Sulforaphane Protect Against Cadmium-Induced Oxidative Damage in mouse Leydigs Cells by Activating Nrf2/ARE Signaling Pathway

 ,
,
Abstract
:1. Introduction
2. Results
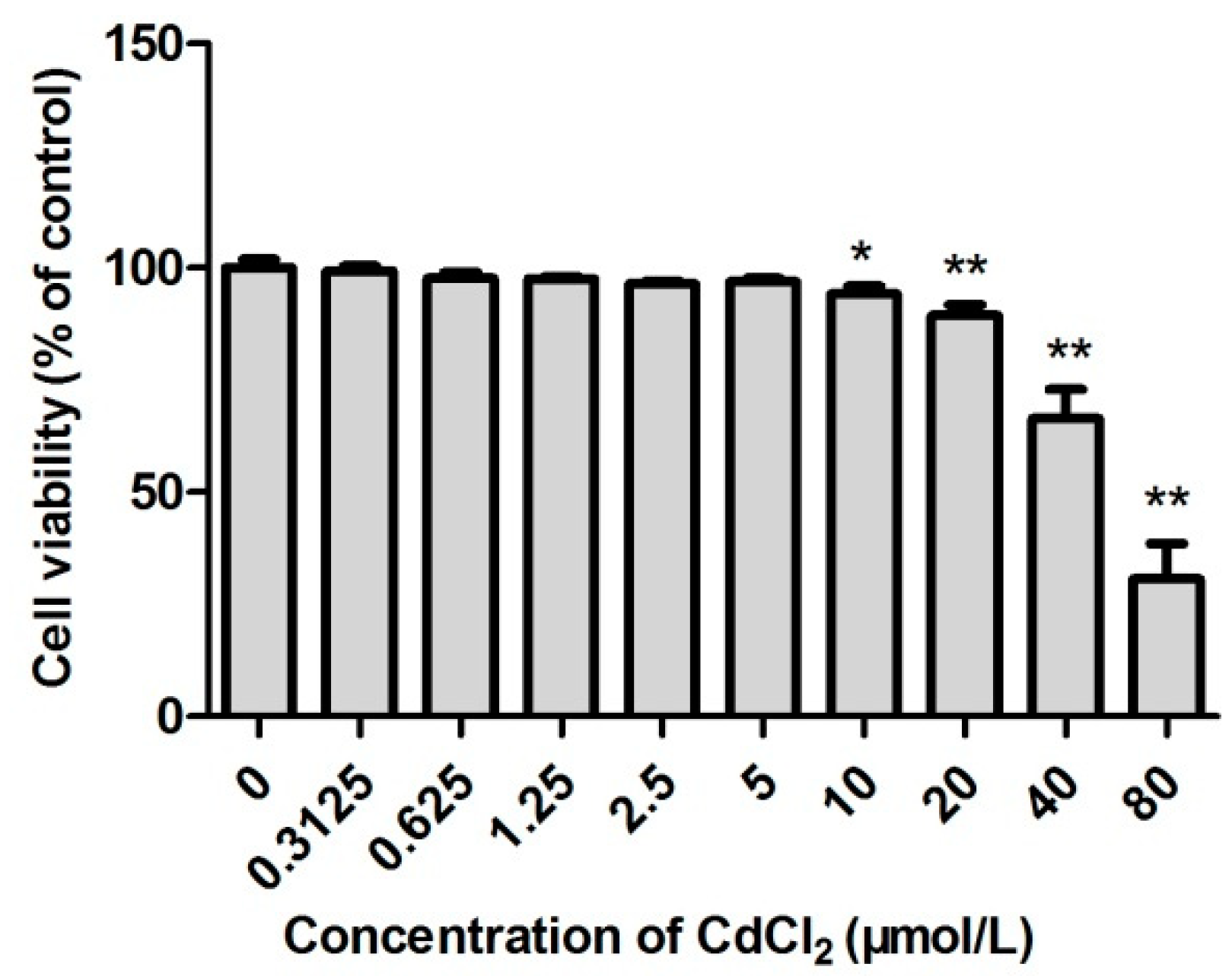
2.1. The Survival Rate of Cd-Induced TM3 Cells
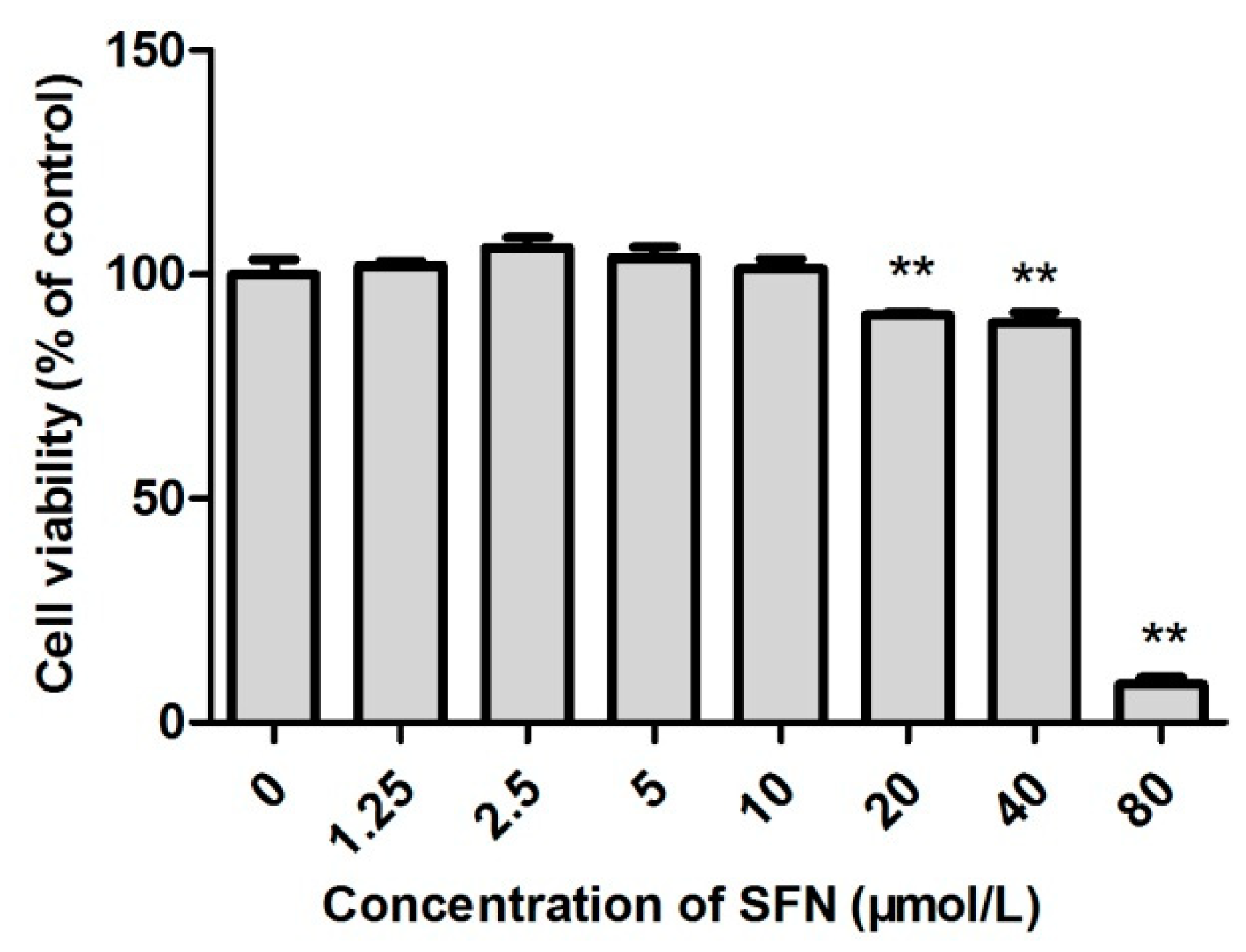
2.2. Survival Rate of SFN-Treated TM3 Cells
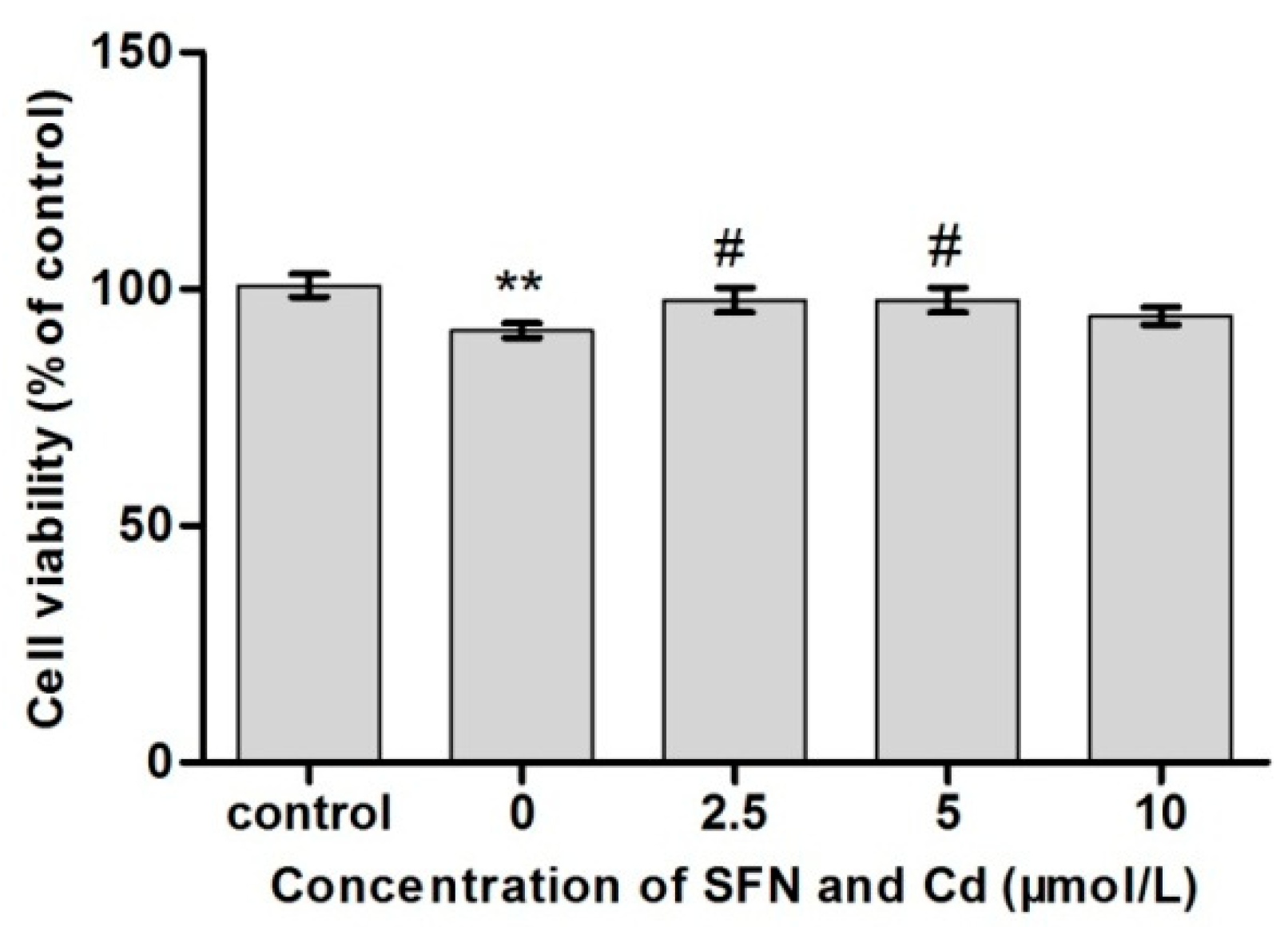
2.3. Survival Rate of TM3 Cells Treated with Cd + SFN
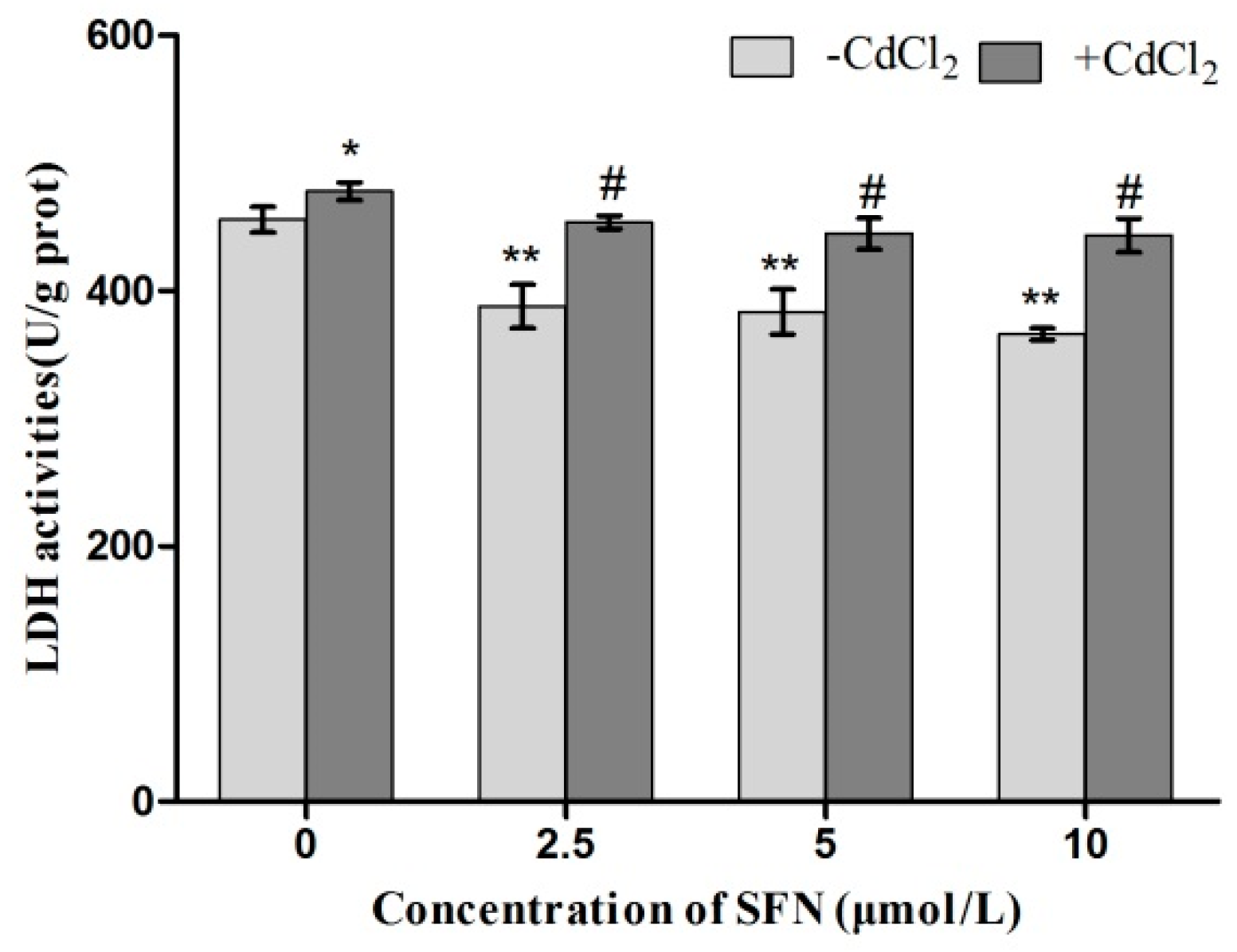
2.4. LDH Activity in TM3 Cells Treated with Cd + SFN
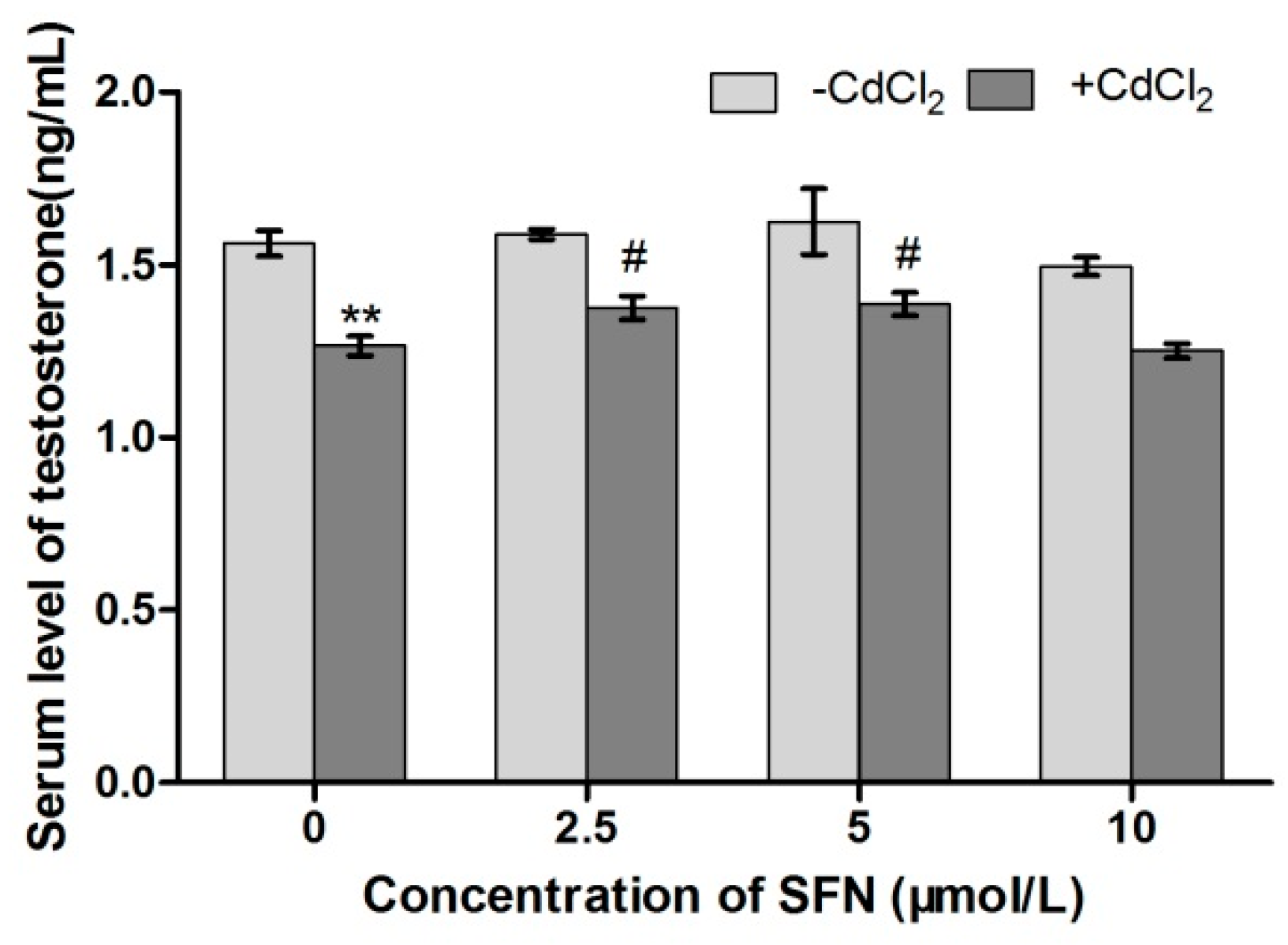
2.5. Concentration of Testosterone
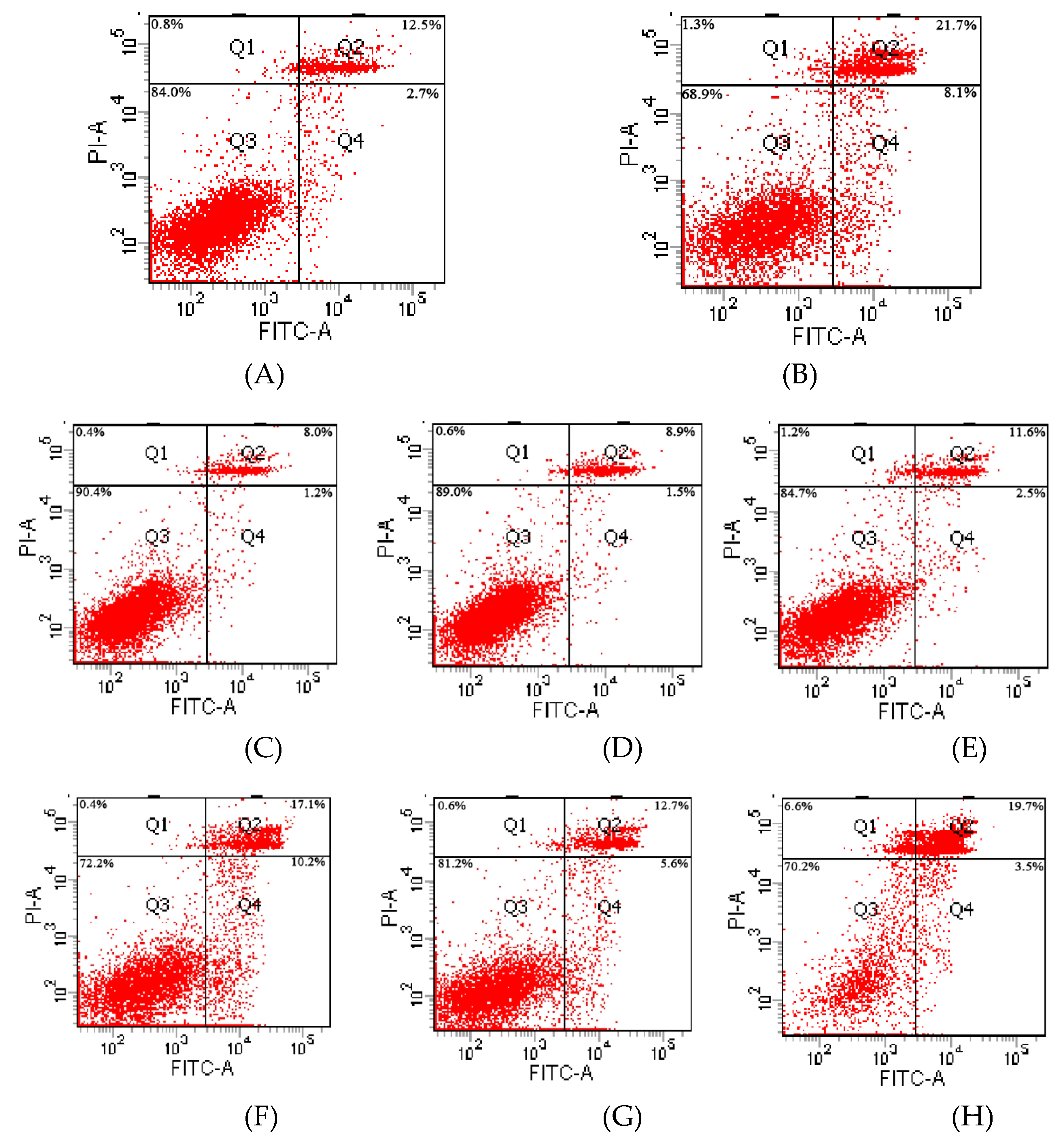
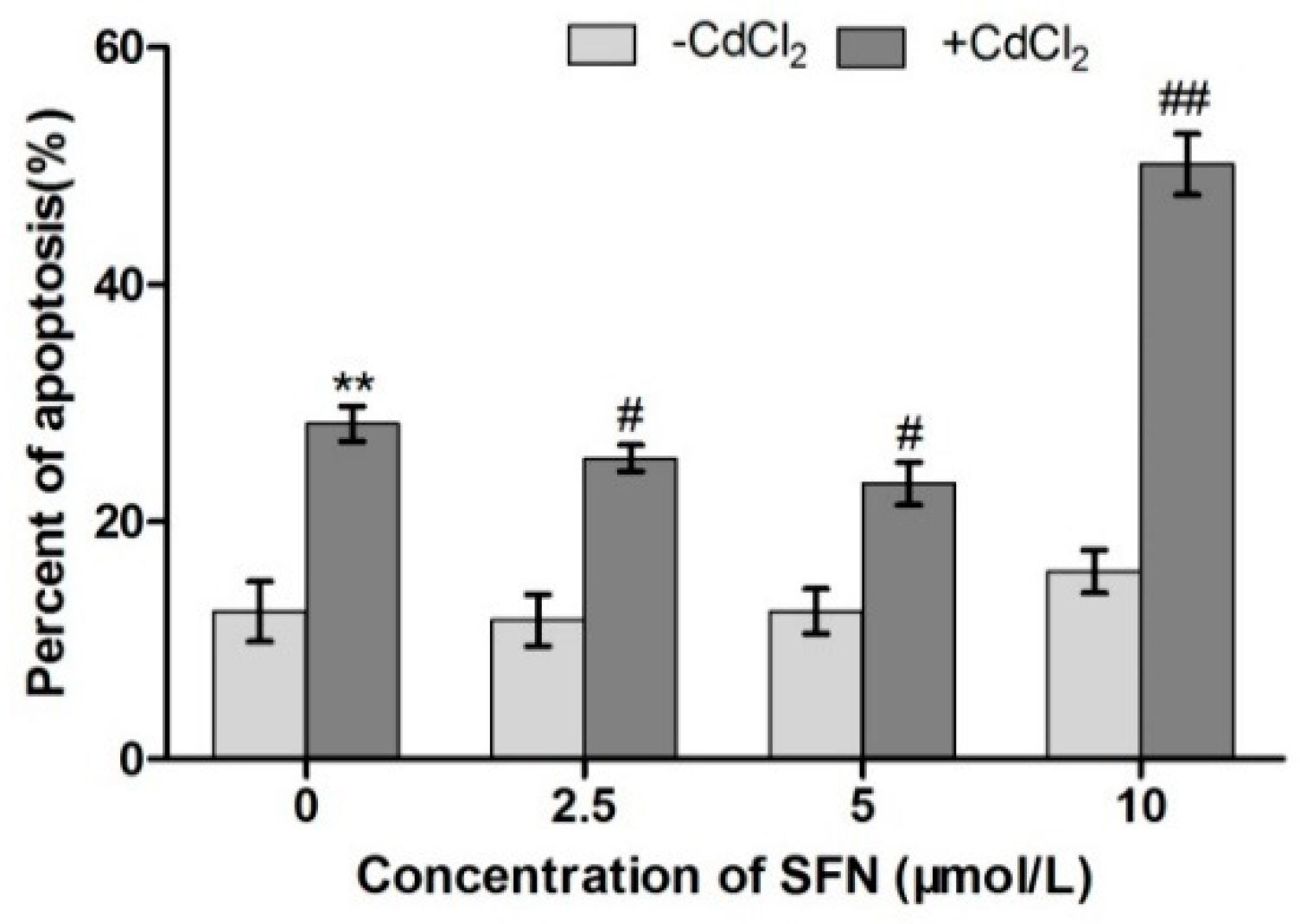
2.6. TM3 Cell Apoptosis
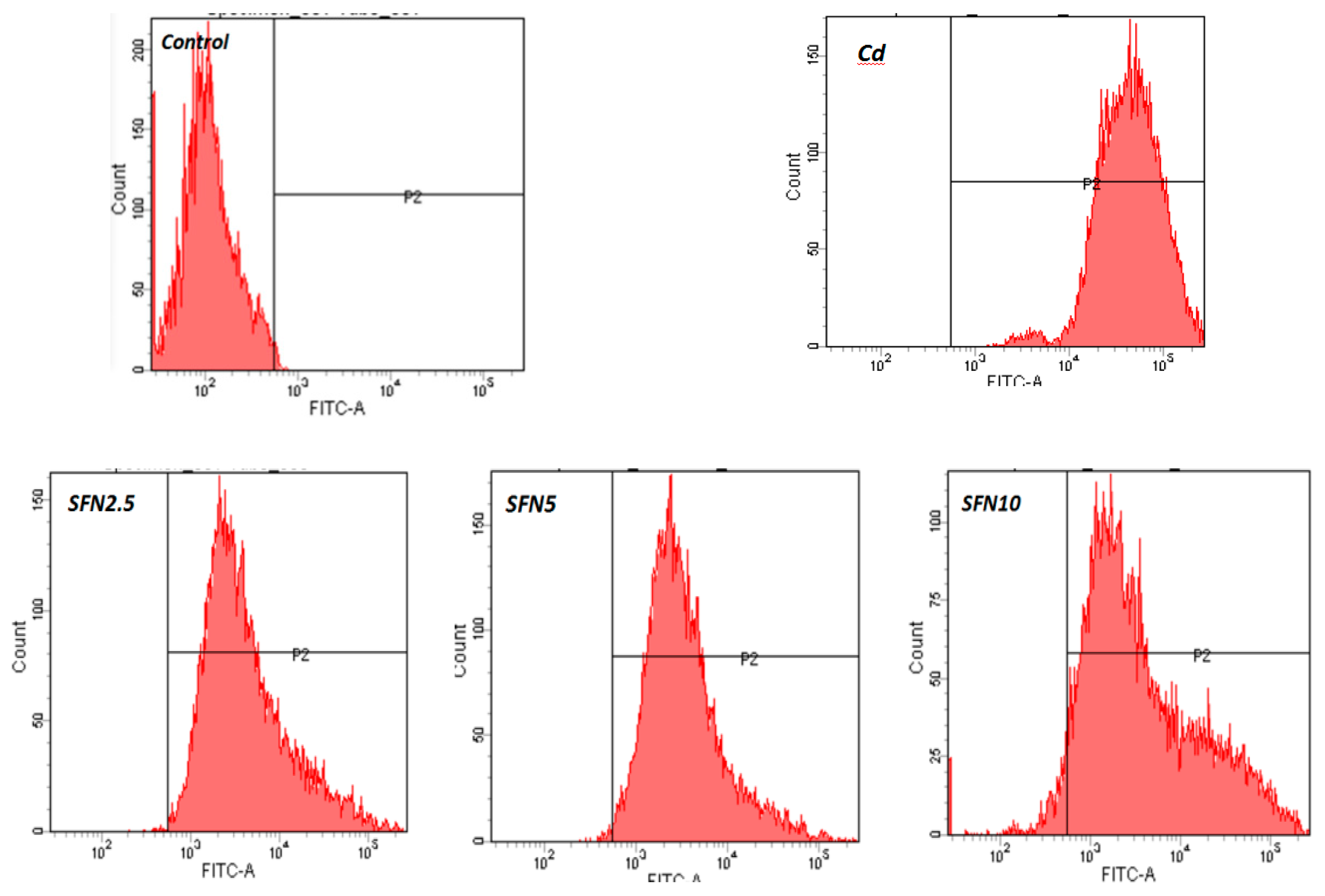
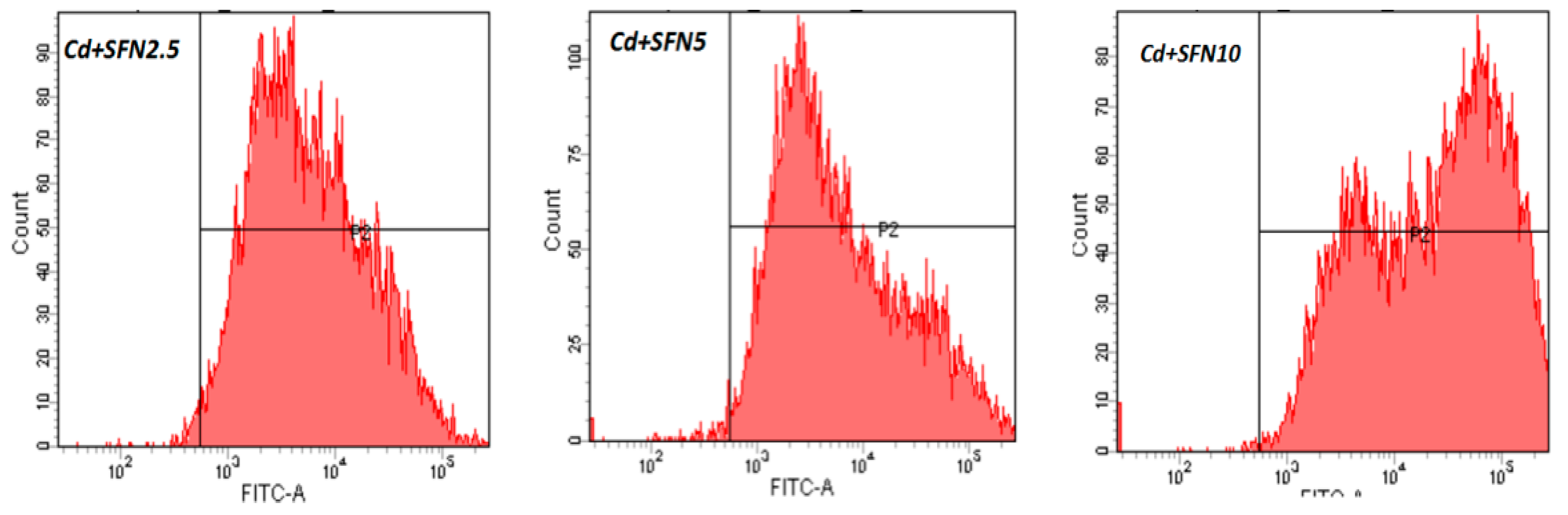
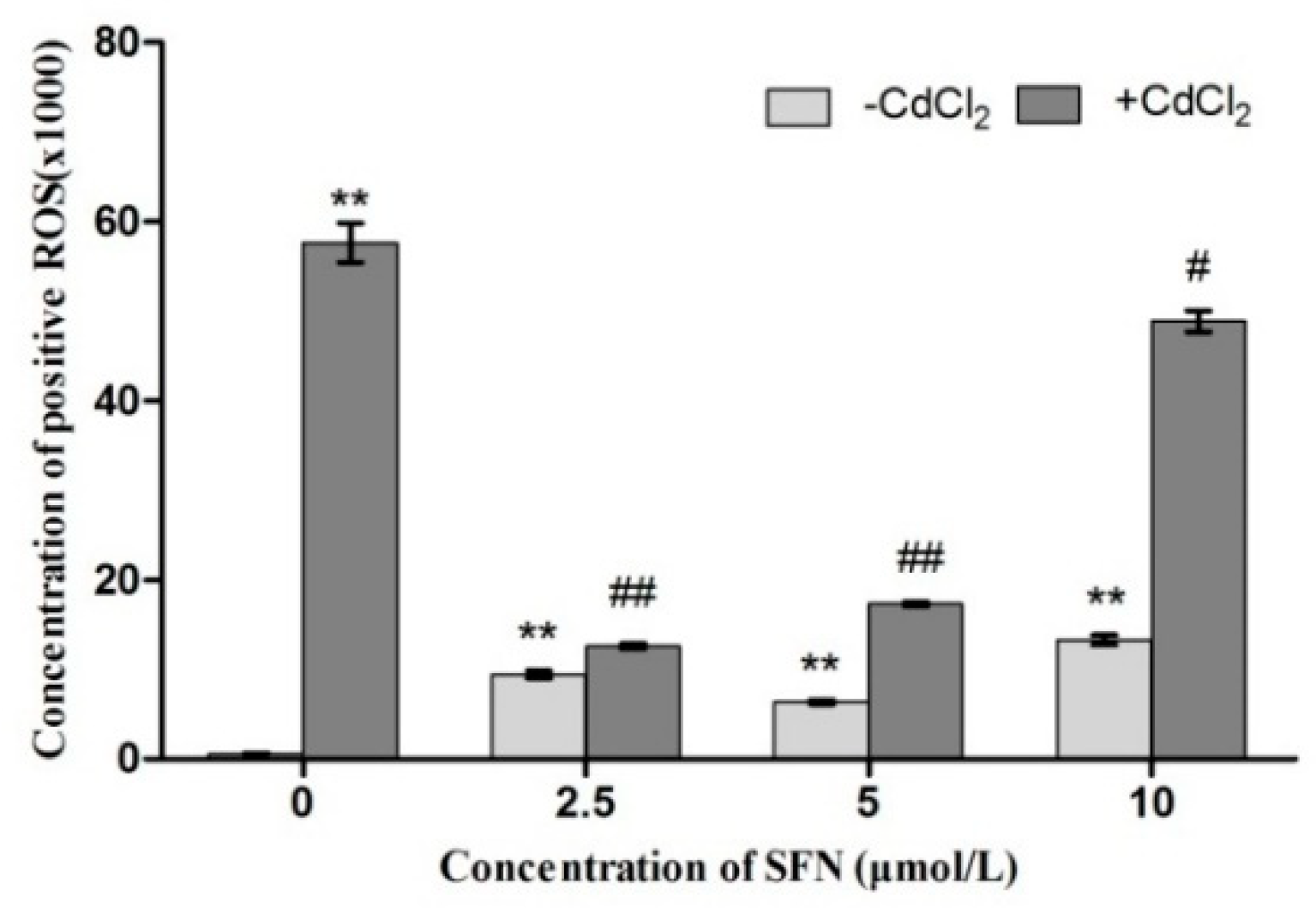
2.7. Detection of ROS Released by TM3 Cells
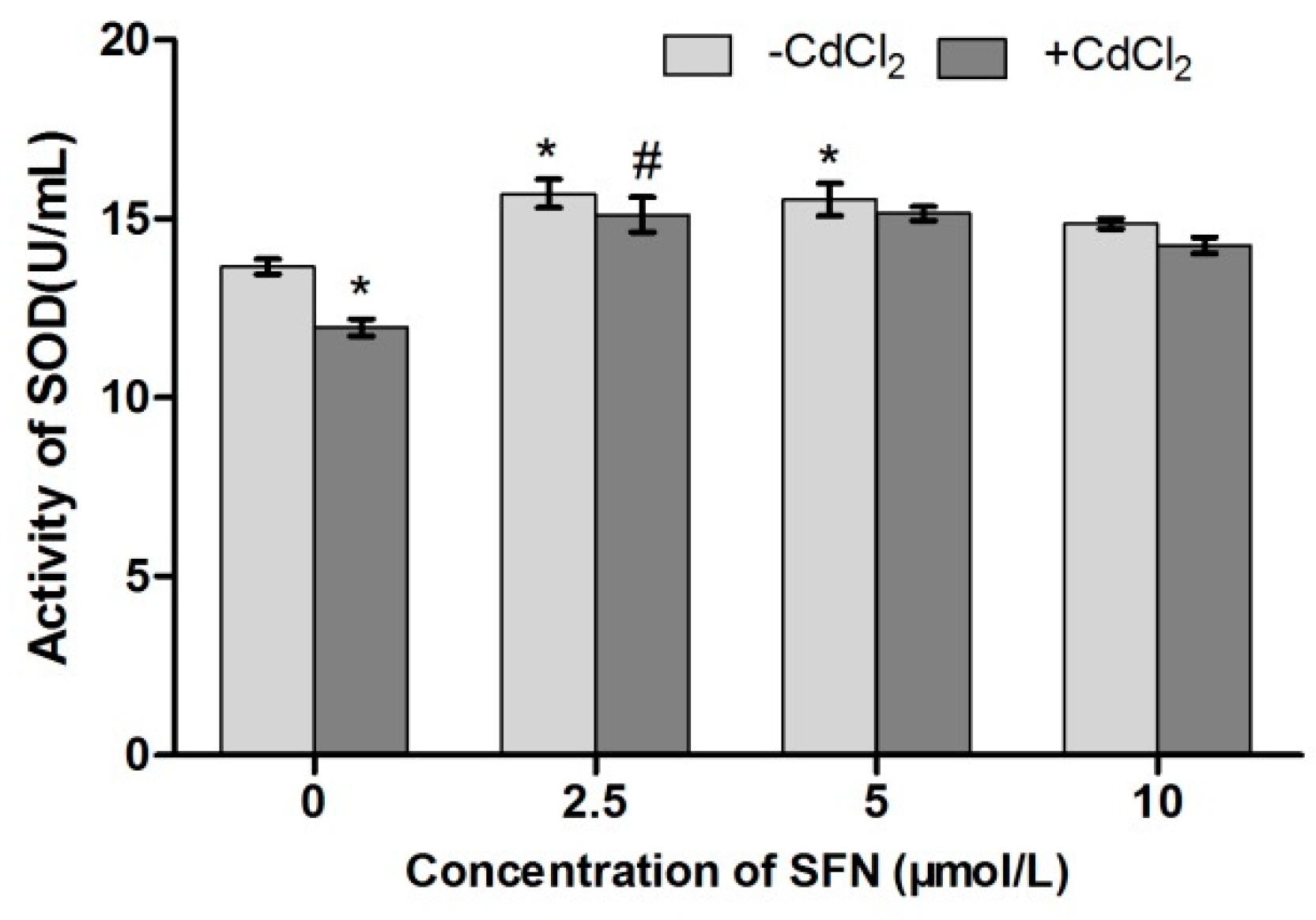
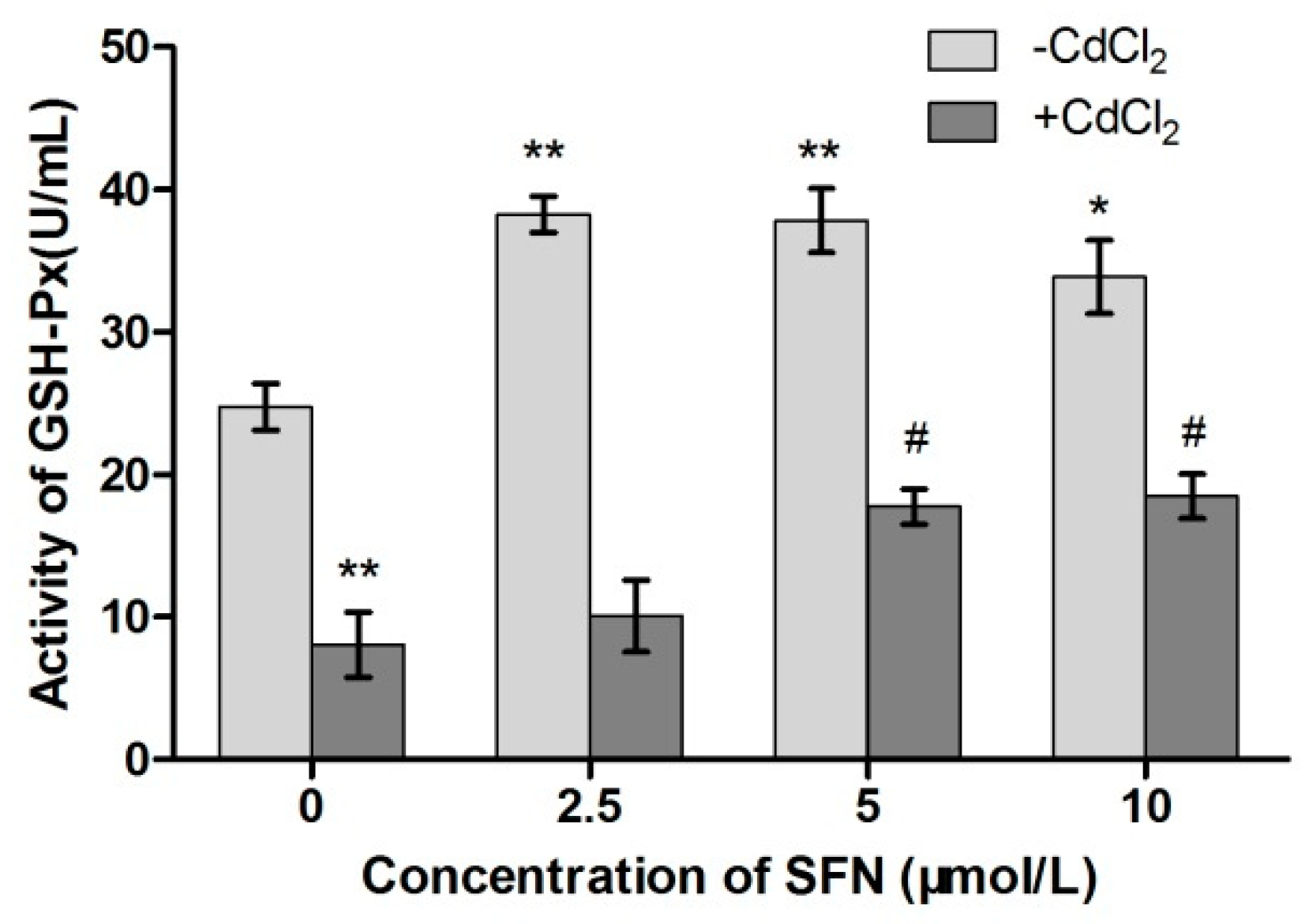
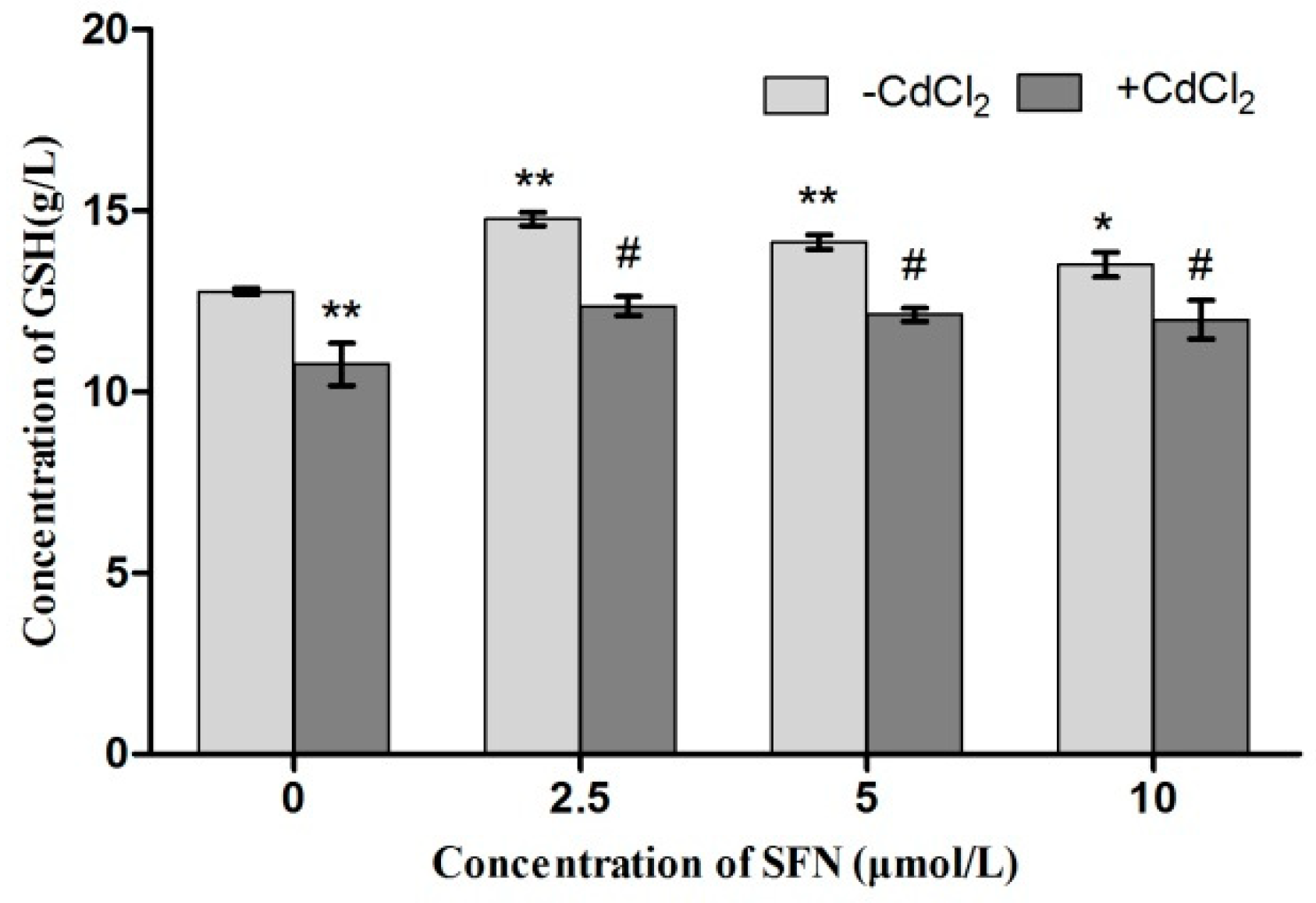
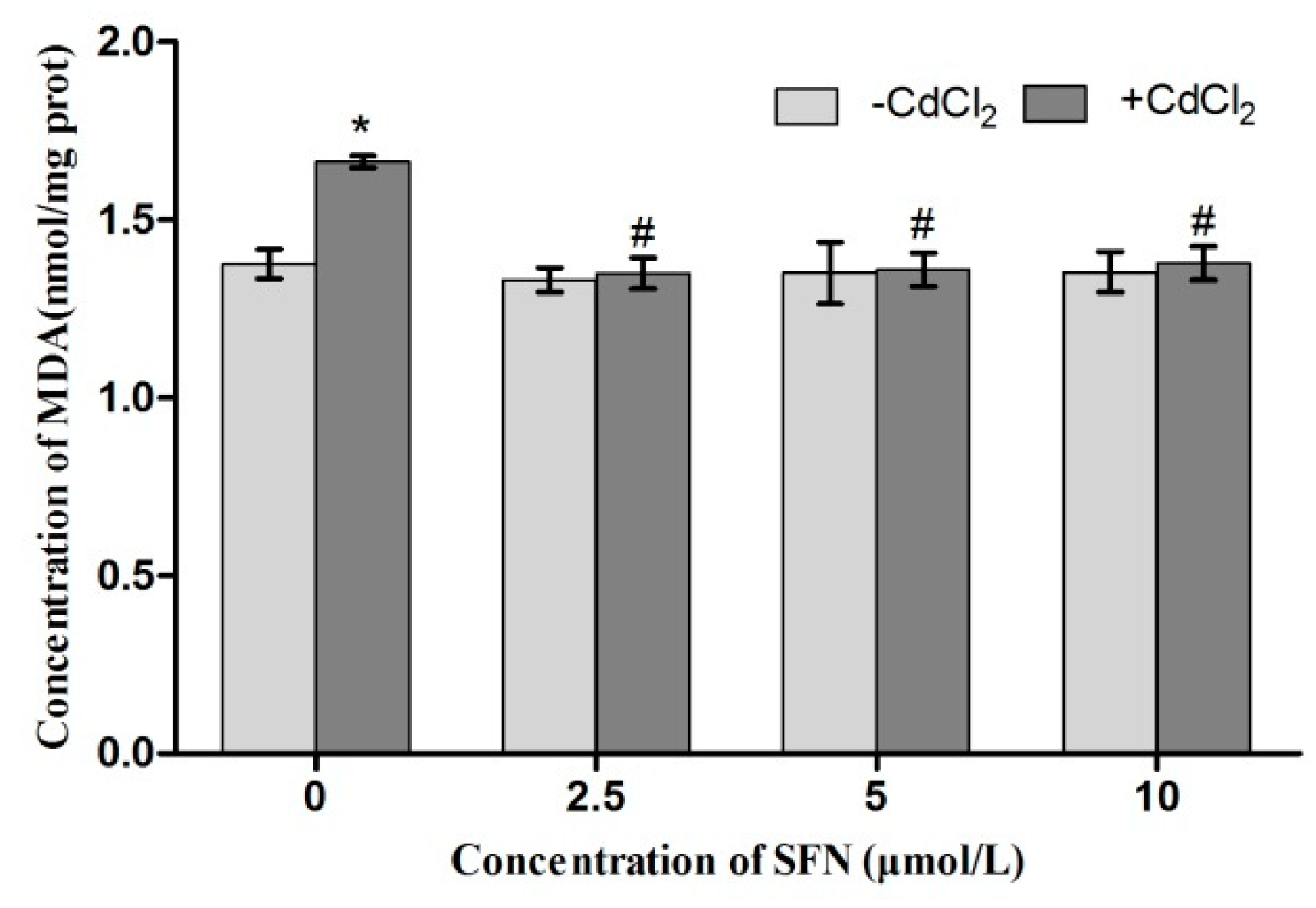
2.8. Detection of Cell Antioxidant Ability
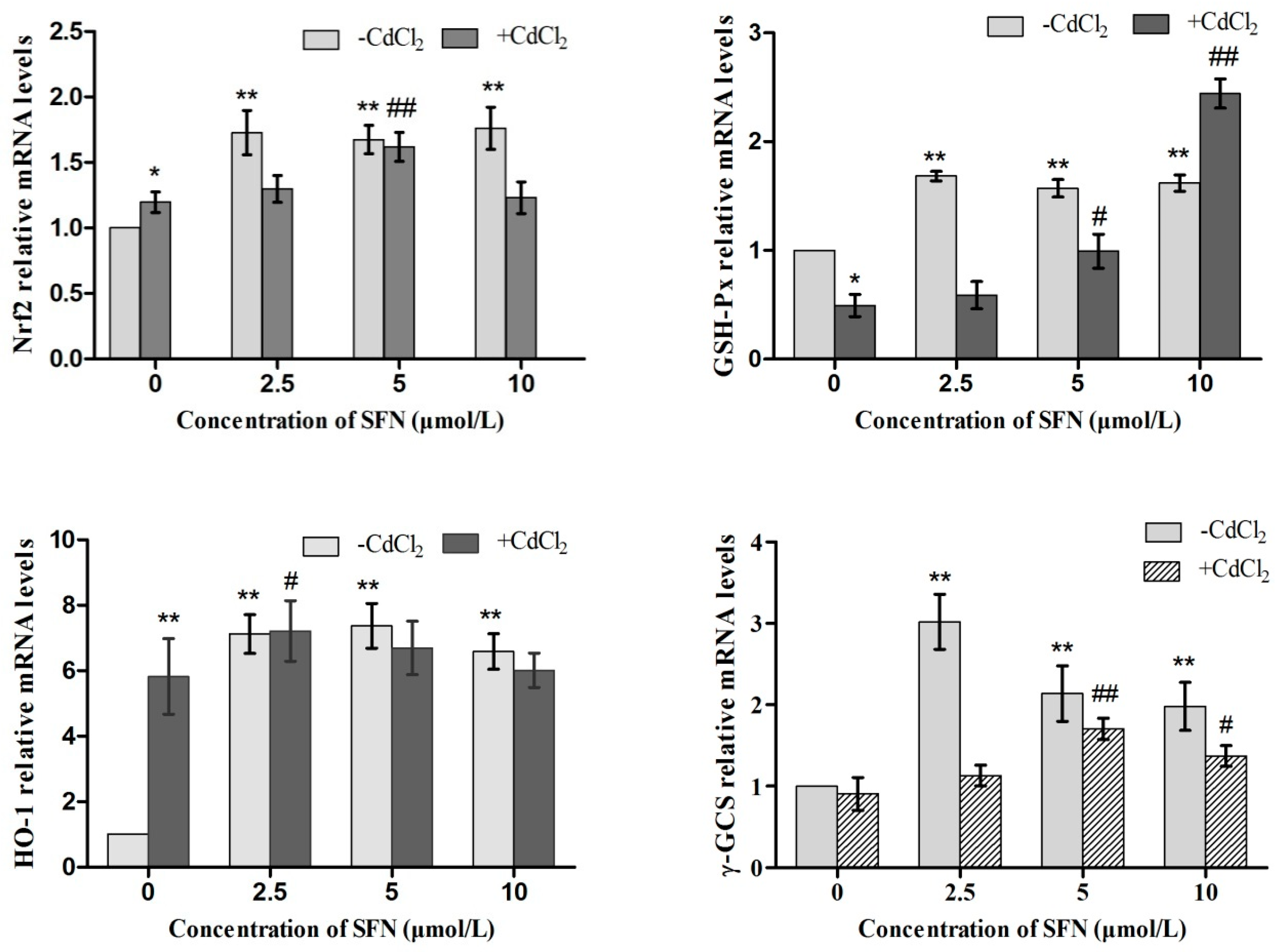
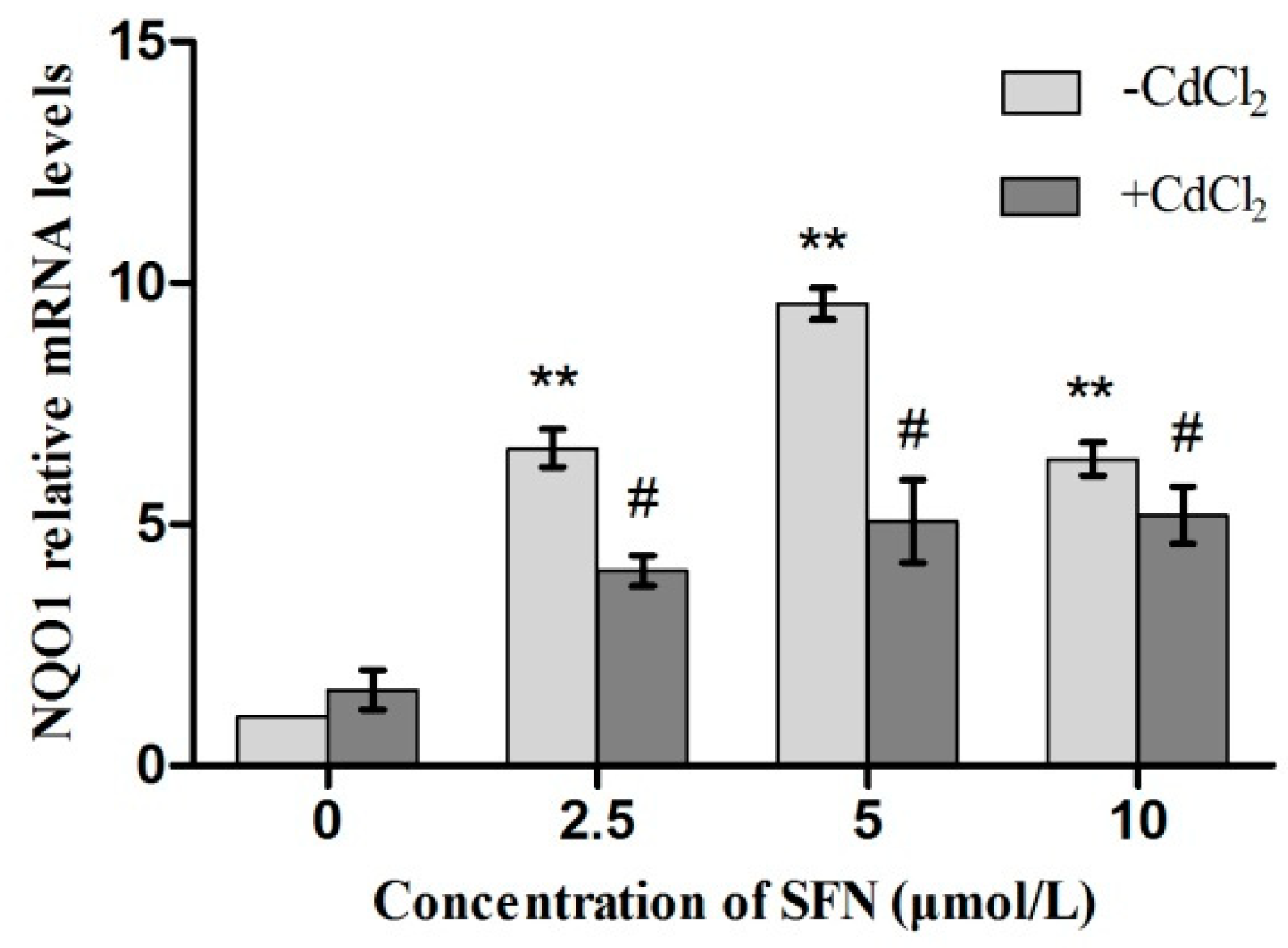
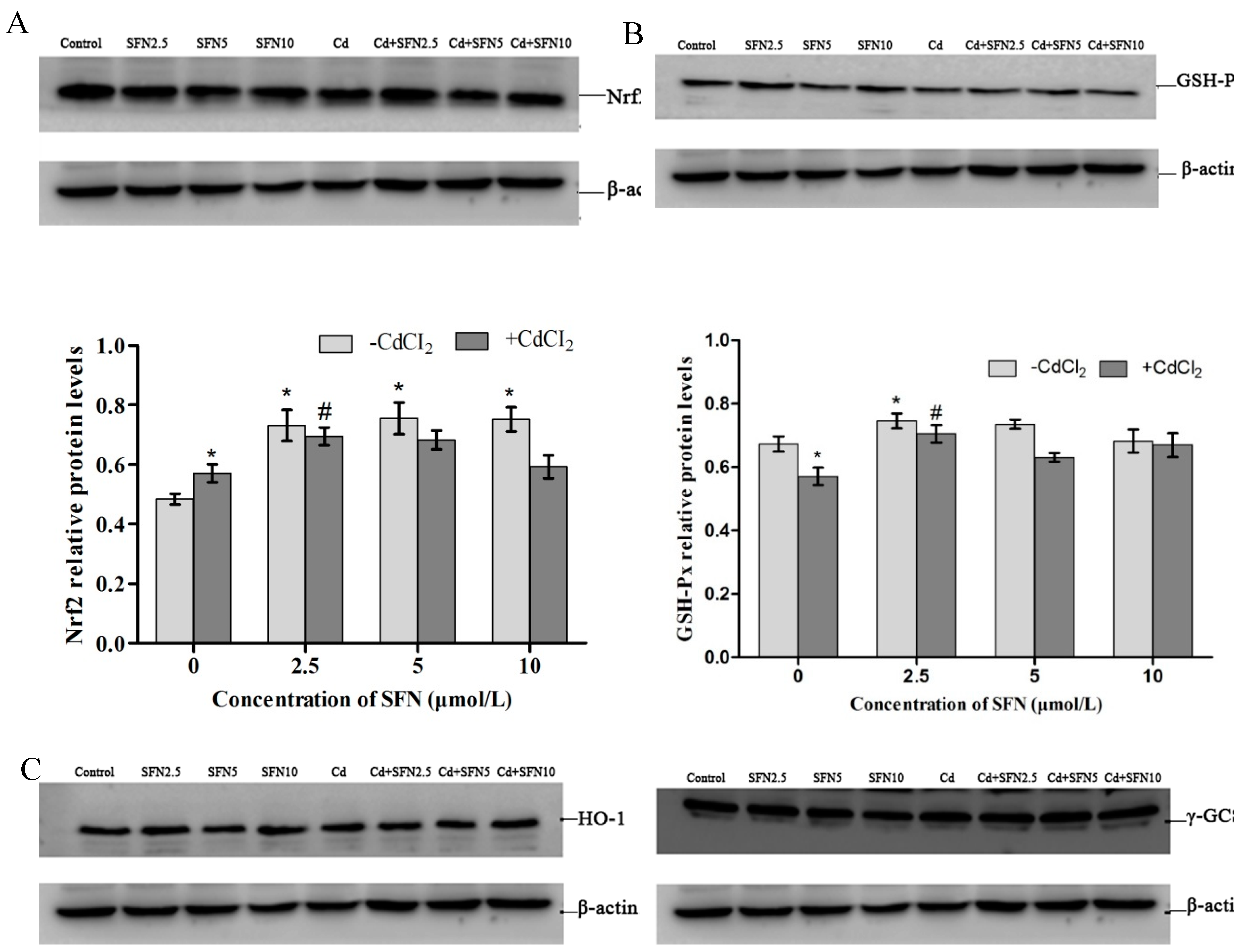
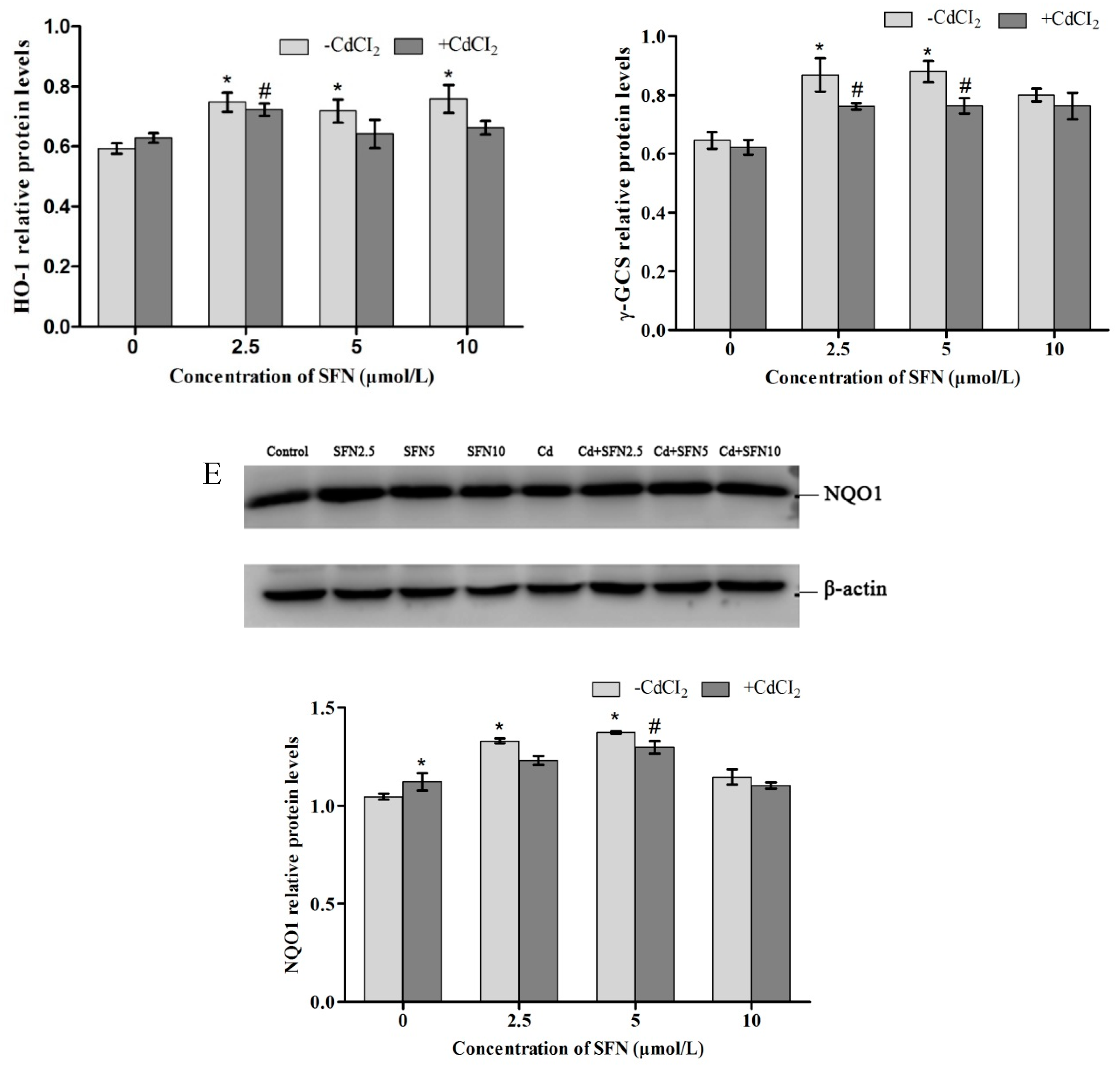
2.9. Expression of Genes Involved in the Nrf2 Pathway
2.10. Expression of Proteins Involved in the Nrf2 Pathway
3. Discussion
4. Materials and Methods
4.1. Treatment of TM3 Cells with Cd and SFN
4.2. Determining the Relative Survival Rate of TM3 Cells
4.3. Determining LDH Activity
4.4. Measuring Testosterone Concentration
4.5. Detecting Antioxidant Ability
4.6. Quantification of Cell Apoptosis
4.7. Expression of Genes Involved in the Nrf2/ARE Signaling Pathway
4.8. Western Blot Analysis
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Zhao, Y.; Li, R.; Lin, Y. Allograft inflammatory factor-1 in grass carp (Ctenopharynogodonidella): Expression and response to cadmium exposure. Fish Shellfish Immunol. 2015, 47, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Laneab, E.A.; Cantyac, M.J.; Morea, S.J. Cadmium exposure and consequence for the health and productivity of farmed ruminants. Res. Vet. Sci. 2015, 101, 132–139. [Google Scholar]
- Zhao, H.; Liu, W.; Wang, Y.; Dai, N.; Gu, J.; Yuan, Y.; Liu, X.; Bian, J.; Liu, Z.P. Cadmium induces apoptosis in primary rat osteoblasts through caspase and mitogen-activated protein kinase pathways. J. Vet. Sci. 2015, 16, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.W.; Zheng, Z.; Cai, J.Z.; Liu, Q.; Yang, J.; Gong, Y.F.; Wu, M.S.; Shen, Q.; Xu, S.W. Effect of cadmium on oxidative stress and immune function of common carp (Cyprinus carpio L.) by transcriptome analysis. Aquat. Toxicol. 2017, 192, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Ognjanović, B.I.; Marković, S.D.; Ethordević, N.Z.; Trbojević, I.S.; Stajn, A.S.; Saicić, Z.S. Cadmium-induced lipid peroxidation and changes in antioxidant defense system in the rat testes: Protective role of coenzyme Q(10) and vitamin E. Reprod. Toxicol. 2010, 29, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Elbaghdady, H.A.M.; Alwaili, M.A.; El-Demerdash, R.S. Amelioration of cadmium-induced testes’ damage in rats by the bone marrow mesenchymalstem cells. Ecotoxicol. Environ. Saf. 2018, 148, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Abdelrazek, H.M.; Helmy, S.A.; Elsayed, D.H.; Ebaid, H.M.; Mohamed, R.M. Ameliorating effects of green tea extract on cadmium induced reproductive injury in male Wistarrats with respect to androgen receptors and caspase-3. Reprod. Biol. 2016, 16, 300–308. [Google Scholar] [CrossRef]
- Lehel, J.; Lanyi, K.; Laczy, P. Food safety significance of heavy metal contamination in food of animal origin. Magy. Allatorv. Lapja 2016, 138, 99–112. [Google Scholar]
- Liu, Q.; Gu, J.H.; Yuan, Y.; Liu, X.Z.; Wang, Y.J.; Wang, H.D.; Liu, Z.P.; Wang, Z.Y.; Bian, J.C. Effect of cadmium on rat Leydig cell testosterone production and DNA integrity in vitro. Biomed. Environ. Sci. 2013, 26, 769. [Google Scholar]
- Farombi, E.O.; Adedara, I.A.; Akinrinde, S.A.; Ojo, O.O.; Eboh, A.S. Protective effects of kolaviron and quercetin on cadmium induced testicular damage and endocrine pathology in rats. Androloqia 2012, 44, 273–284. [Google Scholar] [CrossRef]
- Zhao, L.L.; Ru, Y.F.; Liu, M.; Tang, J.N.; Zheng, J.F.; Wu, B.; Gu, Y.H.; Shi, H.J. Reproductive effects of cadmium on sperm function and early embryonic development in vitro. PLoS ONE 2017, 12, e0186727. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.; Bannigan, J. Cadmium: Toxic effects on the reproductive system and the embryo. Reprod. Toxicol. 2008, 25, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.L. Effects of sulforaphane combined with tea polyphenols on expression of protein kinase A anchorage protein 95 and cyclin E2 in lung cancer tissues. Mod. Pharm. Clin. 2014, 10, 1092–1095. [Google Scholar]
- Bai, Y.; Cui, W.; Xin, Y.; Miao, X.; Barati, M.T.; Zhang, C.; Chen, Q.; Tan, Y.; Cui, T.; Zheng, Y.; et al. Prevention by sulforaphane of diabetic cardiomyopathy is associated with up-regulation of Nrf2 expression and transcription activation. J. Mol. Cell Cardiol. 2013, 57, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, E.; Červinka, M. Sulforaphane induces cytotoxicity and lysosome-and mitochondria-dependent cell death in colon cancer cells with deleted p53. Toxicol. In Vitro 2011, 25, 1302–1309. [Google Scholar] [CrossRef]
- de Figueiredo, S.M.; Binda, N.S.; Nogueira-Machado, J.A.; Vieira-Filho, S.A.; Caligiorne, R.B. The antioxidant properties of organosulfur compounds (sulforaphane). Recent Pat. Endocr. Metab. Immune Drug Discov. 2015, 9, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Wehage, S.L.; Holtzclaw, W.D.; Kensler, T.W.; Egner, P.A.; Shapiro, T.A.; Talalay, P. Protection of Humans by Plant Glucosinolates: Efficiency of Conversion of Glucosinolates to Isothiocyanates by the Gastrointestinal Microflora. Cancer Prev. Res. 2012, 5, 603–611. [Google Scholar] [CrossRef]
- Guerrero-Beltrán, C.E.; Mukhopadhyay, P.; Horváth, B.; Rajesh, M.; Tapia, E.; García-Torres, I.; Pedraza-Chaverri, J.; Pacher, P. Sulforaphane, a natural constituent of broccoli, prevents cell death and inflammation in nephropathy. J. Nutr. Biochem. 2012, 23, 494–500. [Google Scholar] [CrossRef]
- Hlubocky, A.; Wellik, K.; Ross, M.A.; Smith, B.E.; Hoffman-Snyder, C.; Demaerschalk, B.M.; Wingerchuk, D.M. Skin biopsy for diagnosis of small fiber neuropathy: A critically appraised topic. Neurologist 2010, 16, 61–63. [Google Scholar] [CrossRef]
- Li, J.H.; Xu, Z.F.; Li, S.S.; Yang, T.Y.; Liu, W.; Li, Y.H.; Feng, S. Protective effect of sulforaphane on renal damage caused by cadmium in rats. Chin. J. Ind. Med. 2015, 8, 11–15. [Google Scholar]
- Shinkai, Y.; Sumi, D.; Fukami, I.; Ishii, T.; Kumagai, Y. Sulforaphane, an activator of Nrf2, suppresses cellular accumulation of arsenic and its cytotoxicity in primary mouse hepatocytes. FEBS Lett. 2006, 580, 1771–1774. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.F.; Li, Z.; Zhang, H.Q.; Wang, A.S.; Yang, L.F.; Wang, X.; Zhang, X.H. Research progress in regulation of sulforaphane on metabolic diseases. Chin. J. Pharmacol. Toxicol. 2015, 3, 477–481. [Google Scholar]
- Yang, S.H.; Long, M.; Yu, L.H.; Li, L.; Li, P.; Zhang, Y.; Guo, Y.; Gao, F.; Liu, M.D.; He, J.B. Sulforaphane Prevents Testicular Damage in Kunming Mice Exposed to Cadmium via Activation of Nrf2/ARE Signaling Pathways. Int. J. Mol. Sci. 2016, 17, 1703. [Google Scholar] [CrossRef] [PubMed]
- Satarug, S.; Baker, J.R.; Urbenjapol, S.; Haswell-Elkins, M.; Reilly, P.E.B.; Williams, D.J.; Moore, M.R. A global perspective on cadmium pollution and toxicity in nonoccupationally exposed population. Toxicol. Lett. 2003, 137, 65–83. [Google Scholar] [CrossRef]
- Nordberg, G.F. Historical perspectives on cadmium toxicology. Toxicol. Appl. Pharmcol. 2009, 238, 192–200. [Google Scholar] [CrossRef]
- Jin, X.; Jia, T.; Liu, R.; Xu, S. The antagonistic effect of selenium on cadmium-induced apoptosis via PPAR-γ/PI3K/Akt pathway in chicken pancreas. J. Hazard. Mater. 2018, 357, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Yang, SH.; Zhang, Z.W.; He, J.B.; Li, J.L.; Zhang, J.L.; Xing, H.J.; Xu, S.W. Ovarian Toxicity Induced by Dietary Cadmium in Hen. Biol. Trace Elem. Res. 2012, 148, 53–60. [Google Scholar] [CrossRef]
- Adamkovicova, M.; Toman, R.; Martiniakova, M.; Omelka, R.; Babosova, R.; Krajcovicova, V.; Grosskopf, B.; Massanyi, P. Sperm motility and morphology changes in rats exposed to cadmium and diazinon. Reprod. Biol. Endocrinol. 2016, 14, 1–7. [Google Scholar] [CrossRef]
- Li, R.Y.; Luo, X.; Li, L.B.; Peng, Q.; Yang, Y.Y.; Zhao, L.T.; Ma, M.F.; Hou, Z.W. The Protective Effects of Melatonin Against Oxidative Stress and Inflammation Induced by Acute Cadmium Exposure in Mice Testis. Biol. Trace Elem. Res. 2016, 170, 152–164. [Google Scholar] [CrossRef]
- Hayes, J.D.; Kelleher, M.O.; Eggleston, I.M. The cancer actions of phytochemicals derived from chemopreventive glucosinolates. Eur. J. Nutr. 2008, 47, 73–88. [Google Scholar] [CrossRef]
- Briones-Herrera, A.; Eugenio-Pérez, D.; Reyes-Ocampo, J.G.; Rivera-Mancía, S.; Pedraza-Chaverri, J. New highlights on the health-improving effects of sulforaphane. Food Funct. 2018, 9, 2589–2606. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.M.; Wang, G.G.; Xu, D.Q.; Luo, K.; Liu, Y.X.; Zhong, Y.H.; Cai, Y.Q. The protection of selenium on cadmium-induced inhibition of spermatogenesis via activating testosterone synthesis in mice. Food Chem. Toxicol. 2012, 50, 3521–3529. [Google Scholar] [CrossRef] [PubMed]
- Guillouzo, A.; Uguen-Guillouzo, C. In Vitro Methods in Toxicology; Academic Press: New York, NY, USA, 1992. [Google Scholar]
- Alkharashi, N.A.O.; Periasamy, V.S.; Athinarayanan, J.; Alshatwi, A.A. Cadmium triggers mitochondrial oxidative stress in human peripheral blood lymphocytes and monocytes: Analysis using in vitro and system toxicology approaches. J. Trace Elem. Med. Biol. 2017, 42, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Alkharashi, N.A.O.; Periasamy, V.S.; Athinarayanan, J.; Alshatwi, A.A. Assessment of sulforaphane-induced protective mechanisms against cadmium toxicity in human mesenchymal stem cells. Environ. Sci. Pollut. Res. 2018, 25, 10080–10089. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Gallagher, E.P. Role of Nrf2 antioxidant defense in mitigating cadmium-induced oxidative stress in the olfactory system of zebrafish. Toxicol. Appl. Pharm. 2013, 266, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Bashir, N.; Manoharan, V.; Miltonprabu, S. Grape seed proanthocyanidins protects against cadmium induced oxidative pancreatitis in rats by attenuating oxidative stress, inflammation and apoptosis via Nrf-2/HO-1 signaling. J. Nutr. Biochem. 2016, 32, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Li, Q.; Zheng, G.L.; Chen, Y.X.; Huang, M.C.; Zhang, L.J.; Lin, X.Y. Protective effect of Lycium barbarum polysaccharides against cadmium-induced testicular toxicity in male mice. Food Funct. 2017, 8, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, R.; Song, Y.; Li, T.; Ge, M. Protective Effect of Ganoderma Triterpenoids on Cadmium-Induced Testicular Toxicity in Chickens. Biol. Trace Elem. Res. 2019, 187, 281–290. [Google Scholar] [CrossRef]
- Oh, C.J.; Kim, J.Y.; Min, A.K.; Park, K.G.; Harris, R.A.; Kim, H.J.; Lee, I.K. Sulforaphane attenuates hepatic fibrosis via NF-E2-related factor 2-mediated inhibition of transforming growth factor-β/smad signaling. Free Radic. Biol. Med. 2012, 52, 671–682. [Google Scholar] [CrossRef]
- Lu, M.C.; Ji, J.A.; Jiang, Z.Y.; You, Q.D. The Keap1-Nrf2-ARE pathway as a potential preventive and therapeutic target: An update. Med. Res. Rev. 2016, 36, 924–963. [Google Scholar] [CrossRef]
- Tao, S.; Justiniano, R.; Zhang, D.D.; Wondrak, G.T. The Nrf2-inducers tanshinone I and dihydrotanshinone protect human skin cells and reconstructed human skin against solar simulated UV. Redox Biol. 2013, 29, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Wiwanitkit, V. Cadmium related health effects. Environ. Res. 2012, 236, 237–238. [Google Scholar] [CrossRef] [PubMed]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. | Primer Sequence (5′–3′) | Product Length |
|---|---|---|---|
| Nrf2 | NM_010902.3 | Forward: TCCTATGCGTGAATCCCAAT | 103 bp |
| Reverse: GCGGCTTGAATGTTTGTCTT | |||
| GSH-PX | X03920.1 | Forward: GAAGTGCGAAGTGAATGG | 224 bp |
| Reverse: TGTCGATGGTACGAAAGC | |||
| HO-1 | NM_010442.2 | Forward: GGGCTGTGAACTCTGTCCAAT | 162 bp |
| Reverse: GGTGAGGGAACTGTGTCAGG | |||
| γ-GCS | U85414.1 | Forward: TGGATGATGCCAACGAGTC | 185 bp |
| Reverse: CCTAGTGAGCAGTACCACGAATA | |||
| NQO1 | NM_008706.5 | Forward: TTCTGTGGCTTCCAGGTCTT | 104 bp |
| Reverse: TCCAGACGTTTCTTCCATCC | |||
| β-actin | BC138614.1 | Forward: CTGTCCCTGTATGCCTCTG | 221 bp |
| Reverse: TTGATGTCACGCACGATT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.-H.; Li, P.; Yu, L.-H.; Li, L.; Long, M.; Liu, M.-D.; He, J.-B. Sulforaphane Protect Against Cadmium-Induced Oxidative Damage in mouse Leydigs Cells by Activating Nrf2/ARE Signaling Pathway. Int. J. Mol. Sci. 2019, 20, 630. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030630
Yang S-H, Li P, Yu L-H, Li L, Long M, Liu M-D, He J-B. Sulforaphane Protect Against Cadmium-Induced Oxidative Damage in mouse Leydigs Cells by Activating Nrf2/ARE Signaling Pathway. International Journal of Molecular Sciences. 2019; 20(3):630. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030630
Chicago/Turabian StyleYang, Shu-Hua, Peng Li, Li-Hui Yu, Lin Li, Miao Long, Ming-Da Liu, and Jian-Bin He. 2019. "Sulforaphane Protect Against Cadmium-Induced Oxidative Damage in mouse Leydigs Cells by Activating Nrf2/ARE Signaling Pathway" International Journal of Molecular Sciences 20, no. 3: 630. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20030630