Endomorphin-2- and Neurotensin- Based Chimeric Peptide Attenuates Airway Inflammation in Mouse Model of Nonallergic Asthma

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
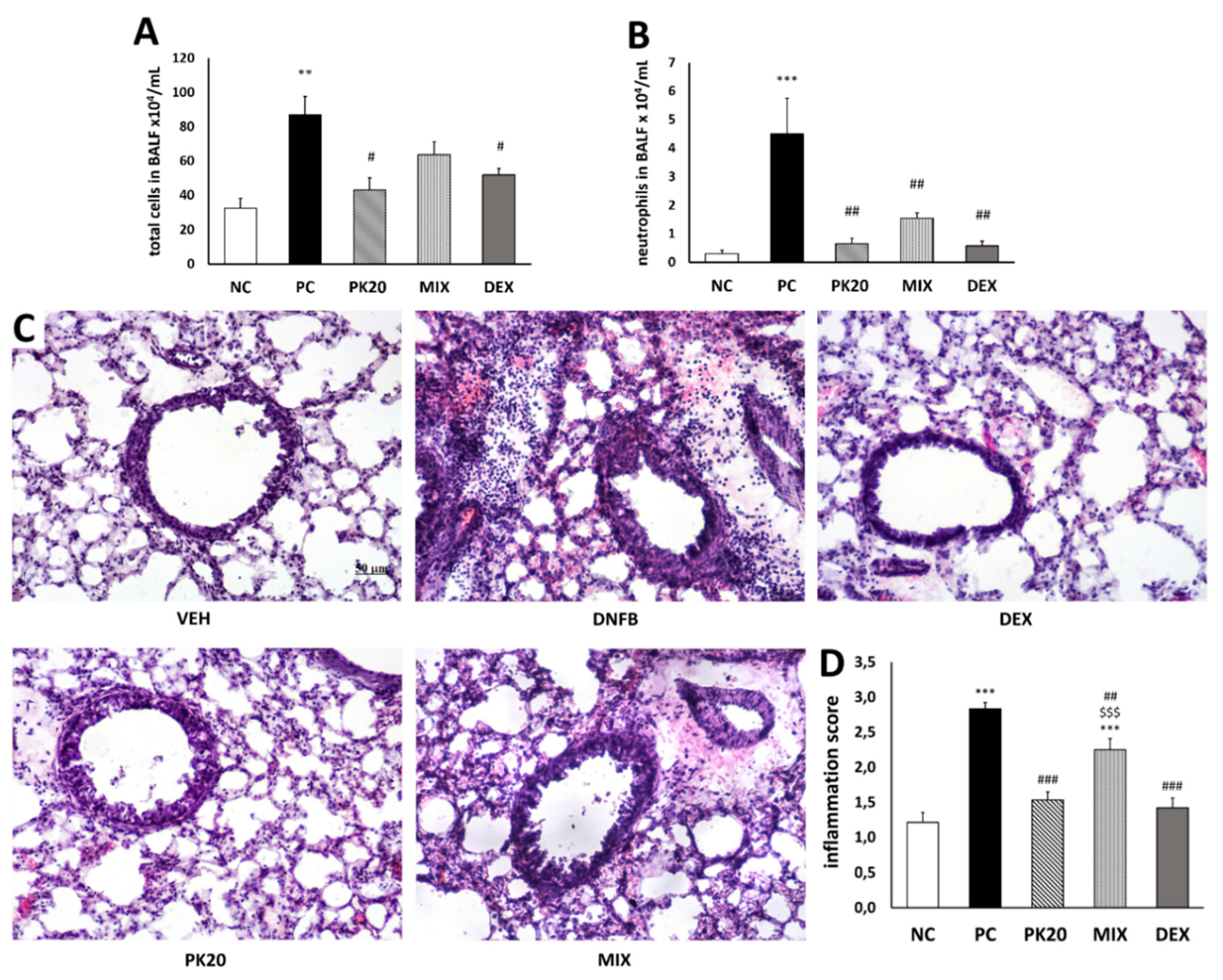
2.1. Effect of PK20 on DNFB-Induced Inflammatory Cell Infiltration in BALF and Lung Tissue
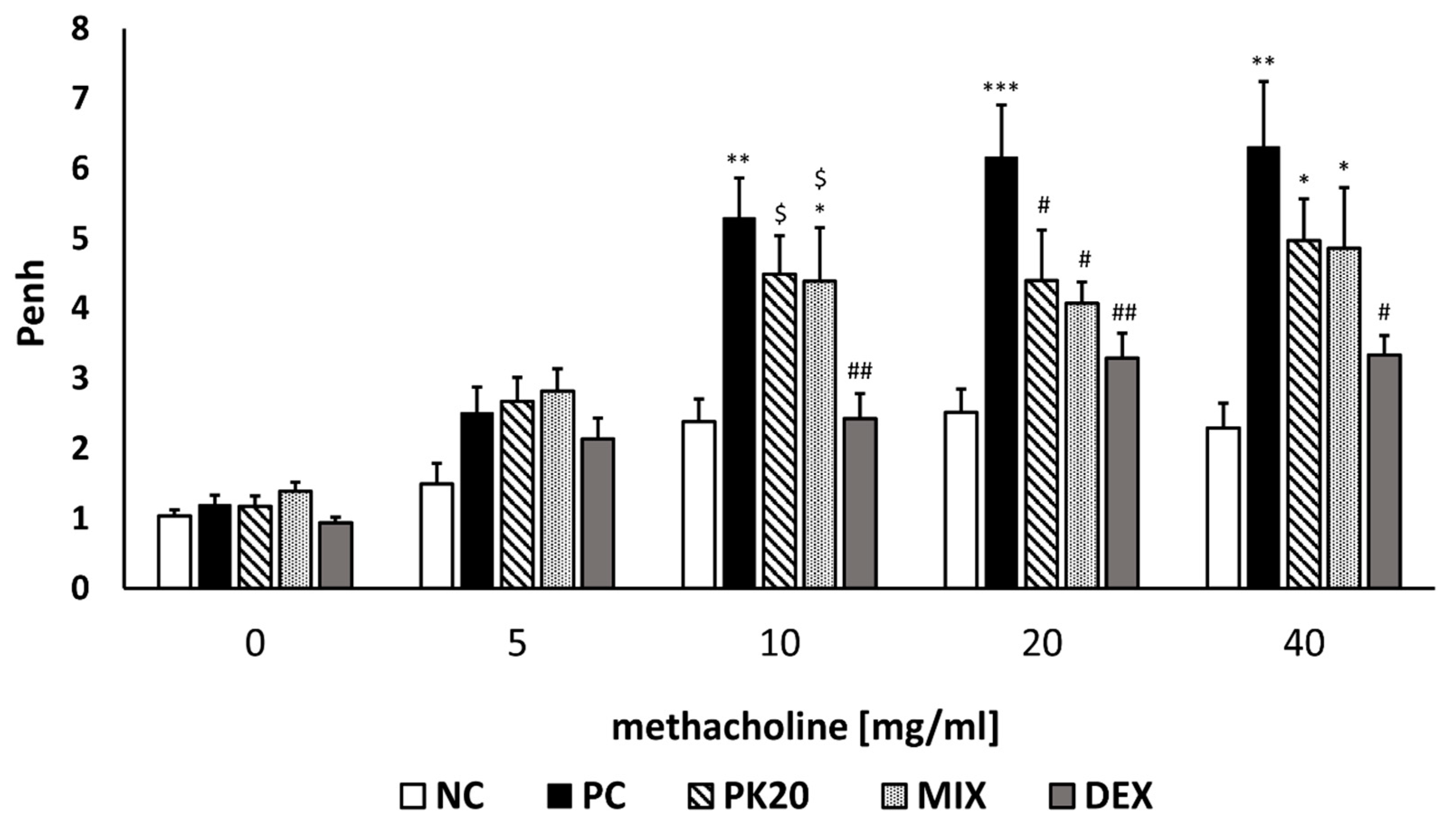
2.2. PK20 Reduces Airway Hyperresponsiveness (AHR)
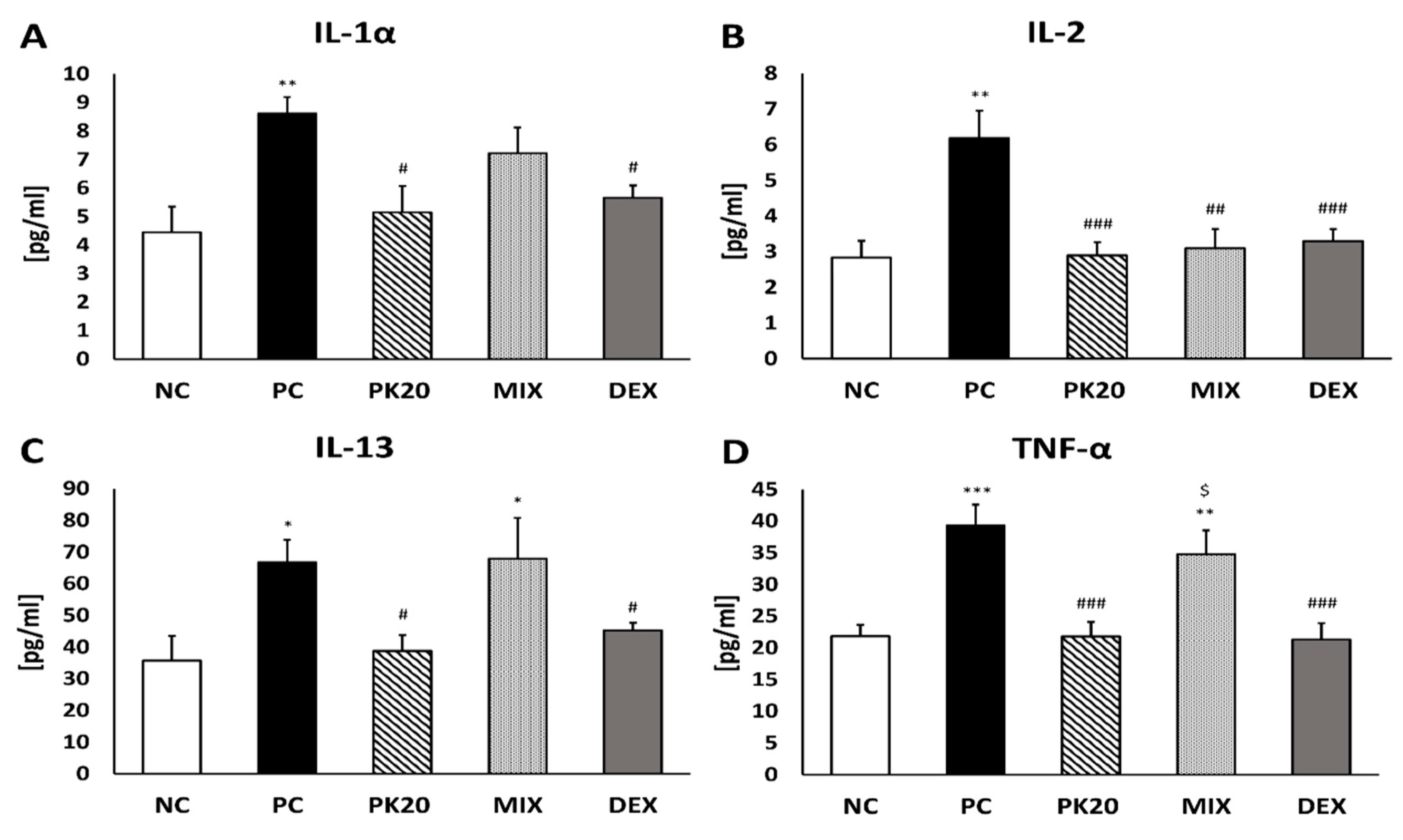
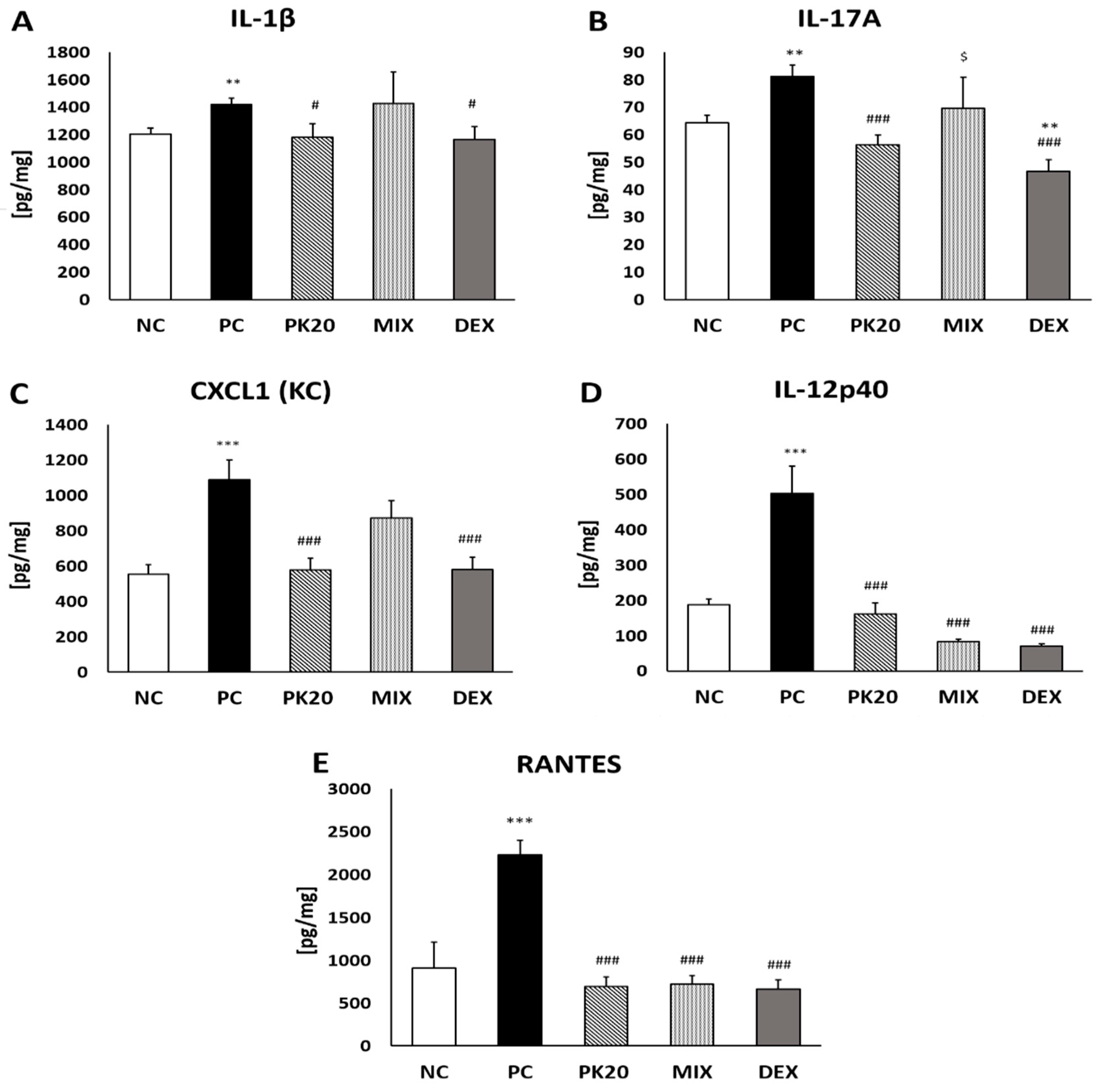
2.3. Effect of PK20 on DNFB-Induced Pro-Inflammatory Cytokine and Chemokine Production
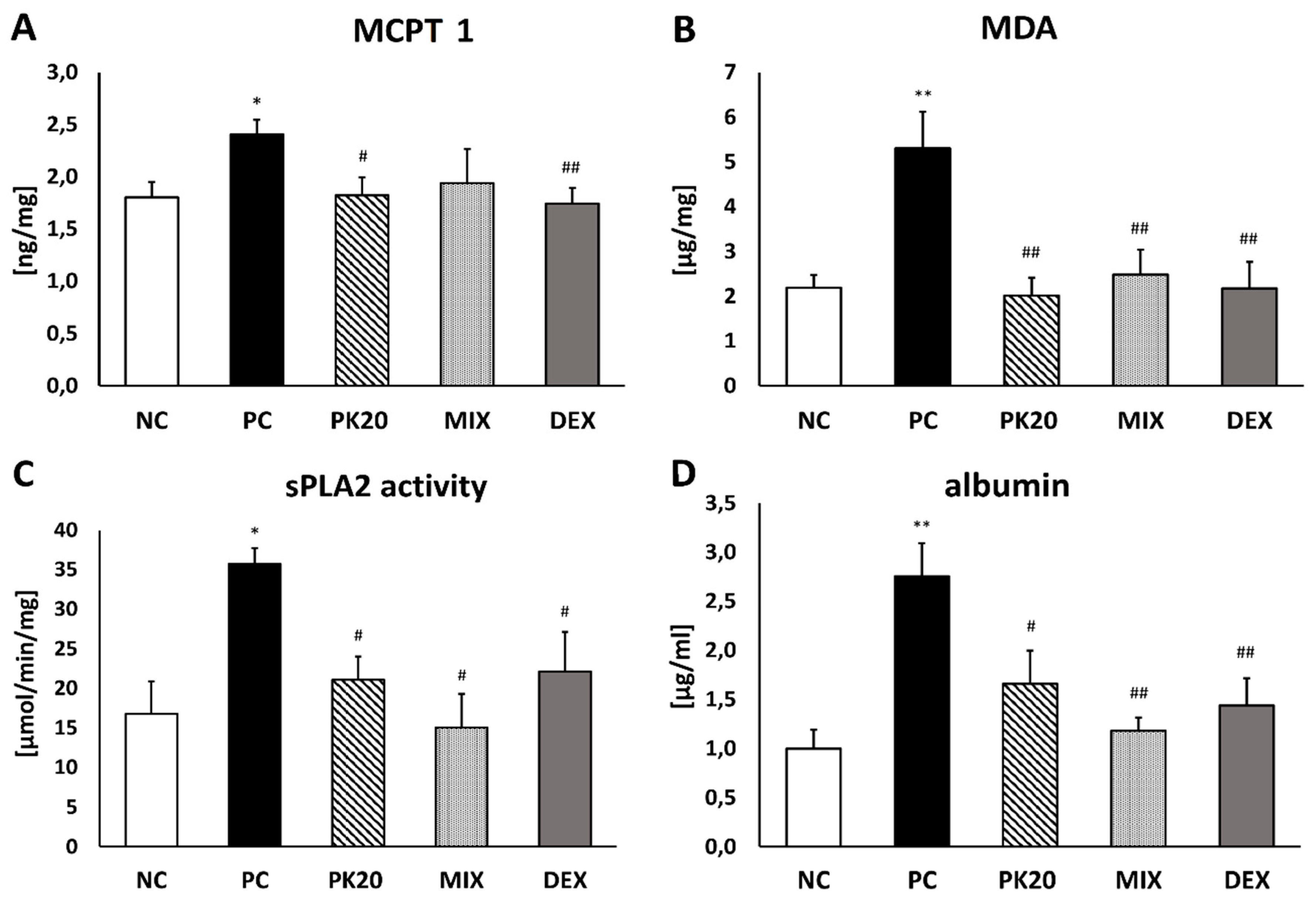
2.4. Effect of PK20 Treatment on Mouse Mast Cell Protease (MCPT 1) Level in Lungs
2.5. Effect of PK20 Treatment on Malondialdehyde Level (MDA) in Lungs
2.6. Effect of PK20 Treatment on Activity of Secreted Phospholipase A2 (sPLA2) in Lungs
2.7. Effect of PK20 Treatment on Albumin Concentration in BALF
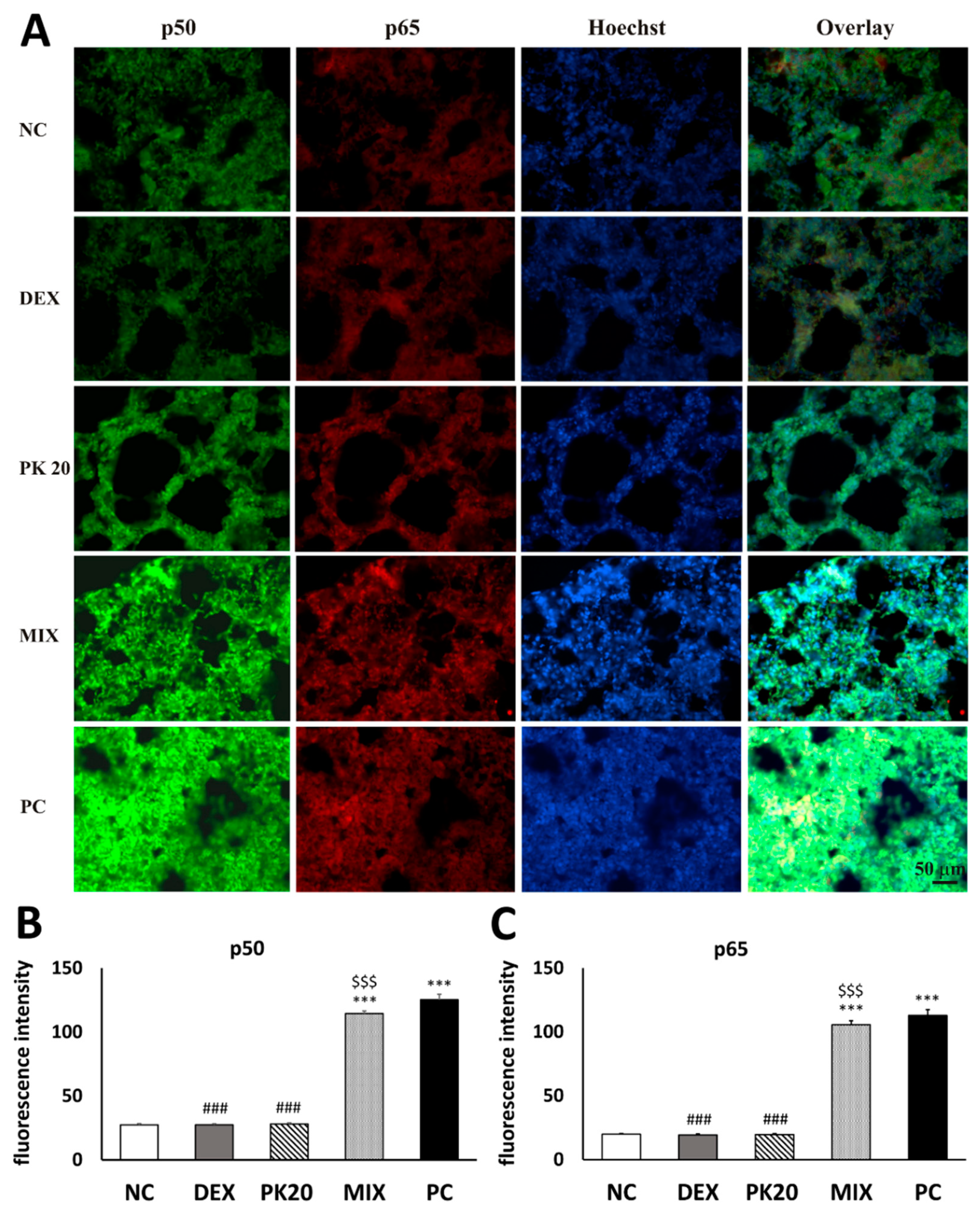
2.8. Effect of PK20 on Lung Nuclear Factor Kappa B (NF-κB) Immunohistochemistry
3. Discussion
4. Materials and Methods
4.1. Drugs and Reagents
4.2. Animals
4.3. DNFB-Induced Experimental Asthma
4.4. Experimental Groups
4.5. Measurement of Airway Hyperresponsiveness
4.6. Total and Differential Cell Counts in Bronchoalveolar Lavage Fluid
4.7. Cytokine and Chemokine Quantification
4.8. Measurement of Oxidative Stress (Malondialdehyde Level) in Lung Tissue
4.9. Measurement of Mouse Mast Cell Protease Level (MCPT 1) in Lung Tissue
4.10. Measurement of Secretory Phospholipase 2 (PLA2) Activity
4.11. Determination of Total Protein and Albumin Concentration
4.12. Lung Histology
4.13. NF-κB Immunohistochemistry
4.14. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Holgate, S.T.; Arshad, H.S.; Roberts, G.C.; Howarth, P.H.; Thurner, P.; Davies, D.E. A new look at the pathogenesis of asthma. Clin. Sci. 2009, 118, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Pelaia, G.; Vatrella, A.; Busceti, M.T.; Gallelli, L.; Calabrese, C.; Terracciano, R.; Maselli, R. Cellular mechanisms underlying eosinophilic and neutrophilic airway inflammation in asthma. Mediat. Inflamm. 2015, 2015, 8. [Google Scholar] [CrossRef] [PubMed]
- Global Asthma Report, Global Asthma Network, Auckland, New Zealand. Available online: http://www.globalasthmanetwork.org/publications/Global_Asthma_Report_2014.pdf,2014 (accessed on 9 September 2016).
- Domínguez-Ortega, J.; Phillips-Anglés, E.; Barranco, P.; Quirce, S. Cost-effectiveness of asthma therapy: A comprehensive review. J. Asthma 2015, 52, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Behera, D.; Sehgal, I.S. Bronchial asthma-issues for the developing world. Indian J. Med. 2015, 141, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.; Lim, J.C.; Soo, H.L.; Stanslas, J. Molecularly targeted therapies for asthma: Current development, challenges and potential clinical translation. Pulm. Pharmacol. Ther. 2016, 40, 52–68. [Google Scholar] [CrossRef]
- Abramson, M.J.; Walters, J.; Walters, E.H. Adverse effects of β-agonists. Am. J. Respir. Med. 2003, 2, 287–297. [Google Scholar] [CrossRef]
- Chung, K.F.; Wenzel, S.E.; Brozek, J.L.; Bush, A.; Castro, M.; Sterk, P.J.; Teague, W.G. International ERS/ATS guidelines on definition, evaluation and treatment of severe asthma. Eur. Respir. J. 2014, 43, 343–373. [Google Scholar] [CrossRef]
- Singh, J.; Shah, R.; Singh, D. Inundation of asthma target research: Untangling asthma riddles. Pulm. Pharmacol. Ther. 2016, 41, 60–85. [Google Scholar] [CrossRef]
- Morphy, R.; Rankovic, Z. Fragments, network biology and designing multiple ligands. Drug Discov. Today 2007, 12, 156–160. [Google Scholar] [CrossRef]
- Zimmermann, G.R.; Lehár, J.; Keith, C.T. Multi-target therapeutics: When the whole is greater than the sum of the parts. Drug Discov. Today 2007, 12, 34–42. [Google Scholar] [CrossRef]
- Kleczkowska, P.; Kowalczyk, A.; Lesniak, A.; Bujalska-Zadrozny, M. The discovery and development of drug combinations for the treatment of various diseases from patent literature (1980-Present). Curr. Top. Med. Chem. 2017, 17, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowska, P.; Kosson, P.; Ballet, S.; Van den Eynde, I.; Tsuda, Y.; Tourwe, D.; Lipkowski, A.W. PK20, a new opioid-neurotensin hybrid peptide that exhibits central and peripheral antinociceptive effects. Mol. Pain 2010, 6, 86. [Google Scholar] [CrossRef] [PubMed]
- Kleczkowska, P.; Kawalec, M.; Bujalska-Zadrozny, M.; Filip, M.; Zabłocka, B.; Lipkowski, A.W. Effects of the hybridization of opioid and neurotensin pharmacophores on cell survival in rat organotypic hippocampal slice cultures. Neurotox. Res. 2015, 28, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Kaczyńska, K.; Kogut, E.; Zając, D.; Jampolska, M.; Andrzejewski, K.; Sulejczak, D.; Lipkowski, A.; Kleczkowska, P. Neurotensin-based hybrid peptide’s anti-inflammatory activity in murine model of a contact sensitivity response. Eur. J. Pharm. Sci. 2016, 93, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Akcan, A.; Muhtaroglu, S.; Akgun, H.; Akyildiz, H.; Kucuk, C.; Sozuer, E.; Yurci, A.; Yilmaz, N. Ameliorative effects of bombesin and neurotensin on trinitrobenzene sulphonic acid-induced colitis, oxidative damage and apoptosis in rats. World J. Gastroenterol. 2008, 14, 1222–1230. [Google Scholar] [CrossRef]
- Russjan, E.; Kaczyńska, K. Beneficial effects of neurotensin in murine model of hapten-induced asthma. Int. J. Mol. Sci. 2019, 20, 5025. [Google Scholar] [CrossRef]
- Stein, C.; Kuchler, S. Targeting inflammation and wound healing by opioids. Trends Pharmcol. Sci. 2013, 34, 303–312. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, X.; Cui, Y.; Fan, Y.Z.; Liu, J.; Wang, R. Abnormal modulation of cholinergic neurotransmission by endomorphin 1 and endomorphin 2 in isolated bronchus of type 1 diabetic rats. Peptides 2006, 27, 2770–2777. [Google Scholar] [CrossRef]
- Groneberg, D.A.; Fischer, A. Endogenous opioids as mediators of asthma. Pulm. Pharmacol. Ther. 2001, 14, 383–389. [Google Scholar] [CrossRef]
- Börzsei, R.; Pozsgai, G.; Bagoly, T.; Elekes, K.; Pintér, E.; Szolcsányi, J.; Helyes, Z. Inhibitory action of endomorphin-1 on sensory neuropeptide release and neurogenic inflammation in rats and mice. Neuroscience 2008, 152, 82–88. [Google Scholar] [CrossRef]
- Van der Kleij, H.P.; Kraneveld, A.D.; Van Houwelingen, A.H.; Kool, M.; Weitenberg, A.C.; Redegeld, F.A.; Nijkamp, F.P. Murine model for non-IgE-mediated asthma. Inflammation 2004, 28, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, T.E.; Kaya, Y.; Durlu-Kandilci, N.T.; Onder, S.; Sahin-Erdem, L.I. The effect of cannabinoids on dinitrofluorobenzene-induced experimental asthma in mice. Respir. Physiol. Neurobiol. 2016, 231, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Russjan, E.; Kaczyńska, K. Murine models of hapten-induced asthma. Toxicology 2018, 410, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. The cytokine network in asthma and chronic obstructive pulmonary disease. J. Clin. Investig. 2008, 118, 3546–3556. [Google Scholar] [CrossRef] [PubMed]
- Smart, S.J.; Casale, T.B. Pulmonary epithelial cells facilitate TNF-alpha-induced neutrophil chemotaxis. A role for cytokine networking. J. Immunol. 1994, 152, 4087–4094. [Google Scholar]
- Joosten, L.A.; Netea, M.G.; Dinarello, C.A. Interleukin-1β in innate inflammation, autophagy and immunity. Semin. Immunol. 2013, 25, 416–424. [Google Scholar] [CrossRef]
- Nabe, T. Tumor necrosis factor alpha-mediated asthma? Int. Arch. Allergy Immunol. 2013, 160, 111–113. [Google Scholar] [CrossRef]
- Tsukagoshi, H.; Sakamoto, T.; Xu, W.; Barnes, P.; Chung, F. Effect of interleukin-1 on airway hyperresponsiveness and inflammation in sensitized and nonsensitized Brown-Norway rats. J. Allergy Clin. Immunol. 1994, 93, 464–469. [Google Scholar] [CrossRef]
- Johnson, V.J.; Yucesoy, B.; Luster, M.I. Prevention of IL-1 signaling attenuates airway hyperresponsiveness and inflammation in a murine model of toluene diisocyanate–induced asthma. J. Allergy Clin. Immunol. 2005, 116, 851–858. [Google Scholar] [CrossRef]
- Kips, J.C.; Tavernier, J.H.; Joos, G.F.; Peleman, R.A.; Pauwels, R.A. The potential role of tumor necrosis factor α in asthma. Clin. Exp. Allergy 1993, 23, 247–250. [Google Scholar] [CrossRef]
- Kim, H.K.; Lee, C.H.; Kim, J.M.; Ayush, O.; Im, S.Y.; Lee, H.K. Biphasic late airway hyperresponsiveness in a murine model of asthma. Int. Arch. Allergy Immunol. 2012, 160, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Barczyk, A.; Pierzchala, W.; Sozanska, E. Interleukin-17 in sputum correlates with airway hyperresponsiveness to methacholine. Respir. Med. 2003, 97, 726–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newcomb, D.C.; Peebles, R.S. Th17-mediated inflammation in asthma. Curr. Opin. Immunol. 2013, 25, 755–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcorn, J.F.; Crowe, C.R.; Kolls, J.K. Th17 cells in asthma and COPD. Annu. Rev. Physiol. 2010, 72, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Sawant, K.V.; Poluri, K.M.; Dutta, A.K.; Sepuru, K.M.; Troshkina, A.; Garofalo, R.P.; Rajarathnam, K. Chemokine CXCL1 mediated neutrophil recruitment: Role of glycosaminoglycan interactions. Sci. Rep. 2016, 6, 33123. [Google Scholar] [CrossRef] [Green Version]
- Rajarathnam, K.; Schnoor, M.; Richardson, R.M.; Rajagopal, S. How do chemokines navigate neutrophils to the target site: Dissecting the structural mechanisms and signaling pathways. Cell. Signal. 2019, 54, 69–80. [Google Scholar] [CrossRef]
- Pan, Z.Z.; Parkyn, L.; Ray, A.; Ray, P. Inducible lung-specific expression of RANTES: Preferential recruitment of neutrophils. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, 658–666. [Google Scholar] [CrossRef]
- Walker, C.; Bode, E.; Boer, L.; Hansel, T.T.; Blaser, K.; Virchow, J. Allergic and nonallergic asthmatics have distinct patterns of T-cell activation and cytokine production in peripheral blood and bronchoalveolar lavage. Am. Rev. Repir. Dis. 1992, 146, 109–115. [Google Scholar] [CrossRef]
- Leung, D.Y.; Martin, R.J.; Szefler, S.J.; Sher, E.R.; Ying, S.; Kay, A.B.; Hamid, Q. Dysregulation of interleukin-4, interleukin-5 and interferon-γ gene expression in steroid resistant asthma. J. Exp. Med. 1995, 181, 33–40. [Google Scholar] [CrossRef]
- Walter, M.J.; Kajiwara, N.; Karanja, P.; Castro, M.; Holtzman, M.J. Interleukin 12p40 production by barrier epithelialc cells during airway inflammation. J. Exp. Med. 2001, 193, 339–351. [Google Scholar] [CrossRef]
- Grünig, G.; Warnock, M.; Wakil, A.E.; Venkayya, R.; Brombacher, F.; Rennick, D.M.; Sheppard, D.; Mohrs, M.; Donaldson, D.D.; Locksley, R.M.; et al. Requirement for IL-13 independently of IL-4 in experimental asthma. Science 1998, 282, 2261–2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nader, M.A. Inhibition of airway inflammation and remodeling by sitagliptin in murine chronic asthma. Int. Immunopharmacol. 2015, 29, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Wills-Karp, M.; Luyimbazi, J.; Xu, X.; Schofield, B.; Neben, T.Y.; Karp, C.L.; Donaldson, D.D. Interleukin-13: Central mediator of allergic asthma. Science 1998, 282, 2258–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humbert, M.; Dursham, S.R.; Kimmitt, P.; Powell, N.; Assoufi, B.; Pfister, R.; Menz, G.; Kay, A.B.; Corrigan, C.J. Elevated expression of messenger ribonucleic acid encoding IL-13 in the bronchial mucosa of atopic and nonatopic subjects with asthma. J. Allergy Clin. Immunol. 1997, 99, 657–665. [Google Scholar] [CrossRef]
- Zuo, L.; Otenbaker, N.P.; Rose, B.A.; Salisbury, K.S. Molecular mechanisms of reactive oxygen species-related pulmonary inflammation and asthma. Mol. Immunol. 2013, 56, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Marcal, L.E.; Rehder, J.; Newburger, P.E.; Condino-Neto, A. Superoxide release and cellular gluthatione peroxidase activity in leukocytes from children with persistent asthma. Braz. J. Med. Biol. Res. 2004, 37, 1607–1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeem, A.; Siddiqui, N.; Alharbi, N.O.; Alharbi, M.M. Airway and systemic oxidant-antioxidant dysregulation in asthma: A possible scenario of oxidants spill over from lung into blood. Pulm. Pharmacol. Ther. 2014, 29, 31–40. [Google Scholar] [CrossRef]
- Joseph, B.Z.; Routes, J.M.; Borish, L. Activities of superoxide dismutases and NADPH oxidase in neutrophils obtained from asthmatic and normal donors. Inflammation 1993, 17, 361–370. [Google Scholar] [CrossRef]
- Chihara, J.; Yamada, H.; Yamamoto, T.; Kurachi, D.; Hayashi-Kameda, N.; Honda, K.; Kayaba, H.; Urayama, O. Priming effect of RANTES on eosinophil oxidative metabolism. Allergy 1998, 53, 1178–1182. [Google Scholar] [CrossRef]
- Hattori, H.; Imai, H.; Furuhama, K.; Sato, O.; Nakagawa, Y. Induction of phospholipid hydroperoxide glutathione peroxidase in human polymorphonuclear neutrophils and HL60 cells stimulated with TNF-alpha. Biochem. Biophys. Res. Commun. 2005, 337, 464–473. [Google Scholar] [CrossRef]
- Shridas, P.; Webb, N.R. Diverse Functions of Secretory Phospholipases A2. Adv. Vasc. Med. 2014, 2014, 689815. [Google Scholar] [CrossRef] [Green Version]
- Hallstrand, T.S.; Chi, E.Y.; Singer, A.G.; Gelb, M.H.; Henderson, W.R. Secreted phospholipase A2 group X overexpression in asthma and bronchial hyperresponsiveness. Am. J. Respir. Crit. Care Med. 2007, 176, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Hallstrand, T.S.; Lai, Y.; Altemeier, W.A.; Appel, C.A.; Johnson, B.; Frevert, C.W.; Hudkins, K.L.; Bollinger, J.G.; Woodruff, P.G.; Hyde, D.M.; et al. Regulation and function of epithelial secreted phospholipase A2 group X in asthma. Am. J. Respir. Crit. Care Med. 2013, 188, 42–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolin, J.D.; Lai, Y.; Ogden, H.L.; Manicone, A.M.; Murphy, R.C.; An, D.; Frevert, C.W.; Ghomashchi, F.; Naika, G.S.; Gelb, M.H.; et al. Secreted PLA2 group X orchestrates innate and adaptive immune responses to inhaled allergen. JCI Insight 2017, 2, 94929. [Google Scholar] [CrossRef] [Green Version]
- Hart, L.A.; Krishnan, V.L.; Adcock, I.M.; Barnes, P.J.; Chung, K.F. Activation and localization of transcription factor, nuclear factor-kappaB, in asthma. Am. J. Respir. Crit. Care Med. 1998, 158, 1585–1592. [Google Scholar] [CrossRef]
- Poynter, M.E.; Irvin, C.G.; Janssen-Heininger, Y.M. Rapid activation of Nuclear Factor- B in airway epithelium in a murine model of allergic airway inflammation. Am. J. Pathol. 2002, 160, 1325–1334. [Google Scholar] [CrossRef]
- Khaddaj-Mallat, R.; Sirois, C.; Sirois, M.; Rizcallah, E.; Marouan, S.; Morin, C.; Rousseau, E. Pro-Resolving Effects of Resolvin D2 in LTD4 and TNF-α Pre-Treated Human Bronchi. PLoS ONE 2016, 11, e0167058. [Google Scholar] [CrossRef]
- Khaddaj-Mallat, R.; Rousseau, E. MAG-EPA and 17,18-EpETE target cytoplasmic signalling pathways to reduce short-term airway hyperresponsiveness. Pflugers Arch. Eur. J. Physiol. 2015, 467, 1591–1605. [Google Scholar] [CrossRef]
- Kantrow, S.P.; Shen, Z.; Jagneaux, T.; Zhang, P.; Nelson, S. Neutrophil-mediated lung permeability and host defense proteins. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, 738–745. [Google Scholar] [CrossRef] [Green Version]
- Maus, U.; von Grote, K.; Kuziel, W.A.; Mack, M.; Miller, E.J.; Cihak, J.; Stangassinger, M.; Maus, R.; Schlöndorff, D.; Seeger, W.; et al. The role of CC chemokine receptor 2 in alveolar monocyte and neutrophil immigration in intact mice. Am. J. Respir. Crit. Care Med. 2002, 166, 268–273. [Google Scholar] [CrossRef]
- Kraneveld, A.D.; Van der Kleij, H.P.; Kool, M.; Van Houwelingen, A.H.; Weitenberg, A.C.; Redegeld, F.A.; Nijkamp, F.P. Key role for mast cells in nonatopic asthma. J. Immunol. 2002, 169, 2044–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biedermann, T.; Kneilling, M.; Mailhammer, R.; Maier, K.; Sander, C.A.; Kollias, G.; Kunkel, S.L.; Hultner, L.; Rocken, M. Mast cells control neutrophil recruitment during T cell-mediated delayed-type hypersensitivity reactions through tumor necrosis factor and macrophage inflammatory protein 2. J. Exp. Med. 2000, 192, 1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verheijden, K.A.; Henricks, P.A.; Redegeld, F.A.; Garssen, J.; Folkerts, G. Measurement of airway function using invasive and non-invasive methods in mild and severe models for allergic airway inflammation in mice. Front. Pharmacol. 2014, 5, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakae, S.; Ho, L.H.; Yu, M.; Monteforte, R.; Iikura, M.; Suto, H.; Galli, S.J. Mast cell-derived TNF contributes to airway hyperreactivity, inflammation, and TH2 cytokine production in an asthma model in mice. J. Allergy Clin. Immunol. 2007, 120, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Tarkowski, M.; Vanoirbeek, J.A.; Vanhooren, H.M.; De Vooght, V.; Mercier, C.M.; Ceuppens, J.; Nemery, B.; Hoet, P.H. Immunological determinants of ventilatory changes induced in mice by dermal sensitization and respiratory challenge with toluene diisocyanate. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Kanagaratham, C.; Marino, R.; Camateros, P.; Ren, J.; Houle, D.; Sladek, R.; Vidal, S.M.; Radzioch, D. Mapping of a chromosome 12 region associated with airway hyperresponsiveness in a recombinant congenic mouse strain and selection of potential candidate genes by expression and sequence variation analyses. PLoS ONE 2014, 9, e104234. [Google Scholar] [CrossRef]
- Finkelmann, F.D. Use of unrestrained, single-chamber barometric plethysmography to evaluate sensitivity to cholinergic stimulation in mouse models of allergic airway disease. J. Allergy Clin. Immunol. 2008, 121, 334–335. [Google Scholar] [CrossRef]
- De Vooght, V.; Vanoirbeek, J.A.; Luyts, K.; Haenen, S.; Nemery, B.; Hoet, P.H. Choice of mouse strain influences the outcome in a mouse model of chemical-induced asthma. PLoS ONE 2010, 5, e12581. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.K.; Foster, P.S. Are mouse models of asthma appropriate for investigating the pathogenesis of airway hyper-responsiveness? Front. Physiol. 2012, 31, 312. [Google Scholar] [CrossRef] [Green Version]
- Tränkner, D.; Hahne, N.; Suginoa, K.; Hoonb, M.A.; Zukera, C. Population of sensory neurons essential for asthmatic hyperreactivity of inflamed airways. Proc. Natl. Acad. Sci. USA 2014, 111, 11515–11520. [Google Scholar] [CrossRef] [Green Version]
- Reich, A.; Szepietewski, J.C. Non-analgesic effects of opioids: Peripheral opioid receptors as promising targets for future anti-pruritic therapies. Curr. Pharm. Des. 2012, 18, 6021–6024. [Google Scholar] [CrossRef] [PubMed]
- Kaczyńska, K.; Zając, D.; Wojciechowski, P.; Kogut, E.; Szereda-Przestaszewska, M. Neuropeptides and breathing in health and disease. Pulm. Pharmacol. Ther. 2018, 48, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Carraway, R.E.; Cochrane, D.E.; Salmonsen, R.; Muraki, K.; Boucher, W. Neurotensin elevates hematocrit and plasma levels of the leukotrienes, LTB4, LTC4, LTD4 and LTE4, in anesthetized rats. Peptides 1991, 12, 1105–1111. [Google Scholar] [CrossRef]
- Robbins, R.A.; Nelson, K.J.; Gossman, G.L.; Rubinstein, I. Neurotensin stimulates neutrophil adherence to bronchial epithelial cells in vitro. Life Sci. 1995, 56, 1353–1359. [Google Scholar] [CrossRef]
- Koon, H.W.; Kim, Y.S.; Xu, H.; Kumar, A.; Zhao, D.; Karagiannides, I.; Dobner, P.R.; Pothoulakis, C. Neurotensin induces IL-6 secretion in mouse preadipocytes and adipose tissues during 2,4,6,-trinitrobenzensulphonic acid-induced colitis. Proc. Natl. Acad. Sci. USA 2009, 106, 8766–8771. [Google Scholar] [CrossRef] [Green Version]
- Merrifield, R.B. Solid Phase Peptide Synthesis. The synthesis of a tetrapeptide. J. Am. Chem. 1963, 8, 2149–2154. [Google Scholar] [CrossRef]
- Kaczyńska, K.; Wojciechowski, P.; Jampolska, M.; Lipkowski, A.W.; Kleczkowska, P. Cardiovascular and respiratory activity of PK20, opioid and neurotensin hybrid peptide in anesthetized and awake rats. Eur. J. Pharmacol. 2017, 797, 20–25. [Google Scholar] [CrossRef]
- Tournoy, K.G.; Kips, J.C.; Schou, C.; Pauwels, R.A. Airway eosinophilia is not a requirement for allergen-induced airway hyperresponsiveness. Clin. Exp. Allergy 2000, 30, 79–85. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russjan, E.; Andrzejewski, K.; Sulejczak, D.; Kleczkowska, P.; Kaczyńska, K. Endomorphin-2- and Neurotensin- Based Chimeric Peptide Attenuates Airway Inflammation in Mouse Model of Nonallergic Asthma. Int. J. Mol. Sci. 2019, 20, 5935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235935
Russjan E, Andrzejewski K, Sulejczak D, Kleczkowska P, Kaczyńska K. Endomorphin-2- and Neurotensin- Based Chimeric Peptide Attenuates Airway Inflammation in Mouse Model of Nonallergic Asthma. International Journal of Molecular Sciences. 2019; 20(23):5935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235935
Chicago/Turabian StyleRussjan, Ewelina, Kryspin Andrzejewski, Dorota Sulejczak, Patrycja Kleczkowska, and Katarzyna Kaczyńska. 2019. "Endomorphin-2- and Neurotensin- Based Chimeric Peptide Attenuates Airway Inflammation in Mouse Model of Nonallergic Asthma" International Journal of Molecular Sciences 20, no. 23: 5935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20235935