Astragaloside IV Alleviates Ammonia-Induced Apoptosis and Oxidative Stress in Bovine Mammary Epithelial Cells


Abstract
:1. Introduction
2. Results
2.1. Effect of Astragaloside IV on Ammonia-Induced Bovine Mammary Epithelial Cell Death
2.2. Effects of Astragaloside IV on mRNA Expressions of Apoptosis-Related Genes Induced by Ammonia in Bovine Mammary Epithelial Cells
2.3. Effects of Ammonia and Astragaloside IV Treatment on p53 Signaling Pathways in Bovine Mammary Epithelial Cells
2.4. Effects of Astragaloside IV on mRNA Expression of Inflammatory Factors (IL-6 and IL-8) Induced by Ammonia in Bovine Mammary Epithelial Cells
2.5. Effects of Astragaloside IV on mRNA Expression of Endoplasmic Reticulum Stress Markers (CHOP and GPR78) Induced by Ammonia in Bovine Mammary Epithelial Cells
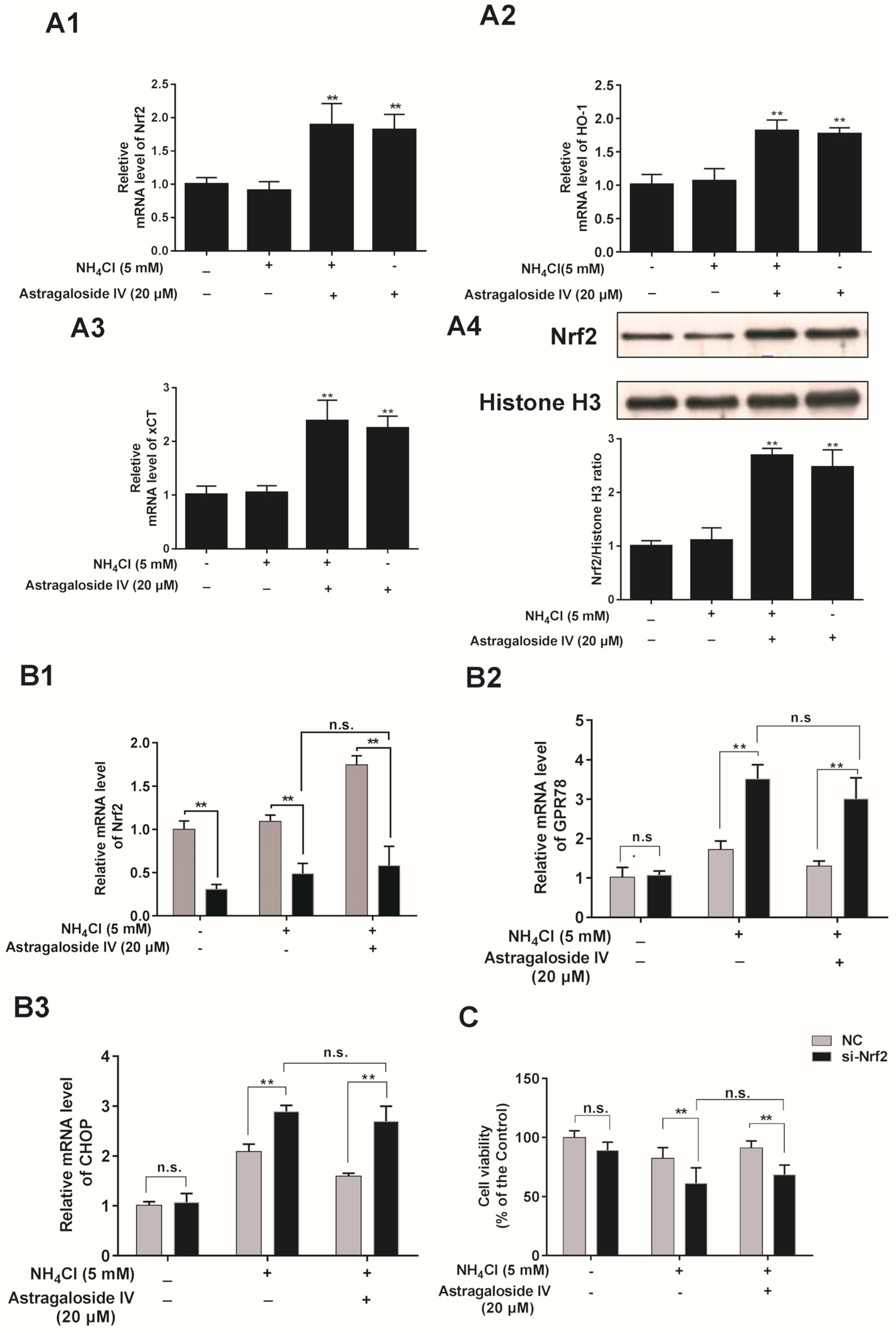
2.6. The Cytoprotective Effects of Astragaloside IV against Oxidative Stress are Dependent on the Induction of Nrf2 in Bovine Mammary Epithelial Cells
2.7. Effects of Ammonia and Astragaloside IV Treatment on the AKT and ERK Signaling Pathways in Bovine Mammary Epithelial Cells
2.8. Activation of AKT and ERK Is Required for the Cytoprotective Effects of Astragaloside IV in Bovine Mammary Epithelial Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Cell Viability Assay
4.3. Detection of Intracellular ROS
4.4. Quantitative Measurement of Apoptosis by AnnexinV-FITC/PI Staining
4.5. RNA Isolation and Quantitative Real-Time PCR (qPCR)
4.6. Transfection and Nrf2 Small RNA Interference
4.7. Western Blot Analysis
4.8. Immunofluorescent Staining
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ROS | Reactive oxygen species |
| CHOP | C/EBP homologous protein |
| GRP78 | Glucose-regulated protein 78 |
| HO-1 | Hemeoxygenase 1 |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
References
- Visek, W.J. Ammonia: Its effects on biological systems, metabolic hormones, and reproduction. J. Dairy Sci. 1984, 67, 481–498. [Google Scholar] [CrossRef]
- Treacher, R.J.; Little, W.; Collis, K.A.; Stark, A.J. The influence of dietary protein intake on milk production and blood composition of high-yielding dairy cows. J. Dairy Res. 1976, 43, 357–369. [Google Scholar] [CrossRef]
- Wang, F.; Chen, S.; Jiang, Y.; Zhao, Y.; Sun, L.; Zheng, B.; Chen, L.; Liu, Z.; Zheng, X.; Yi, K.; et al. Effects of ammonia on apoptosis and oxidative stress in bovine mammary epithelial cells. Mutagenesis 2018, 33, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.K.; Brzezinska-Slebodzinska, E.; Madsen, F.C. Oxidative stress, antioxidants, and animal function. J. Dairy Sci. 1993, 76, 2812–2823. [Google Scholar] [CrossRef]
- Ko, J.K.S.; Lam, F.Y.L.; Cheung, A.P.L. Amelioration of experimental colitis by Astragalus membranaceus through anti-oxidation and inhibition of adhesion molecule synthesis. World J. Gastroenterol. 2005, 11, 5787–5794. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.J.; Hufnagl, P.; Binder, B.R.; Wojta, J. Antiinflammatory activity of astragaloside IV is mediated by inhibition of NF-kappaB activation and adhesion molecule expression. Thromb. Haemost. 2003, 90, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Pugliese, M.; Pugliese, A.; Passantino, A. Biological active ingredients of traditional Chinese herb Astragalus membranaceus on treatment of diabetes: A systematic review. Mini Rev. Med. Chem. 2015, 15, 315–329. [Google Scholar]
- Zhang, W.D.; Zhang, C.; Wang, X.H.; Gao, P.J.; Zhu, D.L.; Chen, H.; Liu, R.H.; Li, H.L. Astragaloside IV dilates aortic vessels from normal and spontaneously hypertensive rats through endothelium-dependent and endothelium-independent ways. Planta Med. 2006, 72, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.P.; Cao, Q. Effects of astragaloside IV on myocardial calcium transport and cardiac function in ischemic rats. Acta Pharmacol. Sin. 2002, 23, 898–904. [Google Scholar] [PubMed]
- Luo, Y.; Qin, Z.; Hong, Z.; Zhang, X.; Ding, D.; Fu, J.H.; Zhang, W.D.; Chen, J. Astragaloside IV protects against ischemic brain injury in a murine model of transient focal ischemia. Neurosci. Lett. 2004, 363, 218–223. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Han, C.; Wang, X.; Xing, G.; Zhou, L.; Li, G.; Niu, Y. Astragaloside IV suppresses collagen production of activated hepatic stellate cells via oxidative stress-mediated p38 MAPK pathway. Free Radic. Biol. Med. 2013, 60, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhang, Y.; Sun, S.; Shen, J.; Qiu, J.; Yin, X.; Yin, H.; Jiang, S. Inhibitory effects of astragaloside IV on diabetic peripheral neuropathy in rats. Can. J. Physiol. Pharmacol. 2006, 84, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Jayakumar, A.R.; Panickar, K.S.; Murthy Ch, R.; Norenberg, M.D. Oxidative stress and mitogen-activated protein kinase phosphorylation mediate ammonia-induced cell swelling and glutamate uptake inhibition in cultured astrocytes. J. Neurosci. Off. J. Soc. Neurosc. 2006, 26, 4774–4784. [Google Scholar] [CrossRef] [PubMed]
- Kosenko, E.; Kaminski, Y.; Lopata, O.; Muravyov, N.; Felipo, V. Blocking NMDA receptors prevents the oxidative stress induced by acute ammonia intoxication. Free Radic. Biol. Med. 1999, 26, 1369–1374. [Google Scholar] [CrossRef]
- Celi, P.; Di Trana, A.; Quaranta, A. Metabolic profile and oxidative status in goats during the peripartum period. Aust. J. Exp. Agric. 2008, 48, 1004–1008. [Google Scholar] [CrossRef]
- Jozwik, A.; Krzyzewski, J.; Strzalkowska, N.; Polawska, E.; Bagnicka, E.; Wierzbicka, A.; Niemczuk, K.; Lipinska, P.; Horbanczuk, J.O. Relations between the oxidative status, mastitis, milk quality and disorders of reproductive functions in dairy cows—A review. Anim. Sci. Pap. Rep. 2012, 30, 297–307. [Google Scholar]
- Miranda, S.G.; Purdie, N.G.; Osborne, V.R.; Coomber, B.L.; Cant, J.P. Selenomethionine increases proliferation and reduces apoptosis in bovine mammary epithelial cells under oxidative stress. J. Dairy Sci. 2011, 94, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.K. Oxidative stress in neurodegeneration: Cause or consequence? Nat. Med. 2004, 10, S18–S25. [Google Scholar] [CrossRef]
- Chen, X.Y.; Shao, J.Z.; Xiang, L.X.; Liu, X.M. Involvement of apoptosis in malathion-induced cytotoxicity in a grass carp (Ctenopharyngodon idellus) cell line. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2006, 142, 36–45. [Google Scholar] [CrossRef]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef]
- Wu, C.C.; Howell, K.E.; Neville, M.C.; Yates, J.R.; McManaman, J.L. Proteomics reveal a link between the endoplasmic reticulum and lipid secretory mechanisms in mammary epithelial cells. Electrophoresis 2000, 21, 3470–3482. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a double-edged sword? Antioxid. Redox Signal. 2007, 9, 2277–2293. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, S.J.; Yun, C.Y.; Oyadomari, S.; Novoa, I.; Zhang, Y.; Jungreis, R.; Nagata, K.; Harding, H.P.; Ron, D. CHOP induces death by promoting protein synthesis and oxidation in the stressed endoplasmic reticulum. Genes Dev. 2004, 18, 3066–3077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Kaufman, R.J. From endoplasmic-reticulum stress to the inflammatory response. Nature 2008, 454, 455–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, T.; Itoh, K.; Yamamoto, M. Roles of Nrf2 in activation of antioxidant enzyme genes via antioxidant responsive elements. Methods Enzymol. 2002, 348, 182–190. [Google Scholar] [PubMed]
- Itoh, K.; Mimura, J.; Yamamoto, M. Discovery of the negative regulator of Nrf2, Keap1: A historical overview. Antioxid. Redox Signal. 2010, 13, 1665–1678. [Google Scholar] [CrossRef]
- Gu, D.M.; Lu, P.H.; Zhang, K.; Wang, X.; Sun, M.; Chen, G.Q.; Wang, Q. EGFR mediates astragaloside IV-induced Nrf2 activation to protect cortical neurons against in vitro ischemia/reperfusion damages. Biochem. Biophys. Res. Commun. 2015, 457, 391–397. [Google Scholar] [CrossRef]
- Kang, K.W.; Lee, S.J.; Kim, S.G. Molecular mechanism of nrf2 activation by oxidative stress. Antioxid. Redox Signal. 2005, 7, 1664–1673. [Google Scholar] [CrossRef]
- Huang, X.P.; Qiu, Y.Y.; Wang, B.; Ding, H.; Tang, Y.H.; Zeng, R.; Deng, C.Q. Effects of Astragaloside IV combined with the active components of Panax notoginseng on oxidative stress injury and nuclear factor-erythroid 2-related factor 2/heme oxygenase-1 signaling pathway after cerebral ischemia-reperfusion in mice. Pharmacogn. Mag. 2014, 10, 402–409. [Google Scholar]
- Sun, J.; Chen, X.L.; Zheng, J.Y.; Zhou, J.W.; Ma, Z.L. Astragaloside IV protects new born rats from anesthesia-induced apoptosis in the developing brain. Exp. Ther. Med. 2016, 12, 1829–1835. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′-3′) | Product Length (bp) |
|---|---|---|
| BAX | Sense Primer: TGCTTCAGGGTTTCATCC Anti-sense Primer: CTTCAGACACTCGCTCAG | 116 |
| BCL-2 | Sense Primer: TTCTCCTGGCTGTCTCTG Anti-sense Primer: CTGCTTCTTGAATCTTCTGC | 135 |
| caspase 3 | Sense Primer: GTTCATCCAGGCTCTTTGTG Anti-sense Primer: AAGGACTCATATTCTATTGCTACC | 108 |
| p53 | Sense Primer: GAAGACCTACCCTGGCAATTAC Anti-sense Primer: AGAACAGCTTGTTAAGGGAAGG | 104 |
| GAPDH | Sense Primer: GTTCAACGGCACAGTCAAG Anti-sense Primer: TACTCAGCACCAGCATCAC | 117 |
| Nrf2 | Sense Primer: CCAGCACAACACATACCATCAG Anti-sense Primer: CGTAGCCGAAGAAACCTCATTG | 156 |
| CHOP | Sense Primer: TGAACGACTCAAACAGGAAATC Anti-sense Primer: ACGCTAAGACCCTTTTCTATCG | 248 |
| GPR78 | Sense Primer: GACCCTGACTCGGGCTAAAT Anti-sense Primer: TGGACAGCGGCACCATATG | 243 |
| HO-1 | Sense Primer: GGCAGCAAGGTGCAAGA Anti-sense Primer: GAAGGAAGCCAGCCAAGAG | 221 |
| xCT | Sense Primer: GATACAAACGCCCAGATATGC Anti-sense Primer: ATGATGAAGCCAATCCCTGTA | 136 |
| IL6 | Sense Primer: ATGCTTCCAATCTGGGTTC Anti-sense Primer: TGAGGATAATCTTTGCGTTC | 269 |
| IL8 | Sense Primer: GCTGGCTGTTGCTCTCTTG Anti-sense Primer: GGGTGGAAAGGTGTGGAATG | 126 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, F.; Zhao, Y.; Chen, S.; Chen, L.; Sun, L.; Cao, M.; Li, C.; Zhou, X. Astragaloside IV Alleviates Ammonia-Induced Apoptosis and Oxidative Stress in Bovine Mammary Epithelial Cells. Int. J. Mol. Sci. 2019, 20, 600. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030600
Wang F, Zhao Y, Chen S, Chen L, Sun L, Cao M, Li C, Zhou X. Astragaloside IV Alleviates Ammonia-Induced Apoptosis and Oxidative Stress in Bovine Mammary Epithelial Cells. International Journal of Molecular Sciences. 2019; 20(3):600. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030600
Chicago/Turabian StyleWang, Fengge, Yun Zhao, Shuxiong Chen, Lu Chen, Liting Sun, Maosheng Cao, Chunjin Li, and Xu Zhou. 2019. "Astragaloside IV Alleviates Ammonia-Induced Apoptosis and Oxidative Stress in Bovine Mammary Epithelial Cells" International Journal of Molecular Sciences 20, no. 3: 600. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030600