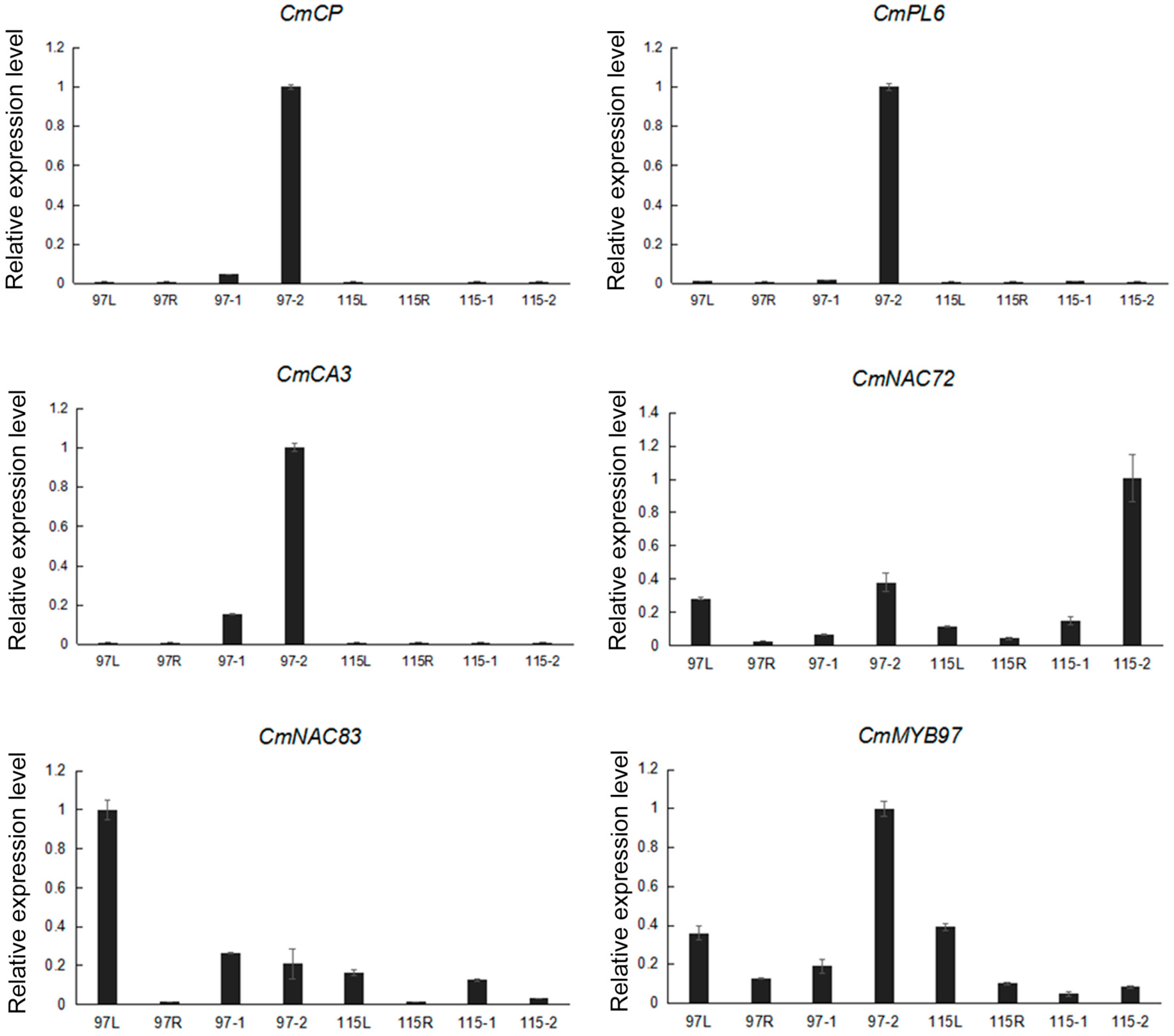
2.4. Validation of Gene Expression by qRT-PCR
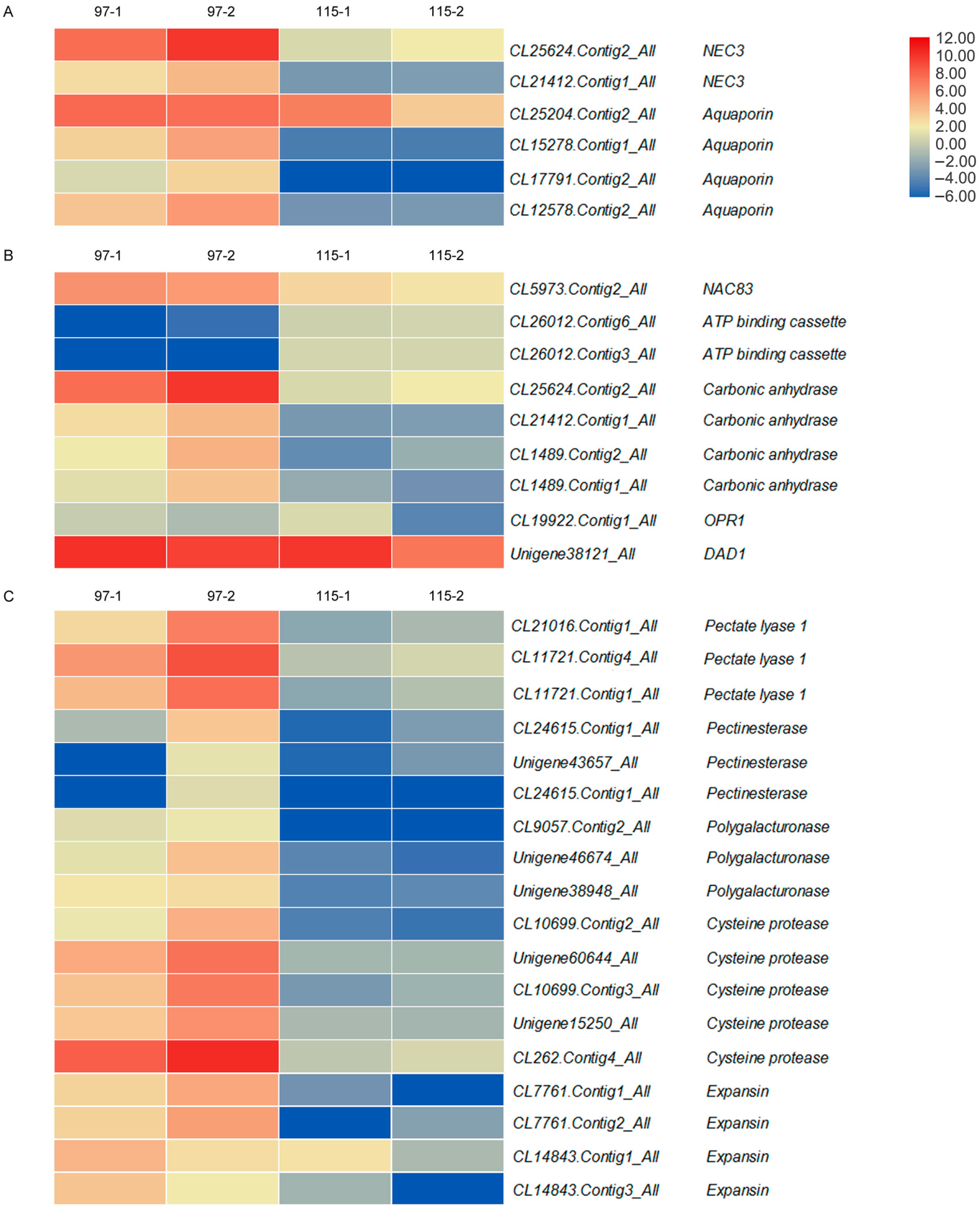
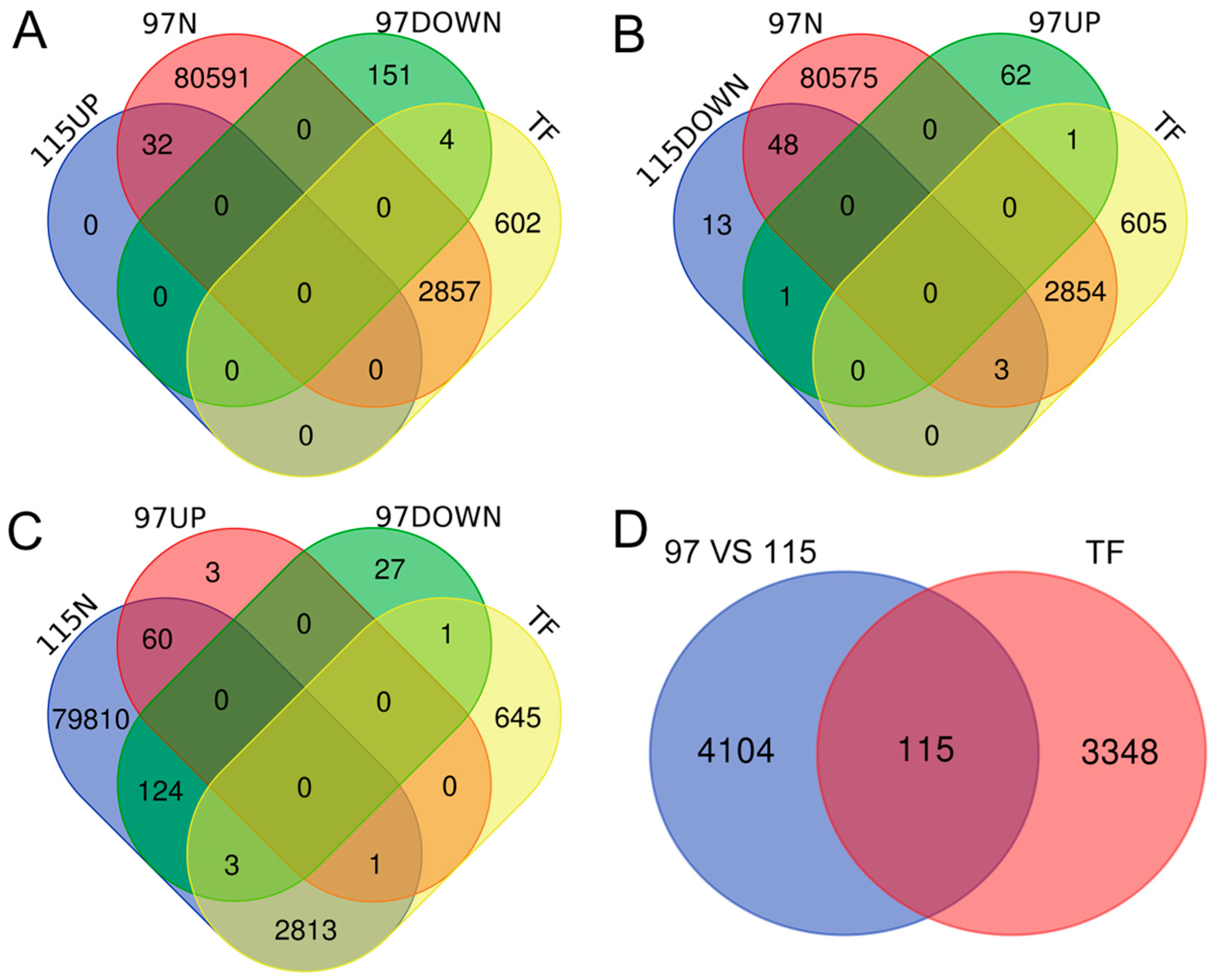
2.5. Anther Dehiscence-Associated DEGs
2.6. Six Selected Gens Expressions in Different Tissues
2.7. Comparisons of Accumulated Galacturonic Acid, Pectinase, Lignin, and Lignin Synthase in Anthers Between the Four Samples
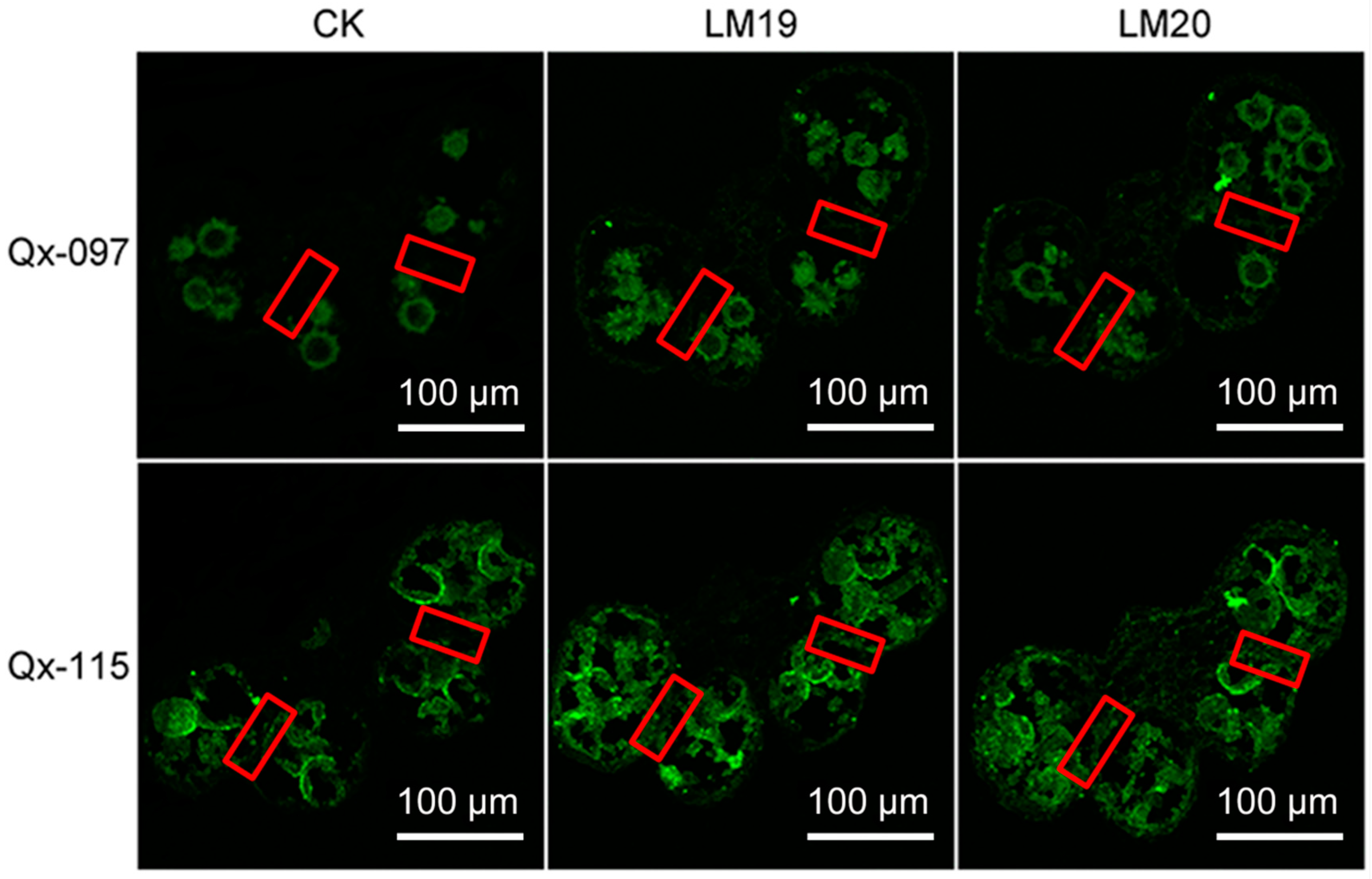
2.8. Pectin Accumulation in ‘Qx-097’ and ‘Qx-115’ Anthers
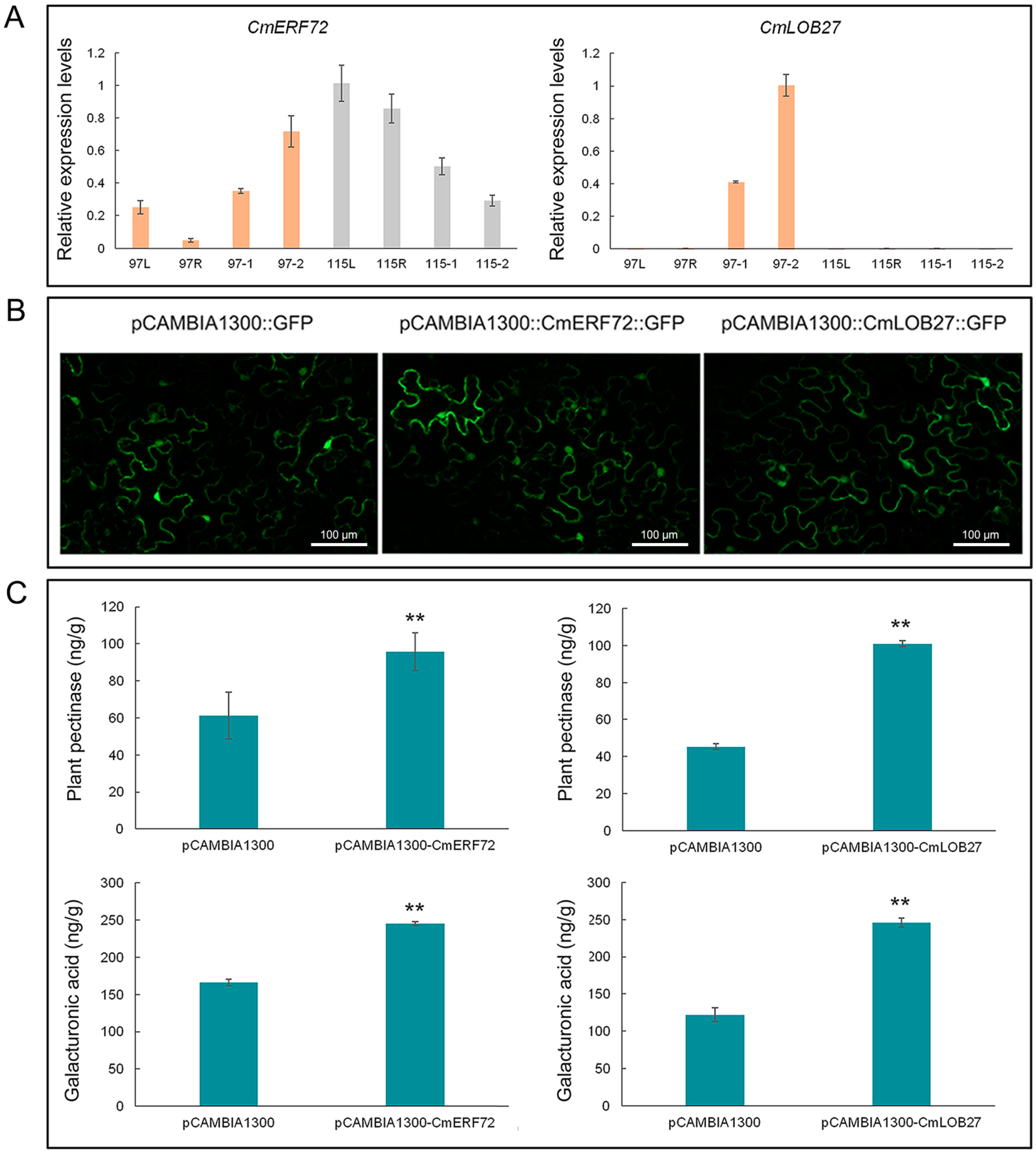
2.9. Expression Model of CmERF72 and CmLOB27 in Chrysanthemum
2.10. Transient Expression of CmERF72 and CmLOB27 in Tobacco Leaves
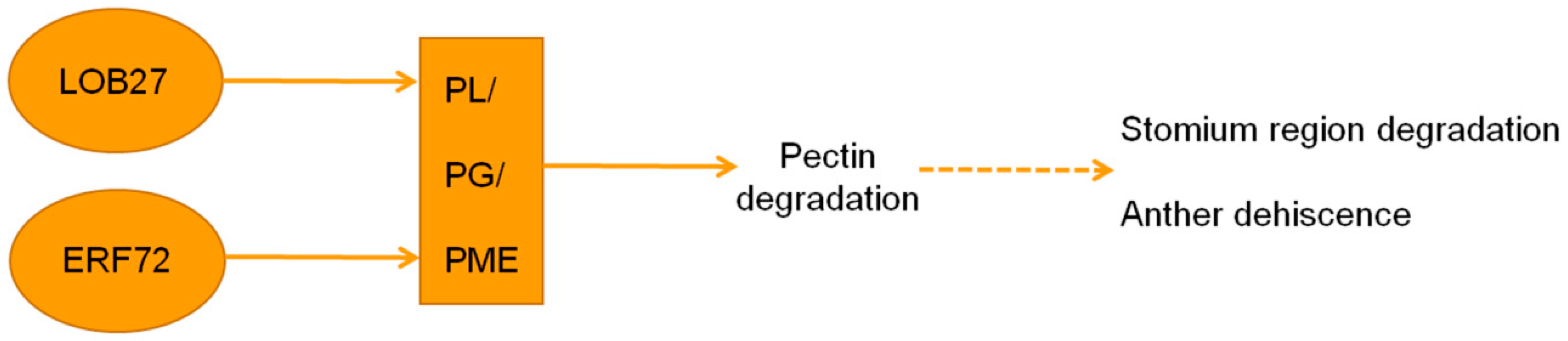
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Cytological Analysis of Anther Dehiscence
4.3. Transmission Electron Microscopy Analysis of Anther Dehiscence
4.4. Total RNA Extraction
4.5. cDNA Library Construction, Illumina Sequencing, and Data Analysis
4.6. De Novo Assembly, Sequence Clustering, Unigene TF Prediction, and Gene Function Annotation
4.7. Expression Analysis of Unigenes
4.8. Validation of Gene Expression Using Quantitative Real-Time PCR (qRT-PCR)
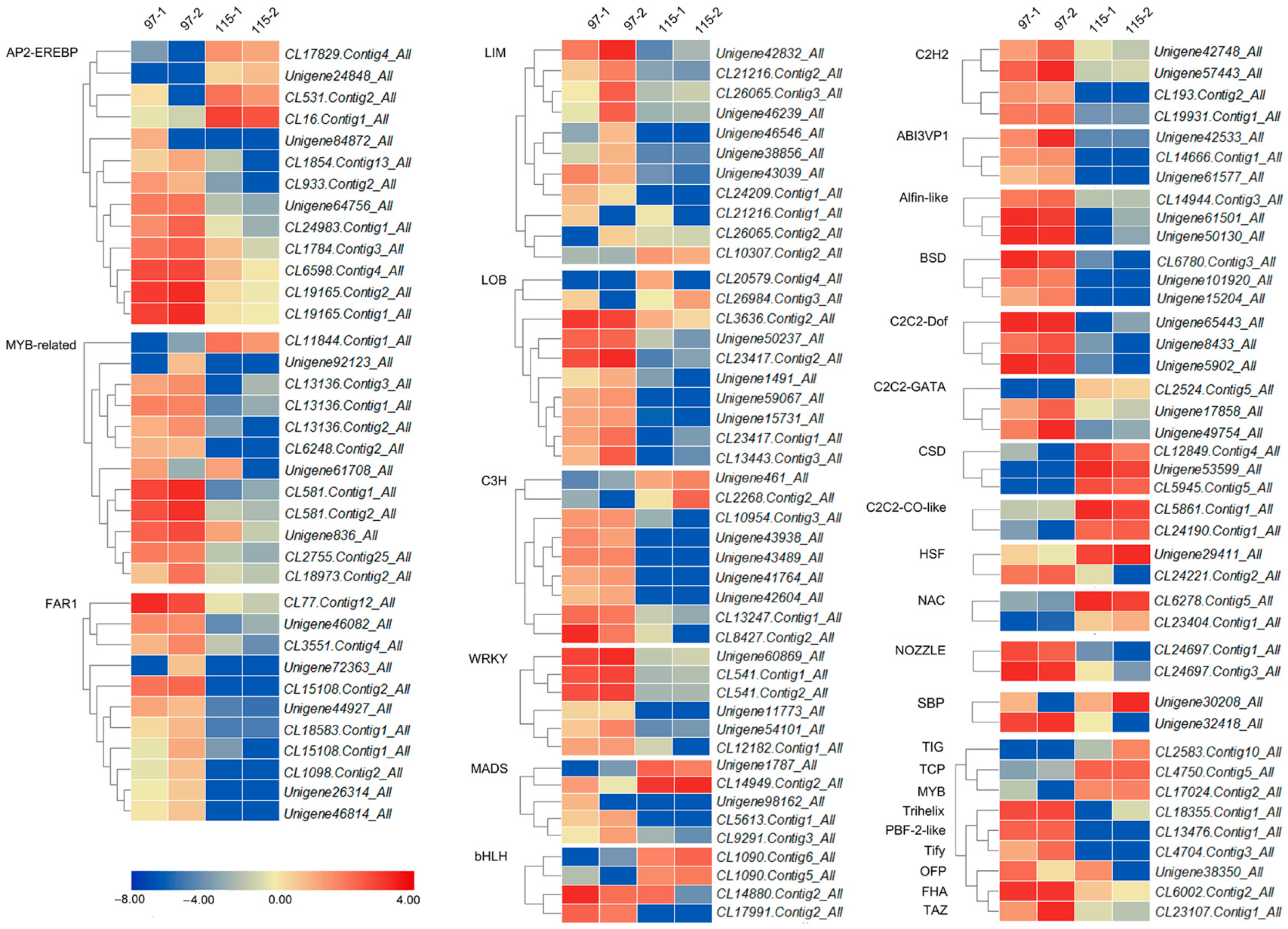
4.9. Screening of Differentially Expressed TFs
4.10. Determination of Galacturonic Acid, Pectinase, Lignin, and Lignin Synthase
4.11. Immunohistochemical Analysis
4.12. Isolation of CmERF72 and CmLOB27 and Construction of Expression Vectors
4.13. Transient Expression Analysis of CmERF72 and CmLOB27
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CA | Carbonic anhydrase |
| CP | Cysteine protease |
| DEG | Differentially expressed gene |
| EXP | Expansin |
| FAA | Formalin–acetic acid–alcohol |
| ORF | Open reading frame |
| PCA | Principal component analysis |
| PCD | Programmed cell death |
| PG | Polygalacturonase |
| PL | Pectate lyase |
| PME | Pectin methylesterase |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| SSR | Simple sequence repeat |
| TEM | Transmission electron microscope |
References
- Da Silva, J.A.T.; Kulus, D. Chrysanthemum biotechnology: Discoveries from the recent literature. Folia Hortic. 2014, 26, 67–77. [Google Scholar] [CrossRef]
- Lee, Y.W.; Choi, S.Y.; Lee, E.K.; Sohn, J.H.; Park, J.W.; Hong, C.S. Cross-allergenicity of pollens from the Compositae family: Artemisia vulgaris, Dendranthema grandiflorum, and Taraxacum officinale. Ann. Allergy 2007, 99, 526–533. [Google Scholar] [CrossRef]
- Groenewoud, G.C.; de Jong, N.W.; Burdorf, A.; de Groot, H.; van Wyk, R.G. Prevalence of occupational allergy to Chrysanthemum pollen in greenhouses in the Netherlands. Allergy 2002, 57, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, H.; Chen, F.; Jiang, J.; Fang, W.; Liao, Y.; Teng, N. Factors affecting quantity of pollen dispersal of spray cut chrysanthemum (Chrysanthemum morifolium). BMC Plant Biol. 2014, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Cecchetti, V.; Altamura, M.M.; Falasca, G.; Costantino, P.; Cardarelli, M. Auxin regulates Arabidopsis anther dehiscence, pollen maturation, and filament elongation. Plant Cell 2008, 20, 1760–1774. [Google Scholar] [CrossRef]
- Wilson, Z.A.; Song, J.; Taylor, B.; Yang, C. The final split: The regulation of anther dehiscence. J. Exp. Bot. 2011, 62, 1633–1649. [Google Scholar] [CrossRef]
- Rehman, S.; Yun, S.J. Developmental regulation of K accumulation in pollen, anthers, and papillae: Are anther dehiscence, papillae hydration, and pollen swelling leading to pollination and fertilization in barley (Hordeum vulgare L.) regulated by changes in K concentration? J. Exp. Bot. 2006, 57, 1315–1321. [Google Scholar] [CrossRef]
- Bots, M.; Feron, R.; Uehlein, N.; Weterings, K.; Kaidenhoff, R.; Mariani, T. PIP1 and PIP2 aquaporins are differentially expressed during tobacco anther and stigma development. J. Exp. Bot. 2005, 56, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, F.; Martin, V.; Colaneri, A.; Gonzalez-Schain, N.; Perales, M.; Martin, M.; Lombardo, C.; Braun, H.P.; Bartoli, C.; Zabaleta, E. Ectopic expression of mitochondrial gamma carbonic anhydrase 2 causes male sterility by anther indehiscence. Plant Mol. Biol. 2009, 70, 471–485. [Google Scholar] [CrossRef]
- Nelson, M.R.; Band, L.R.; Dyson, R.J.; Lessinnes, T.; Wells, D.M.; Yang, C.; Everitt, N.M.; Jensen, O.E.; Wilson, Z.A. A biomechanical model of anther opening reveals the roles of dehydration and secondary thickening. New Phytol. 2012, 196, 1030–1037. [Google Scholar] [CrossRef]
- He, H.; Bai, M.; Tong, P.; Hu, Y.; Yang, M.; Wu, H. CELLULASE6 and MANNANASE7 affect cell differentiation and silique dehiscence. Plant Physiol. 2018, 176, 2186–2201. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Kay, P.; Wilson, S.; Swain, S.M. ARABIDOPSIS DEHISCENCE ZONE POLYGALACTURONASE1 (ADPG1), ADPG2, and QUARTET2 are polygalacturonases required for cell separation during reproductive development in Arabidopsis. Plant Cell 2009, 21, 216–233. [Google Scholar] [CrossRef] [PubMed]
- Fei, J.; Tan, S.; Zhang, F.; Hua, L.; Liao, Y.; Fang, W.; Chen, F.; Teng, N. Morphological and physiological differences between dehiscent and indehiscent anthers of Chrysanthemum morifolium. J. Plant Res. 2016, 129, 1069–1082. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Hu, R.; Wang, H.; Cao, Y.; He, G.; Fu, C.; Zhou, G. MlWRKY12, a novel Miscanthus transcription factor, participates in pith secondary cell wall formation and promotes flowering. Plant Sci. 2013, 212, 1–9. [Google Scholar] [CrossRef]
- Zhong, R.; Lee, C.; Ye, Z. Global analysis of direct targets of secondary wall NAC master switches in Arabidopsis. Mol. Plant 2010, 3, 1087–1103. [Google Scholar] [CrossRef]
- Senatore, A.; Trobacher, C.P.; Greenwood, J.S. Ricinosomes predict programmed cell death leading to anther dehiscence in tomato. Plant Physiol. 2009, 149, 775–790. [Google Scholar] [CrossRef]
- Mitsuda, N.; Iwase, A.; Yamamoto, H.; Yoshida, M.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. NAC transcription factors, NST1 and NST3, are key regulators of the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell 2007, 19, 270–280. [Google Scholar] [CrossRef]
- Zhang, C.; Norris-Caneda, K.H.; Rottmann, W.H.; Gulledge, J.E.; Chang, S.; Kwan, B.Y.; Thomas, A.M.; Mandel, L.C.; Kothera, R.T.; Victor, A.D.; et al. Control of pollen-mediated gene flow in transgenic trees. Plant Physiol. 2012, 159, 1319–1334. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Thevenin, J.; Pollet, B.; Letarnec, B.; Saulnier, L.; Gissot, L.; Maia-Grondard, A.; Lapierre, C.; Jouanin, L. The simultaneous repression of CCR and CAD, two enzymes of the lignin biosynthetic pathway, results in sterility and dwarfism in Arabidopsis thaliana. Mol. Plant 2011, 4, 70–82. [Google Scholar] [CrossRef]
- Yang, C.; Xu, Z.; Song, J.; Conner, K.; Barrena, G.V.; Wilson, Z.A. Arabidopsis MYB26/MALE STERILE35 regulates secondary thickening in the endothecium and is essential for anther dehiscence. Plant Cell 2007, 19, 534–548. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, S.; Osakabe, Y.; Maruyama, K.; Ito, T.; Osakabe, K.; Sato, T.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Receptor-like protein kinase 2 (RPK 2) is a novel factor controlling anther development in Arabidopsis thaliana. Plant J. 2007, 50, 751–766. [Google Scholar] [CrossRef] [PubMed]
- Mitsuda, N.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. The NAC transcription factors NST1 and NST2 of Arabidopsis regulate secondary wall thickenings and are required for anther dehiscence. Plant Cell 2005, 17, 2993–3006. [Google Scholar] [CrossRef] [PubMed]
- Keijzer, C.J. The processes of anther dehiscence and pollen dispersal.II. The formation and the transfer mechanism of pollenkitt, cell-wall development of the loculus tissues and a function of orbicules in pollen dispersal. New Phytol. 1987, 105, 499. [Google Scholar] [CrossRef]
- Manning, J.C.; Goldblatt, P. Endothecium in iridaceae and its systematic implications. Am. J. Bot. 1990, 77, 527–532. [Google Scholar] [CrossRef] [PubMed]
- Jian-Fang, H.U.; Zhang, D.Y.; Gao, F.; Zhang, W. Anatomical structure of anther with abnormal dehiscence in ‘Jingbaili’ (Pyrus ussuriensis Maxim.) Pear. Acta Bot Bor-Occid Sin. 2009, 29, 1138–1143. [Google Scholar]
- Sharma, A.; Singh, M.B.; Bhalla, P.L. Anther ontogeny in Brachypodium distachyon. Protolasma 2015, 252, 439–450. [Google Scholar] [CrossRef]
- Yang, C.; Song, J.; Ferguson, A.C.; Klisch, D.; Simpson, K.; Mo, R.; Taylor, B.; Mitsuda, N.; Wilson, Z.A. Transcription factor MYB26 is key to spatial specificity in anther secondary thickening formation. Plant Physiol. 2017, 175, 333–350. [Google Scholar] [CrossRef]
- Ko, J.H.; Jeon, H.W.; Kim, W.C.; Kim, J.Y.; Han, K.H. The MYB46/MYB83-mediated transcriptional regulatory programme is a gatekeeper of secondary wall biosynthesis. Ann. Bot. 2014, 114, 1099–1107. [Google Scholar] [CrossRef]
- Weng, J.; Mo, H.; Chapple, C. Over-expression of F5H in COMT-deficient Arabidopsis leads to enrichment of an unusual lignin and disruption of pollen wall formation. Plant J. 2010, 64, 898–911. [Google Scholar] [CrossRef]
- Schilmiller, A.L.; Stout, J.; Weng, J.; Humphreys, J.; Ruegger, M.O.; Chapple, C. Mutations in the cinnamate 4-hydroxylase gene impact metabolism, growth and development in Arabidopsis. Plant J. 2009, 60, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Rieu, I.; Wolters-Arts, M.; Derksen, J.; Mariani, C.; Weterings, K. Ethylene regulates the timing of anther dehiscence in tobacco. Planta 2003, 217, 131–137. [Google Scholar] [PubMed]
- Cosgrove, D.J. Loosening of plant cell walls by expansins. Nature 2000, 407, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.P.; Nguema-Ona, E.E.; Vicre-Gibouin, M.; Sorensen, I.; Willats, W.G.; Driouich, A.; Farrant, J.M. Arabinose-rich polymers as an evolutionary strategy to plasticize resurrection plant cell walls against desiccation. Planta 2013, 237, 739–754. [Google Scholar] [CrossRef]
- Jiang, F.; Lopez, A.; Jeon, S.; de Freitas, S.T.; Yu, Q.; Wu, Z.; Labavitch, J.M.; Tian, S.; Powell, A.L.T.; Mitcham, E. Disassembly of the fruit cell wall by the ripening-associated polygalacturonase and expansin influences tomato cracking. Hortic. Res. 2019, 6. [Google Scholar] [CrossRef] [Green Version]
- Chea, S.; Yu, D.J.; Park, J.; Oh, H.D.; Chung, S.W.; Lee, H.J. Fruit softening correlates with enzymatic and compositional changes in fruit cell wall during ripening in ‘Bluecrop’ highbush blueberries. Sci. Hortic. 2019, 245, 163–170. [Google Scholar] [CrossRef]
- Wang, D.; Yeats, T.H.; Uluisik, S.; Rose, J.K.C.; Seymour, G.B. Fruit softening: Revisiting the role of pectin. Trends Plant Sci. 2018, 23, 302–310. [Google Scholar] [CrossRef]
- Corral-Martinez, P.; Garcia-Fortea, E.; Bernard, S.; Driouich, A.; Segui-Simarro, J.M. Ultrastructural immunolocalization of arabinogalactan protein, pectin and hemicellulose epitopes through anther development in Brassica napus. Plant Cell Physiol. 2016, 57, 2161–2174. [Google Scholar] [CrossRef] [Green Version]
- Husbands, A.; Bell, E.M.; Shuai, B.; Smith, H.M.S.; Springer, P.S. LATERAL ORGAN BOUNDARIES defines a new family of DNA-binding transcription factors and can interact with specific bHLH proteins. Nucleic Acids Res. 2007, 35, 6663–6671. [Google Scholar] [CrossRef]
- Xu, C.; Luo, F.; Hochholdinger, F. LOB domain proteins: Beyond lateral organ boundaries. Trends Plant Sci. 2016, 21, 159–167. [Google Scholar] [CrossRef]
- Ba, L.; Shan, W.; Kuang, J.; Feng, B.; Xiao, Y.; Lu, W.; Chen, J. The banana MaLBD (LATERAL ORGAN BOUNDARIES DOMAIN) transcription factors regulate EXPANSIN expression and are involved in fruit ripening. Plant Mol. Biol. Rep. 2014, 32, 1103–1113. [Google Scholar] [CrossRef]
- Palmerossuárez, P.A.; Massangesánchez, J.A.; Sánchezsegura, L.; Martínezgallardo, N.A.; Espitia, R.E.; Gómezleyva, J.F.; Délanofrier, J.P. AhDGR2, an amaranth abiotic stress-induced DUF642 protein gene, modifies cell wall structure and composition and causes salt and ABA hyper-sensibility in transgenic Arabidopsis. Planta 2016, 245, 1–18. [Google Scholar]
- Li, M.; Zhang, Y.; Zhang, Z.; Ji, X.; Zhang, R.; Liu, D.; Gao, L.; Zhang, J.; Wang, B.; Wu, Y.; et al. Hypersensitive ethylene signaling and ZMdPG1 expression lead to fruit softening and dehiscence. PLoS ONE 2013, 8, e58745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Liu, Y.; Liang, Y.; Lu, J.; Jiang, C.; Fei, Z.; Jiang, C.Z.; Ma, C.; Gao, J. Rosa hybrida RhERF1 and RhERF4 mediate ethylene- and auxin-regulated petal abscission by influencing pectin degradation. Plant J. 2019, 99, 1159–1171. [Google Scholar] [CrossRef]
- Teng, N.; Wang, Y.; Sun, C.; Fang, W.; Chen, F. Factors influencing fecundity in experimental crosses of water lotus (Nelumbo nucifera Gaertn.) cultivars. BMC Plant Biol. 2012, 12, 82. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Huang, Z.; Wang, Y.; Chen, F.; Teng, N.; Fang, W.; Liu, Z. Overcoming pre-fertilization barriers in the wide cross between Chrysanthemum grandiflorum (Ramat.) Kitamura and C. nankingense (Nakai) Tzvel. by using special pollination techniques. Euphytica 2011, 178, 195–202. [Google Scholar] [CrossRef]
- Cock, P.J.; Fields, C.J.; Goto, N.; Heuer, M.L.; Rice, P.M. The Sanger FASTQ file format for sequences with quality scores, and the Solexa/Illumina FASTQ variants. Nucleic Acids Res. 2010, 38, 1767–1771. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. TIGR Gene Indices clustering tools (TGICL): A software system for fast clustering of large EST datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [Green Version]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41, e121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33, W116–W120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Tarazona, S.; Garcia-Alcalde, F.; Dopazo, J.; Ferrer, A.; Conesa, A. Differential expression in RNA-seq: A matter of depth. Genome Res. 2011, 21, 2213–2223. [Google Scholar] [CrossRef] [Green Version]
- Song, A.; Lu, J.; Jiang, J.; Chen, S.; Guan, Z.; Fang, W.; Chen, F. Isolation and characterisation of Chrysanthemum crassum SOS1, encoding a putative plasma membrane Na+/H+ antiporter. Plant Biol. 2012, 14, 706–713. [Google Scholar] [CrossRef]
- Wu, Z.; Liang, J.; Zhang, S.; Zhang, B.; Zhao, Q.; Li, G.; Yang, X.; Wang, C.; He, J.; Yi, M. A canonical DREB2-type transcription factor in lily is post-translationally regulated and mediates heat stress response. Front. Plant Sci. 2018, 9, 243. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Values | Total | Nr | Nt | Swiss-Prot | KEGG | COG | Interpro | GO |
|---|---|---|---|---|---|---|---|---|
| Number | 213,845 | 115,949 | 95,803 | 79,969 | 87,073 | 41,570 | 76,348 | 37,081 |
| Percentage (%) | 100 | 54.22 | 44.08 | 37.40 | 40.72 | 19.44 | 35.70 | 17.34 |
| Samples. | Galacturonic Acid (ng/g) | Pectinase (ng/g) | Lignin (ng/g) | LNS (ng/g) |
|---|---|---|---|---|
| Qx-097-1 | 113.298 ± 5.389 b | 117.105 ± 2.093 a | 387.261 ± 7.674 c | 528.150 ± 31.466 c |
| Qx-097-2 | 83.651 ± 4.791 c | 107.070 ± 4.063 b | 449.010 ± 18.205 a | 670.039 ± 39.355 a |
| Qx-115-1 | 137.171 ± 4.533 a | 69.677 ± 0.934 d | 415.938 ± 4.386 b | 591.352 ± 19.321 b |
| Qx-115-2 | 139.494 ± 1.983 a | 94.747 ± 3.134 c | 462.407 ± 16.761 a | 695.570 ± 44.867 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Wu, Z.; Wu, H.; Fang, W.; Chen, F.; Teng, N. Transcriptome Profiling Unravels a Vital Role of Pectin and Pectinase in Anther Dehiscence in Chrysanthemum. Int. J. Mol. Sci. 2019, 20, 5865. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235865
Li Q, Wu Z, Wu H, Fang W, Chen F, Teng N. Transcriptome Profiling Unravels a Vital Role of Pectin and Pectinase in Anther Dehiscence in Chrysanthemum. International Journal of Molecular Sciences. 2019; 20(23):5865. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235865
Chicago/Turabian StyleLi, Qian, Ze Wu, Huijun Wu, Weimin Fang, Fadi Chen, and Nianjun Teng. 2019. "Transcriptome Profiling Unravels a Vital Role of Pectin and Pectinase in Anther Dehiscence in Chrysanthemum" International Journal of Molecular Sciences 20, no. 23: 5865. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235865