Molecular Implications of Natriuretic Peptides in the Protection from Hypertension and Target Organ Damage Development

Abstract
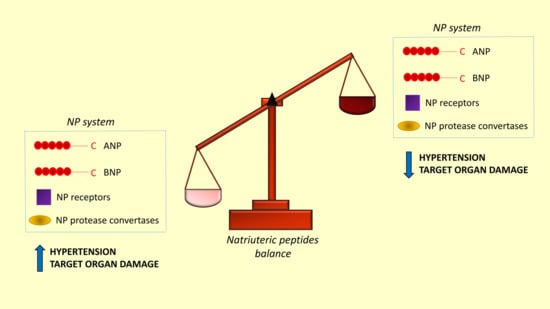
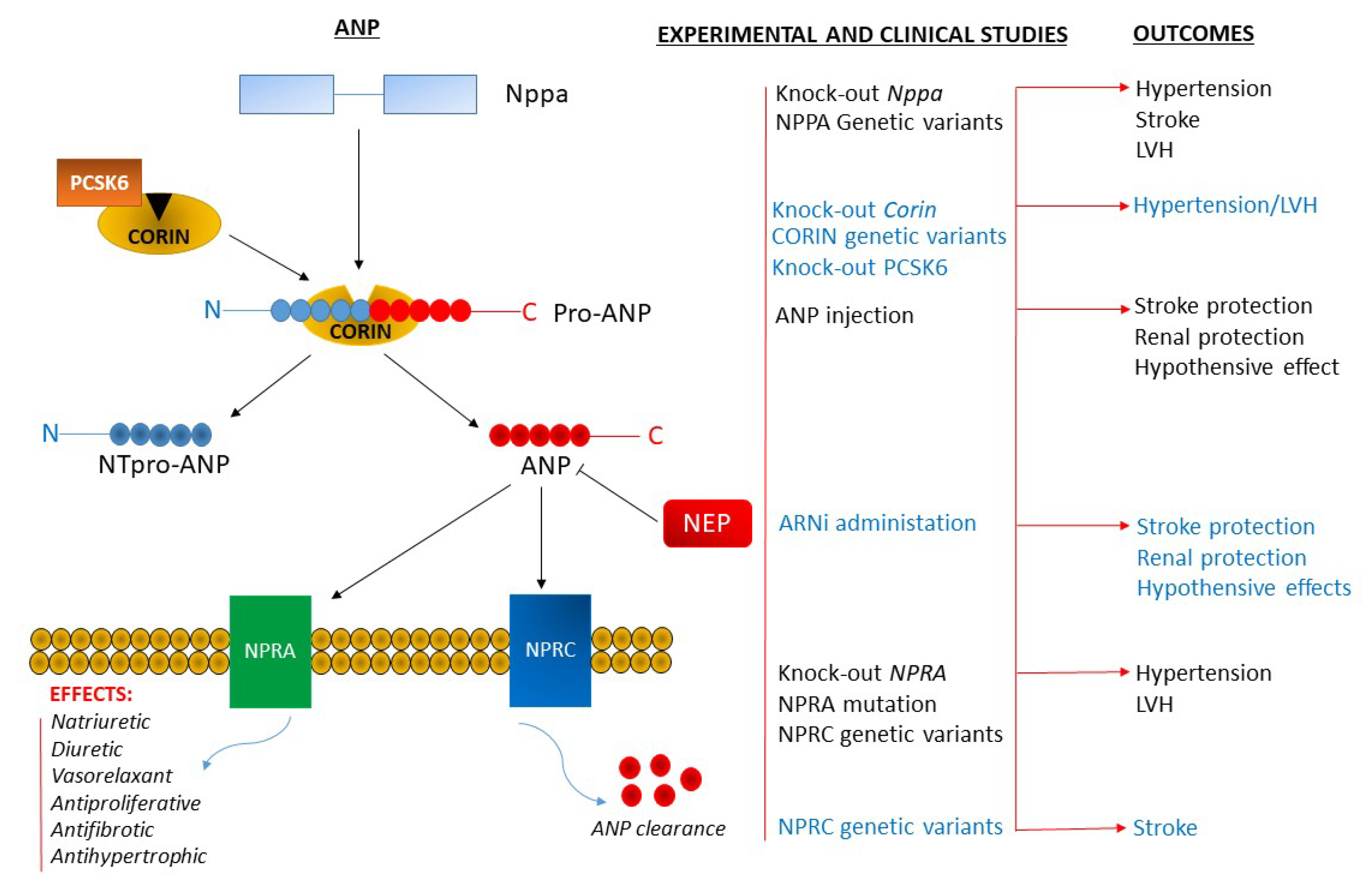
:
{kind=link}
{kind=link}
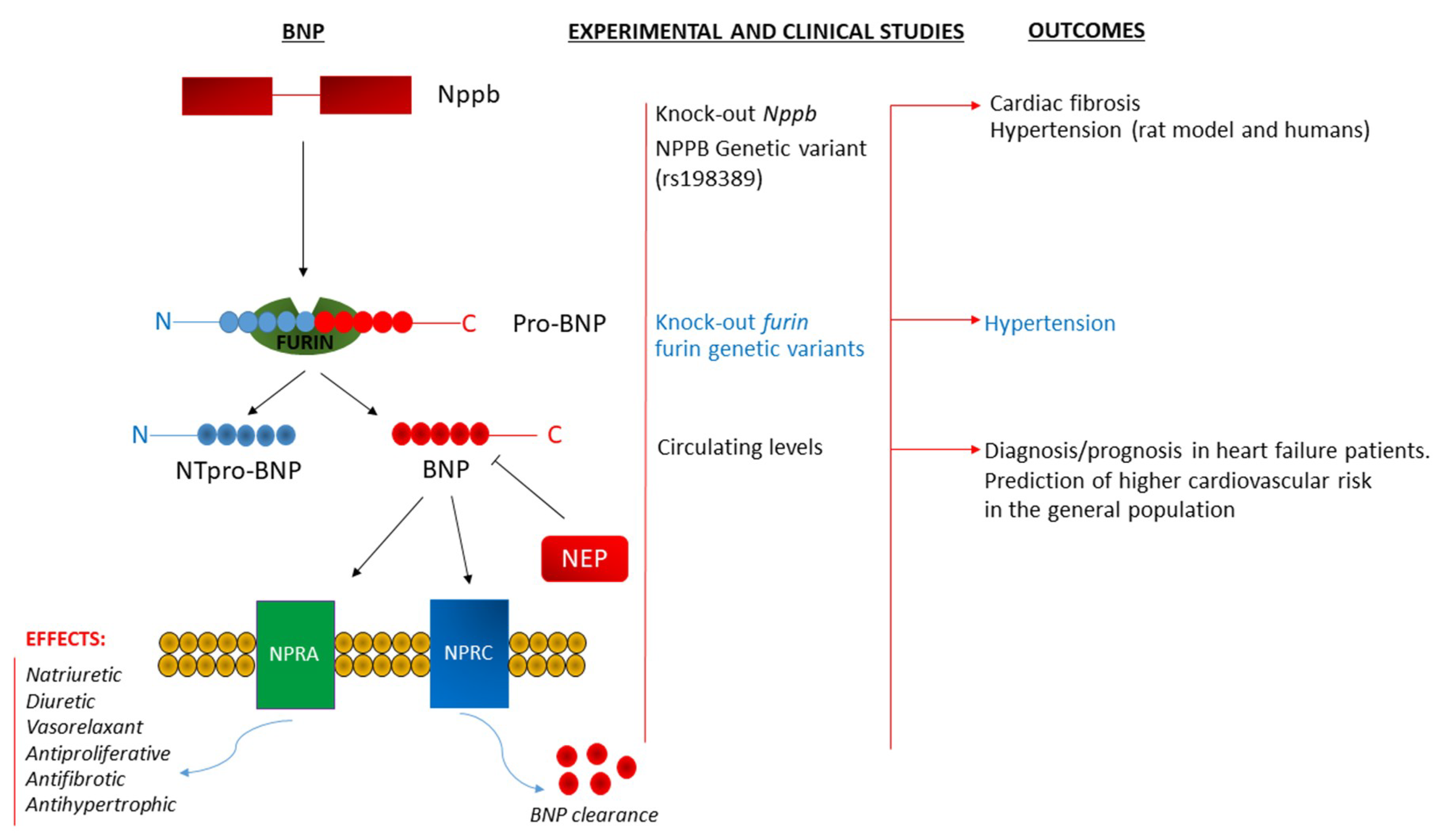{kind=link}
1. Introduction
2. Role of NP Circulating Levels in the Development of High Blood Pressure
3. Role of Other Components of the NP Family
4. Impact of NPs on Predisposition to Develop Hypertensive Target Organ Damage
5. NP-Based Therapies for the Treatment of Hypertension and Related Organ Damage
Funding
Conflicts of Interest
References
- Kunes, J.; Zicha, J. The interaction of genetic and environmental factors in the etiology of hypertension. Physiol. Res. 2009, 58 (Suppl. 2), S33–S41. [Google Scholar]
- Levin, E.R.; Gardner, D.G.; Samson, W.K. Natriuretic peptides. NEJM 1998, 339, 321–328. [Google Scholar] [PubMed]
- Volpe, M.; Rubattu, S.; Burnett, J., Jr. Natriuretic peptides in cardiovascular diseases: Current use and perspectives. Eur. Heart J. 2014, 35, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Volpe, M.; De Luca, N.; Indolfi, C.; Mele, A.F.; Rivera, M.P.; Condorelli, M.; Trimarco, B. Acute hemodynamic effects of atrial natriuretic peptide in essential hypertension. Kidney Int. Suppl. 1998, 25, 583–585. [Google Scholar]
- Ishii, M.; Sugimoto, T.; Matsuoka, H.; Hirata, Y.; Ishimitsu, T.; Fukul, K.; Sugimoto, T.; Kanagawa, K.; Matsuo, H. A comparative study on the hemodynamic, renal and endocrine effects of alpha-human atrial natriuretic polypeptide in normotensive persons and patients with essential hypertension. Jpn. Circ. J. 1986, 50, 1181–1184. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, T.; Ishibashi, M.; Sekihara, H.; Takaku, F.; Nakaoka, H.; Fujii, J. Plasma levels of atrial natriuretic peptide in primary aldosteronism and essential hypertension. J. Clin. Endocrinol. Metab. 1986, 63, 815–818. [Google Scholar] [CrossRef] [PubMed]
- Stepniakowski, K.; Januszewicz, A.; Lapiński, M.; Feltynowski, T.; Chodakowska, J.; Ignatowska-Switalska, H.; Wocial, B.; Januszewicz, W. Plasma atrial natriuretic peptide (ANP) concentration in patients with pheochromocytoma. Blood Press 1992, 1, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Vesely, D.L. Atrial natriuretic peptide prohormone gene expression: Hormones and diseases that upregulate its expression. IUBMB Life 2002, 53, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Miyata, M.; Sasaki, T.; Ikeda, Y.; Shinsato, T.; Kubozono, T.; Furusho, Y.; Kusumoto, A.; Hamasaki, S.; Tei, C. COLD-CHF Investigators. Comparative study of therapeutic effects of short- and long-acting loop diuretics in outpatients with chronic heart failure (COLD-CHF). J. Cardiol. 2012, 59, 352–358. [Google Scholar] [CrossRef]
- John, S.W.; Krege, J.H.; Oliver, P.M.; Hagaman, J.R.; Pang, S.C.; Flynn, T.G.; Smithies, O. Genetic decreases in atrial natriuretic peptide and salt-sensitive hypertension. Science 1995, 267, 679–681. [Google Scholar] [CrossRef]
- Steinhelper, M.E.; Cochrane, K.L.; Filed, L.J. Hypotension in transgenic mice expressing atrial natriuretic factor fusion genes. Hypertension 1990, 16, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Oliver, P.M.; Fox, J.E.; Kim, R.; Rockman, H.A.; Kim, H.S.; Reddick, R.L.; Pandey, K.N.; Milgram, S.L.; Smithies, O.; Maeda, N. Hypertension, cardiac hypertrophy, and sudden death in mice lacking natriuretic peptide receptor A. Proc. Natl. Acad. Sci. USA 1997, 94, 14730–14735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichiki, T.; Huntley, B.K.; Burnett, J., Jr. BNP molecular forms and processing by the cardiac serine protease corin. Adv. Clin. Chem. 2013, 61, 1–31. [Google Scholar] [PubMed]
- Tamura, N.; Ogawa, Y.; Chusho, H.; Nakamura, K.; Nakao, K.; Suda, M.; Kasahara, M.; Hashimoto, R.; Katsuura, G.; Mukoyama, M.; et al. Cardiac fibrosis in mice lacking brain natriuretic peptide. Proc. Natl. Acad. Sci. USA 2000, 97, 4239–4244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holditch, S.J.; Schreiber, C.A.; Nini, R.; Tonne, J.M.; Peng, K.W.; Geurts, A.; Jacob, H.J.; Burnett, J.C.; Cataliotti, A.; Ikeda, Y. B-type natriuretic peptide deletion leads to progressive hypertension, associated organ damage, and reduced survival: Novel model for human hypertension. Hypertension 2015, 66, 199–210. [Google Scholar] [PubMed]
- Rubattu, S.; Evangelista, A.; Barbato, D.; Barba, G.; Stanzione, R.; Iacone, R.; Volpe, M.; Strazzullo, P. Atrial natriuretic peptide (ANP) gene promoter variant and increased susceptibility to early development of hypertension in humans. J. Hum. Hypertens. 2007, 21, 822–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, N.; Sugiyama, T.; Morita, H.; Nabika, T.; Kurihara, H.; Yamori, Y.; Yazaki, Y. Genetic analysis of the atrial natriuretic peptide gene in essential hypertension. Clin. Sci. 2000, 98, 251–258. [Google Scholar] [CrossRef]
- Conen, D.; Glynn, R.J.; Buring, J.E.; Ridker, P.M.; Zee, R.Y. Natriuretic peptide precursor A gene polymorphisms and risk of blood pressure progression and incident hypertension. Hypertension 2007, 50, 1114–1119. [Google Scholar] [CrossRef]
- Zhang, S.; Mao, G.; Zhang, Y.; Tang, G.; Wen, Y.; Hong, X.; Jiang, S.; Yu, Y.; Xu, X. Association between human atrial natriuretic peptide Val7Met polymorphism and baseline blood pressure, plasma trough irbesartan concentrations, and the antihypertensive efficacy of irbesartan in rural Chinese patients with essential hypertension. Clin. Ther. 2005, 27, 1774–1784. [Google Scholar] [CrossRef]
- Beige, J.; Ringel, J.; Hohenbleicher, H.; Rubattu, S.; Kreutz, R.; Sharma, A.M. HpaII-polymorphism of the atrial natriuretic peptide gene and essential hypertension in whites. Am. J. Hypertens. 1997, 10, 1316–1318. [Google Scholar]
- Newton-Cheh, C.; Johnson, T.; Gateva, V.; Tobin, M.D.; Bochud, M.; Coin, L.; Najjar, S.S.; Zhao, J.H.; Heath, S.C.; Eyheramendy, S.; et al. Genome-wide association study identifies eight loci associated with blood pressure. Nat. Genet. 2009, 41, 666–676. [Google Scholar] [CrossRef] [Green Version]
- Pereira, N.L.; Tosakulwong, N.; Scott, C.G.; Jenkins, G.D.; Prodduturi, N.; Chai, Y.; Olson, T.M.; Rodeheffer, R.J.; Redfield, M.M.; Weinshilboum, R.M.; et al. Circulating atrial natriuretic peptide genetic association study identifies a novel gene cluster associated with stroke in whites. Circ. Cardiovasc. Genet. 2015, 8, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Arora, P.; Wu, C.; Khan, A.M.; Bloch, D.B.; Davis-Dusenbery, B.N.; Ghorbani, A.; Spagnolli, E.; Martinez, A.; Ryan, A.; Tainsh, L.T.; et al. Atrial natriuretic peptide is negatively regulated by microRNA-425. J. Clin. Investig. 2013, 123, 3378–3382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannone, V.; Boerrigter, G.; Cataliotti, A.; Costello-Boerrigter, L.C.; Olson, T.M.; McKie, P.M.; Heublein, D.M.; Lahr, B.D.; Bailey, K.R.; Averna, M.; et al. A genetic variant of the atrial natriuretic peptide gene is associated with cardiometabolic protection in the general community. J. Am. Coll. Cardiol. 2011, 58, 629–636. [Google Scholar] [CrossRef]
- Cannone, V.; Cefalu’, A.B.; Noto, D.; Scott, C.G.; Bailey, K.R.; Cavera, G.; Pagano, M.; Sapienza, M.; Averna, M.R.; Burnett, J.C., Jr. The atrial natriuretic peptide genetic variant rs5068 is associated with a favorable cardiometabolic phenotype in a Mediterranean population. Diabetes Care 2013, 36, 2850–2856. [Google Scholar] [CrossRef] [PubMed]
- Rubattu, S.; Sciarretta, S.; Volpe, M. Atrial natriuretic peptide gene variants and circulating levels: Implications in cardiovascular diseases. Clin. Sci. 2014, 127, 1–13. [Google Scholar] [CrossRef]
- Musani, S.K.; Fox, E.R.; Kraja, A.; Bidulescu, A.; Lieb, W.; Lin, H.; Beecham, A.; Chen, M.H.; Felix, J.F.; Fox, C.S.; et al. Genome-wide association analysis of B-type natriuretic peptide in blacks: The Jacson Heart Study. Circ. Cardiovasc. Genet. 2015, 8, 122–130. [Google Scholar] [CrossRef]
- Johansson, A.; Eriksson, N.; Lindholm, D.; Varenhorst, C.; James, S.; Syvanen, A.C.; Axelsson, T.; Siegbahn, A.; Barratt, B.J.; Becker, R.C.; et al. Genome-wide association and Mendelian randomization study of NT-proBNP in patients with acute coronary syndrome. Hum. Mol. Genet. 2016, 25, 1447–1456. [Google Scholar] [CrossRef] [Green Version]
- Del Greco, M.F.; Pattaro, C.; Luchner, A.; Pichler, I.; Winkler, T.; Hicks, A.A.; Fuchsberger, C.; Franke, A.; Melville, S.A.; Peters, A.; et al. Genome-wide association analysis and fine mapping of NT-proBNP level provide novel insight into the role of the MTHFR-CLCN6-NPPA-NPPB gene cluster. Hum. Mol. Genet. 2011, 20, 1660–1671. [Google Scholar] [Green Version]
- Folkersen, L.; Fauman, E.; Sabater-Lleal, M.; Strawbridge, R.J.; Franberg, M.; Sennblad, B.; Baldassarre, D.; Veglia, F.; Humphries, S.E.; Rauramaa, R.; et al. Mapping of 79 loci for 83 plasma protein biomarkers in cardiovascular disease. PLoS Genet. 2017, 13, e1006706. [Google Scholar] [CrossRef]
- Salo, P.P.; Havulinna, A.S.; Tukiainen, T.; Raitakari, O.; Lehtimaki, T.; Kahonen, M.; Kettunen, J.; Männikkö, M.; Eriksson, J.G.; Jula, A.; et al. Genome-Wide Association Study implicates atrial natriuretic peptide rather than B-type natriuretic peptide in the regulation of blood pressure in the general population. Circ. Cardiovasc. Genet. 2017, 10, e001713. [Google Scholar] [CrossRef] [PubMed]
- Lanfear, D.E.; Stolker, J.M.; Marsh, S.; Rich, M.W.; McLeod, H.L. Genetic variation in the B-type natriuretic peptide pathway affects BNP levels. Cardiovasc. Drugs Ther. 2007, 21, 55–62. [Google Scholar] [CrossRef]
- Seidelmann, S.B.; Vardeny, O.; Claggett, B.; Yu, B.; Shah, A.M.; Ballantyne, C.M.; Selvin, E.; MacRae, C.A.; Boerwinkle, E.; Solomon, S.D. An NPPB promoter polymorphism associated with elevated N-terminal pro-B-type natriuretic peptide and lower blood pressure, hypertension, and mortality. J. Am. Heart Assoc. 2017, 6, e005257. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Wu, F.; Morser, J.; Wu, Q. Corin, a transmembrane cardiac serine protease, acts as a pro-atrial natriuretic peptide-converting enzyme. Proc. Natl. Acad. Sci. USA 2000, 97, 8525–8529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Yan, W.; Pan, J.; Morser, J.; Wu, Q. Processing of pro-atrial natriuretic peptide by corin in cardiac myocytes. J. Biol. Chem. 2002, 277, 16900–16905. [Google Scholar] [CrossRef]
- Chan, J.C.; Knudson, O.; Wu, F.; Morser, J.; Dole, W.P.; Wu, Q. Hypertension in mice lacking the proatrial natriuretic peptide convertase corin. Proc. Natl. Acad. Sci. USA 2005, 102, 785–790. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Cao, P.; Dong, N.; Peng, J.; Zhang, C.; Wang, H.; Zhou, T.; Yang, J.; Zhang, Y.; Martelli, E.E.; et al. PCSK6-mediated corin activation is essential for normal blood pressure. Nat. Med. 2015, 21, 1048–1053. [Google Scholar] [CrossRef] [Green Version]
- Volpe, M.; Rubattu, S. Novel insights into the mechanisms regulating pro-atrial natriuretic peptide cleavage in the heart and blood pressure regulation. Proprotein Convertase Subtilisin/Kexin 6 is the corin activating enzyme. Circ. Res. 2016, 118, 196–198. [Google Scholar] [CrossRef]
- Rame, J.E.; Drazner, M.H.; Post, W.; Peshock, R.; Lima, J.; Cooper, R.S.; Dries, D.L. Corin I555 (P568) is associated with enhanced cardiac hypertrophy response to increased systemic afterload. Hypertension 2007, 49, 857–864. [Google Scholar] [CrossRef]
- Rame, J.E.; Tam, S.W.; McNamara, D.; Worcel, M.; Sabolinski, M.L.; Wu, A.H.; Dries, D.L. Dysfunctional corin I555(P568) allele is associated with impaired brain natriuretic peptide processing and advserse outcomes in blacks with systolic heart failure: Results from the genetic risk assessment in heart failure substudy. Circ. Heart Fail. 2009, 2, 541–548. [Google Scholar] [CrossRef]
- Dong, N.; Zhou, T.; Zhang, Y.; Liu, M.; Li, H.; Huang, X.; Liu, Z.; Wu, Y.; Fukuda, K.; Qin, J.; et al. Corin mutations K317E and S472G from preeclamptic patients alter zymogen activation and cell surface targeting. J. Biol. Chem. 2014, 289, 17909–17916. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, T.; Niu, Y.; He, M.; Wang, C.; Liu, M.; Yang, J.; Zhang, Y.; Zhou, J.; Fukuda, K.; et al. Identification and functional analysis of CORIN variants in hypertensive patients. Hum. Mutat. 2017, 38, 1700–1710. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Luo, W.; Juhong, Z.; Yang, J.; Wang, H.; Zhou, L.; Chang, J. Associations between genetic variations in the FURIN gene and hypertension. BMC Med. Genet. 2010, 11, 124. [Google Scholar] [CrossRef] [PubMed]
- International Consortium for Blood Pressure Genome-Wide Association; Eehret, G.B.; Munroe, P.B.; Rice, K.M.; Bochud, M.; Johnson, A.D.; Chasman, D.I.; Smith, A.V.; Tobin, M.D.; Verwoert, G.C.; et al. Genetic variants in novel pathways influence blood pressure and cardiovascular disease risk. Nature 2011, 478, 103–109. [Google Scholar] [Green Version]
- Zhang, H.; Mo, X.B.; Xu, T.; Bu, X.Q.; Lei, S.F.; Zhang, Y.H. Novel genes affecting blood pressure detected via gene-based association analysis. G3 2015, 5, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Soma, M.; Takahashi, Y.; Rehemudula, D.; Kanmatsuse, K.; Furuya, K. Functional deletion mutation of the 5’-flanking region of type A human natriuretic peptide receptor gene and its association with essential hypertension and left ventricular hypertrophy in the Japanese. Circ. Res. 2000, 86, 841–845. [Google Scholar] [CrossRef]
- Sun, H.; Yang, Z.Q.; Liu, S.Y.; Yu, L.; Huang, K.; Lin, K.Q. Correlation between natriuretic peptide receptor C (NPR3) gene polymorphisms and hypertension in the Dai people of China. Genet. Mol. Res. GMR 2015, 14, 8786–8795. [Google Scholar] [CrossRef]
- Pitzalis, M.V.; Sarzani, R.; Dessi-Fulgheri, P.; Iacoviello, M.; Forleo, C.; Lucarelli, K.; Pietrucci, F.; Salvi, F.; Sorrentino, S.; Romito, R.; et al. Allelic variants of natriuretic peptide receptor genes are associated with family history of hypertension and cardiovascular phenotype. J. Hypertens. 2003, 21, 1491–1496. [Google Scholar] [CrossRef]
- Ren, M.; Ng, F.L.; Warren, H.R.; Witkowska, K.; Baron, M.; Jia, Z.; Cabrera, C.; Zhang, R.; Mifsud, B.; Munroe, P.B.; et al. The biological impact of blood pressure-associated genetic variants in the natriuretic peptide receptor C gene on human vascular smooth muscle. Hum. Mol. Genet. 2018, 27, 199–210. [Google Scholar]
- Pistola, F.; Sacco, S.; Degan, D.; Tiseo, C.; Ornello, R.; Carolei, A. Hypertension and stroke: Epidemiological aspects and clinical evaluation. High Blood Press. Cardiovasc. Prev. 2016, 23, 9–18. [Google Scholar] [CrossRef]
- Rubattu, S.; Volpe, M.; Kreutz, R.; Ganten, U.; Ganten, D.; Lindpaintner, K. Chromosomal mapping of genetic loci contributing to stroke in an animal model of a complex human disease. Nat. Genet. 1996, 13, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Rubattu, S.; Lee, M.A.; De Paolis, P.; Giliberti, R.; Gigante, B.; Lombardi, A.; Volpe, M.; Lindpaintner, K. Altered structure, regulation and function of the gene encoding atrial natriuretic peptide in the stroke-prone spontaneously hypertensive rat. Circ. Res. 1999, 85, 900–905. [Google Scholar] [CrossRef] [PubMed]
- Rubattu, S.; Giliberti, R.; De Paolis, P.; Stanzione, R.; Spinsanti, P.; Volpe, M. Effect of a regulatory mutation on the rat atrial natriuretic peptide gene transcription. Peptides 2002, 23, 555–560. [Google Scholar] [CrossRef]
- Rubattu, S.; Giliberti, R.; Ganten, U.; Volpe, M. A differential brain ANP expression cosegregates with occurrence of early stroke in the stroke-prone phenotype of the spontaneously hypertensive rat. J. Hypertens. 1999, 17, 1849–1852. [Google Scholar] [CrossRef] [PubMed]
- Jeffs, B.; Clark, J.S.; Anderson, N.H.; Gratton, J.; Brosnan, M.J.; Gauguier, D.; Reid, J.L.; Macrae, I.M.; Dominiczak, A.F. Sensitivity to cerebral ischaemic insult in a rat model of stroke is determined by a single genetic locus. Nat. Genet. 1997, 16, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, M.J.; Clark, J.S.; Jeffs, B.; Negrin, C.D.; Van Vooren, P.; Arribas, S.M.; Carswell, H.; Aitman, T.J.; Szpirer, C.; Macrae, I.M.; et al. Genes encoding atrial and brain natriuretic peptides as candidates for sensitivity to brain ischemia in stroke-prone hypertensive rats. Hypertension 1999, 33, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Morales, M.A.; Castello-Rruiz, M.; Burguete, M.C.; Jover-Mengual, T.; Aliena-Valero, A.; Centeno, J.M.; Alborch, E.; Salom, J.B.; Torregrosa, G.; Miranda, F.J. Molecular mechanisms underlying the neuroprotective role of atrial natriuretic peptide in experimental acute ischemic stroke. Mol. Cell. Endocrinol. 2018, 472, 1–9. [Google Scholar] [CrossRef]
- Rubattu, S.; Cotugno, M.; Forte, M.; Stanzione, R.; Bianchi, F.; Madonna, M.; Marchitti, S.; Volpe, M. Effects of dual Angiotensin type 1 receptor/neprilysin inhibition versus Angiotensin type 1 receptor inhibition on target organ injury in the stroke-prone spontaneously hypertensive rat. J. Hypertens. 2018, 36, 1902–1914. [Google Scholar]
- Rubattu, S.; Stanzione, R.; Di Angelantonio, E.; Zanda, B.; Evangelista, A.; Tarasi, D.; Gigante, B.; Pirisi, A.; Brunetti, E.; Volpe, M. Atrial natriuretic peptide gene polymorphisms and the risk of ischemic stroke in humans. Stroke 2004, 35, 814–818. [Google Scholar] [CrossRef]
- Cannone, V.; Huntley, B.K.; Olson, T.M.; Heublein, D.M.; Scott, C.G.; Bailey, K.R.; Redfield, M.M.; Rodeheffer, R.J.; Burnett, J.C., Jr. Atrial natriuretic peptide genetic variant rs5065 and riskfor cardiovascular disease in the general community: A 9-year follow-up study. Hypertension 2013, 62, 860–865. [Google Scholar] [CrossRef]
- Scarpino, S.; Marchitti, S.; Stanzione, R.; Evangelista, A.; Di Castro, S.; Savoia, C.; Quarta, G.; Sciarretta, S.; Ruco, L.; Volpe, M.; et al. ROS-mediated differential effects of the human atrial natriuretic peptide T2238C genetic variant on endothelial cells in vitro. J. Hypertens. 2009, 27, 1804–1813. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, S.; Marchitti, S.; Bianchi, F.; Moyes, A.; Barbato, E.; Di Castro, S.; Stanzione, R.; Cotugno, M.; Castello, L.; Calvieri, C.; et al. The C2238 Atrial Natriuretic Peptide Molecular Variant is Associated with Endothelial Damage and Dysfunction Through Natriuretic Peptide Receptor C Signaling. Circ. Res. 2013, 112, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
- Rubattu, S.; Marchitti, S.; Bianchi, F.; Di Castro, S.; Stanzione, R.; Cotugno, M.; Bozzao, C.; Sciarretta, S.; Volpe, M. The C2238/αANP variant is a negative modulator of both viability and function of coronary artery smooth muscle cells. PLoS ONE 2014, 9, e113108. [Google Scholar] [CrossRef] [PubMed]
- Stanzione, R.; Sciarretta, S.; Marchitti, S.; Bianchi, F.; Di Castro, S.; Scarpino, S.; Cotugno, M.; Frati, G.; Volpe, M.; Rubattu, S. C2238/αANP modulates Apolipoprotein E through Egr-1/miR199a in vascular smooth muscle cells in vitro. Cell Death Dis. 2015, 6, e2033. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, R.; Pignatelli, P.; Frati, G.; Nocella, C.; Stanzione, R.; Pastori, D.; Marchitti, S.; Valenti, V.; Santulli, M.; Barbato, E.; et al. C2238 ANP gene variant promotes increased platelet aggregation through the activation of Nox2 and the reduction of cAMP. Sci. Rep. 2017, 7, 3797. [Google Scholar] [CrossRef] [PubMed]
- Strisciuglio, T.; Barbato, E.; De Biase, C.; Di Gioia, G.; Cotugno, M.; Stanzione, R.; Trimarco, B.; Sciarretta, S.; Volpe, M.; Wijns, W.; et al. T2238C atrial natriuretic peptide gene variant and the response to antiplatelet therapy in stable ischemic heart disease patients. J. Cardiovasc. Transl. Res. 2018, 11, 36–41. [Google Scholar] [CrossRef]
- Rubattu, S.; Sciarretta, S.; Marchitti, S.; Bianchi, F.; Forte, M.; Volpe, M. The T2238C atrial natriuretic peptide molecular variant and the risk of cardiovascular diseases. Int. J. Mol. Sci. 2018, 19, 540. [Google Scholar] [CrossRef]
- Lynch, A.I.; Boerwinkle, E.; Davis, B.R.; Ford, C.E.; Eckfeldt, J.H.; Leiendecker-Foster, C.; Arnett, D.K. Pharmacogenetic association of the NPPA T2238C genetic variant with cardiovascular disease outcomes in patients with hypertension. JAMA 2008, 299, 296–307. [Google Scholar] [CrossRef]
- Calvieri, C.; Rubattu, S.; Volpe, M. Molecular mechanisms underlying cardiac antihypertrophic and antifibrotic effects of natriuretic peptides. J. Mol. Med. 2012, 90, 5–13. [Google Scholar] [CrossRef]
- Molkentin, J.D. A friend within the heart: Natriuretic peptide receptor signaling. J. Clin. Investig. 2003, 111, 1275–1277. [Google Scholar] [CrossRef]
- Rubattu, S.; Bigatti, G.; Evangelista, A.; Lanzani, C.; Stanzione, R.; Zagato, L.; Manunta, P.; Marchitti, S.; Venturelli, V.; Bianchi, G.; et al. Association of atrial natriuretic and type-A natriuretic peptide receptor gene polymorphisms with left ventricular mass in human essential hypertension. J. Am. Coll. Cardiol. 2006, 48, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Rubattu, S.; Sciarretta, S.; Ciavarella, G.M.; Venturelli, V.; De Paolis, P.; Tocci, G.; De Biase, L.; Ferrucci, A.; Volpe, M. Reduced levels of pro-atrial natriuretic peptide in hypertensive patients with metabolic syndrome and their relationship with LVH. J. Hypertens. 2007, 25, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Wang, H.; Dong, N.; Zhang, C.; Xue, B.; Wu, Q. Localization of corin and atrial natriuretic peptide expression in human renal segments. Clin. Sci. 2016, 130, 1655–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshihara, F.; Tokudome, T.; Kishimoto, I.; Otani, K.; Kuwabara, A.; Horio, T.; Kawano, Y.; Kangawa, K. Aggravated renal tubular damage and interstitial fibrosis in mice lacking guanylyl cyclase A (GC-A), a receptor for atrial and B-type natriuretic peptides. Clin. Exp. Nephrol. 2015, 19, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Caniffi, C.; Bouchet, G.; Costa, M.A.; Elesgaray, R.; Arranza, C.; Tomat, A.L. Chronic treatment with atrial natriuretic peptide in spontaneously hypertensive rats: Beneficial renal effects and sex differences. PLoS ONE 2015, 10, e0120362. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, P.; Boesen, M.; Olsen, M.; Wachtell, K.; Groenning, B. N-terminal pro brain natriuretic peptide in arterial hypertension—A marker for left ventricular dimensions and prognosis. Eur. J. Heart Fail. 2004, 6, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Olsen, M.H.; Wachtell, K.; Nielsen, O.W.; Hall, C.; Wergeland, R.; Ibsen, H.; Kjeldsen, S.E.; Devereux, R.B.; Dahlöf, B.; Hildebrandt, P.R. N-terminal brain natriuretic peptide predicted cardiovascular events stronger than high-sensitivity C-reactive protein in hypertension: A LIFE substudy. J. Hypertens. 2006, 24, 1531–1539. [Google Scholar] [CrossRef]
- Ikram, M.A.; Seshadri, S.; Bis, J.C.; Fornage, M.; DeStefano, A.L.; Aulchenko, Y.S.; Debette, S.; Lumley, T.; Folsom, A.R.; van den Herik, E.G.; et al. Genomewide association studies of stroke. NEJM 2009, 360, 1718–1728. [Google Scholar] [CrossRef]
- Rubattu, S.; Giusti, B.; Lotta, L.A.; Peyvandi, F.; Cotugno, M.; Stanzione, R.; Marchitti, S.; Palombella, A.M.; Di Castro, S.; Rasura, M.; et al. Association of a single nucleotide polymorphism of the NPR3 gene promoter with early onset ischemic stroke in an Italian cohort. Eur. J. Intern. Med. 2013, 24, 80–82. [Google Scholar] [CrossRef]
- Volpe, M.; Tocci, G.; Battistoni, A.; Rubattu, S. Angiotensin II receptor blocker nephrilysin inhibitor (ARNI): New avenues in cardiovascular therapy. High Blood Press. Cardiovasc. Prev. 2015, 22, 241–246. [Google Scholar] [CrossRef]
- Von Llueder, T.G.; Atar, D.; Krum, H. Current role of neprilysin inhibitors in hypertension and heart failure. Pharmacol. Ther. 2014, 144, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Kario, K.; Sun, N.; Chiang, F.T.; Supasyndh, O.; Baek, S.H.; Inubushi-Molessa, A.; Zhang, Y.; Gotou, H.; Lefkowitz, M.; Zhang, J. Efficacy and safety of LCZ696, a first-in-class angiotensin receptor neprilysin inhibitor, in Asian patients with hypertension: A randomized, double-blind, placebo-controlled study. Hypertension 2014, 63, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Ruilope, L.M.; Dukat, A.; Bohm, M.; Lacourciere, Y.; Gong, J.; Lefkowitz, M.P. Blood-pressure reduction with LCZ696, a novel dual-acting inhibitor of the angiotensin II receptor and neprilysin: A randomized, double-blind, placebo-controlled, active comparator study. Lancet 2010, 375, 1255–1266. [Google Scholar] [CrossRef]
- Wang, T.D.; Tan, R.S.; Lee, H.Y.; Ihm, S.H.; Rhee, M.Y.; Tomlinson, B.; Pal, P.; Yang, F.; Hirschhorn, E.; Prescott, M.F.; et al. Effects of Sacubitril/Valsartan (LCZ696) on natriuresis, diuresis, blood pressures, and NT-proBNP in salt-sensitive hypertension. Hypertension 2017, 69, 32–41. [Google Scholar] [PubMed]
- Kusaka, H.; Sueta, D.; Koibuchi, N.; Hasegawa, Y.; Nakagawa, T.; Lin, B.; Ogawa, H.; Kim-Mitsuyama, S. LCZ696, Angiotensin II receptor-neprilysin inhibitor, ameliorates high-salt-induced hypertension and cardiovascular injury more than valsartan alone. Am. J. Hypertens. 2015, 28, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.; Cockcroft, J.R.; Kario, K.; Zappe, D.H.; Brunel, P.C.; Wang, Q.; Guo, W. Effects of sacubitril/valsartan versus olmesartan on central hemodynamics in the erderly with systolic hypertension: The PARAMETER study. Hypertension 2017, 69, 411–420. [Google Scholar] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubattu, S.; Forte, M.; Marchitti, S.; Volpe, M. Molecular Implications of Natriuretic Peptides in the Protection from Hypertension and Target Organ Damage Development. Int. J. Mol. Sci. 2019, 20, 798. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040798
Rubattu S, Forte M, Marchitti S, Volpe M. Molecular Implications of Natriuretic Peptides in the Protection from Hypertension and Target Organ Damage Development. International Journal of Molecular Sciences. 2019; 20(4):798. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040798
Chicago/Turabian StyleRubattu, Speranza, Maurizio Forte, Simona Marchitti, and Massimo Volpe. 2019. "Molecular Implications of Natriuretic Peptides in the Protection from Hypertension and Target Organ Damage Development" International Journal of Molecular Sciences 20, no. 4: 798. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040798