Unravelling the Complexity of Plant Defense Induced by a Simultaneous and Sequential Mite and Aphid Infestation


Abstract
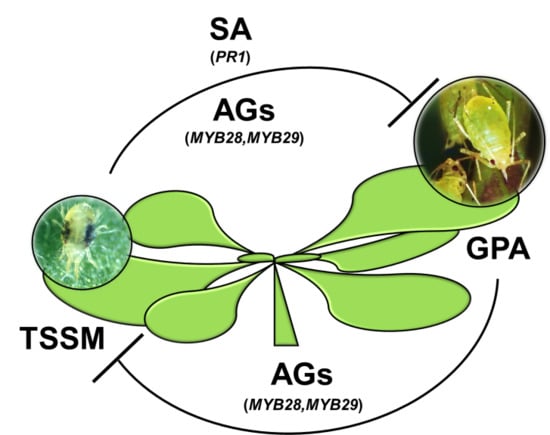
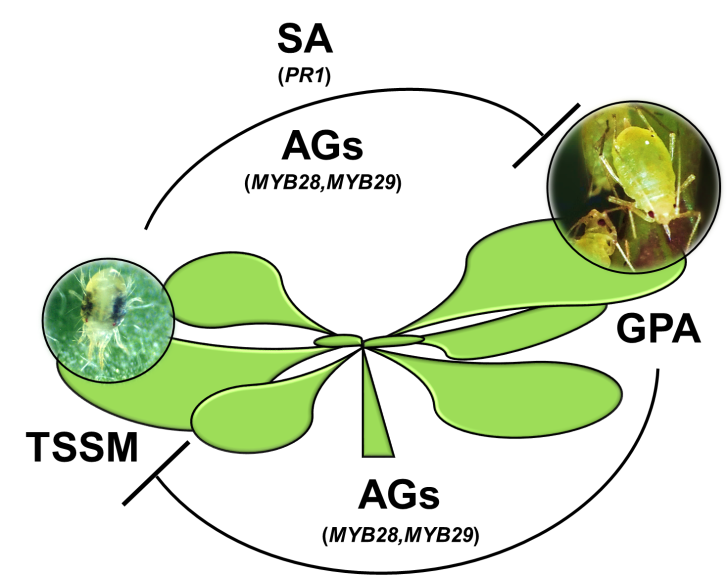
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
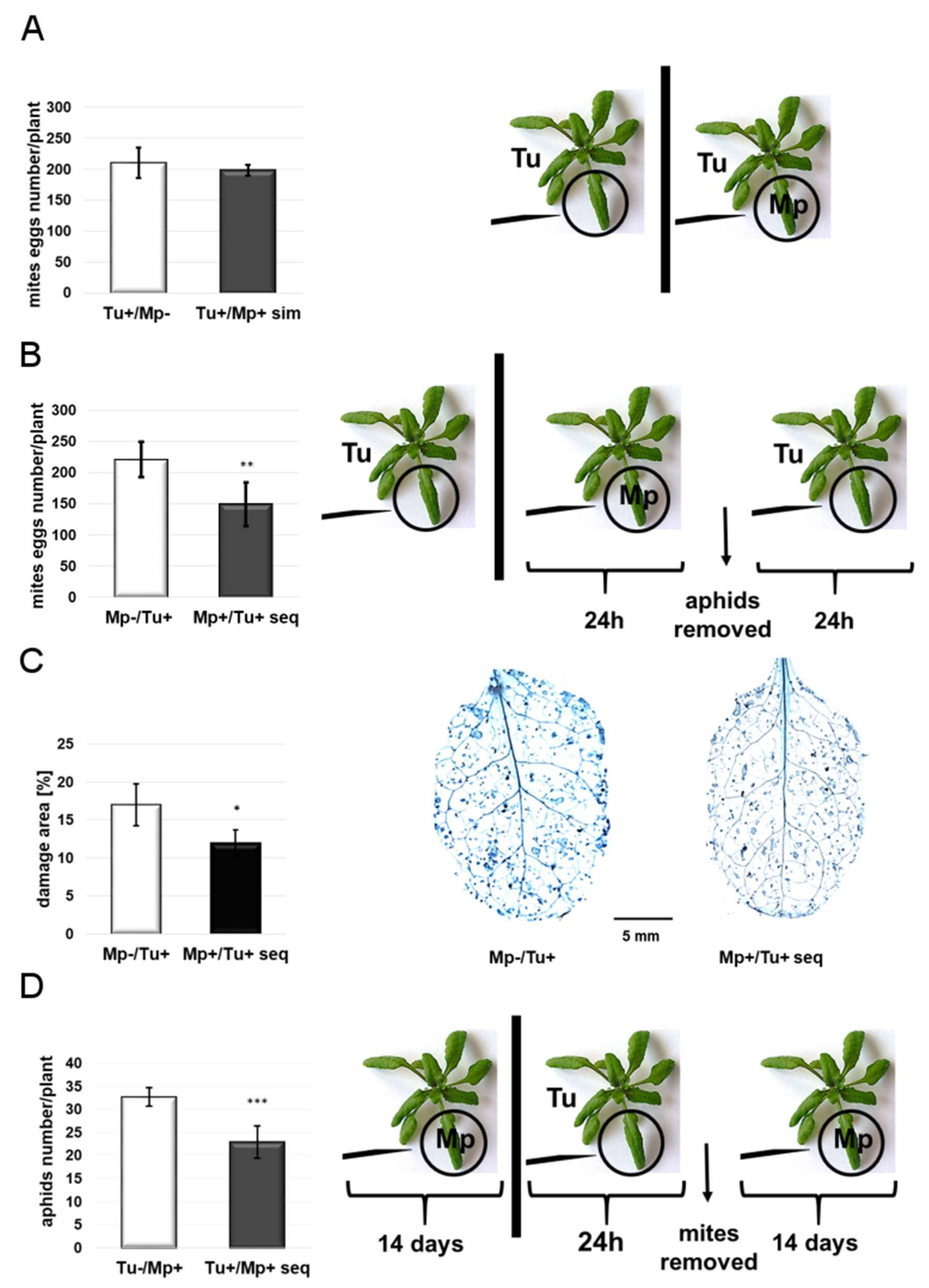
2.1. TSSM and GPA Reproductive Performance
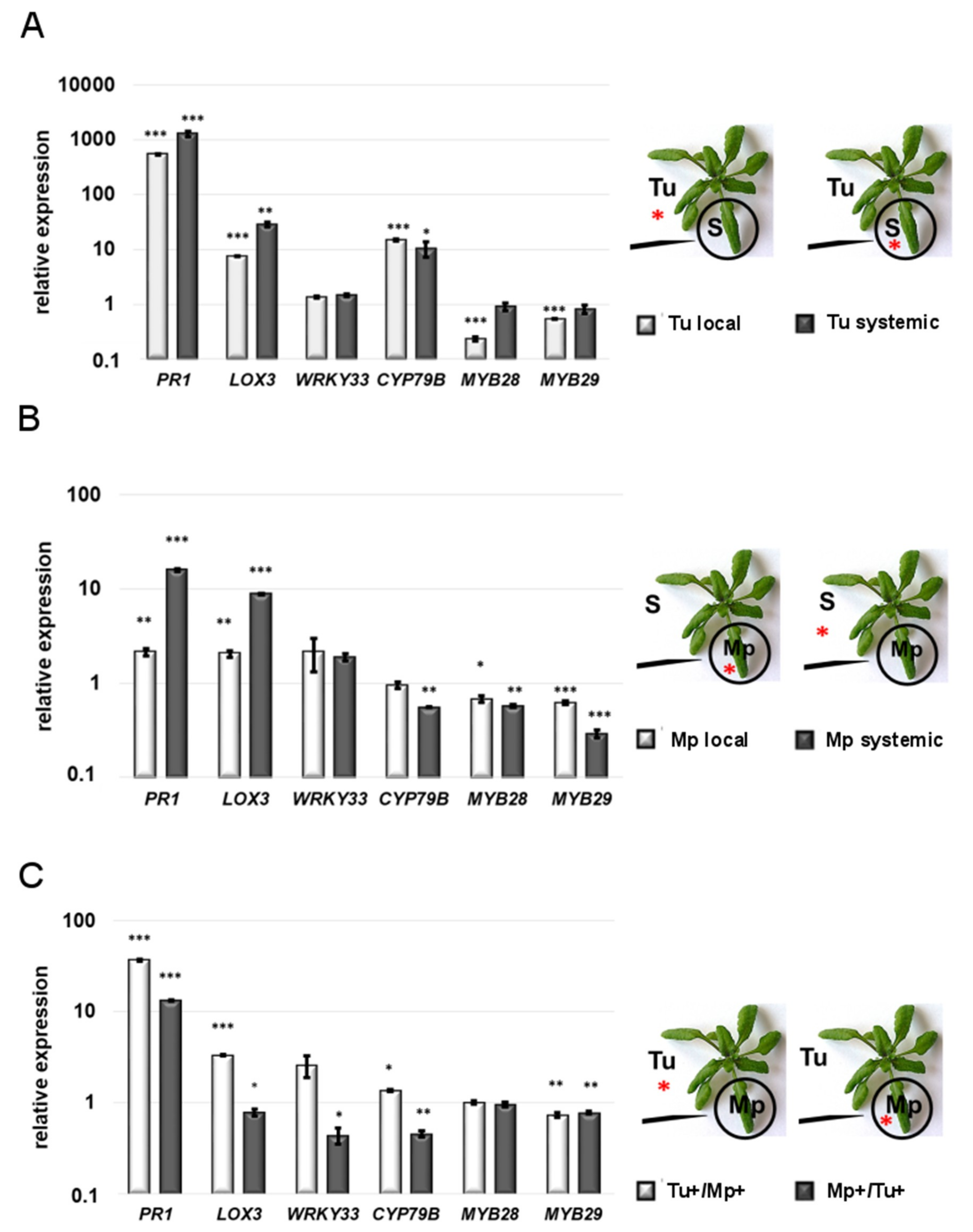
2.2. Gene Expression Analysis
2.2.1. Local Responses to Single TSSM and GPA Infestation
2.2.2. Systemic Responses to Single TSSM and GPA Infestation
2.2.3. Systemic Responses to Double Infestation
3. Discussion
4. Materials and Methods
4.1. Mite (TSSM) and Aphid (GPA) Population Rearing
4.2. Plant Material and Experimental Design
4.3. TSSM and GPA Single Infestation
4.4. Double Infestation Experiments
4.4.1. Simultaneous Infestation
4.4.2. Sequential Infestation
4.5. Leaf-Damage Assessment
4.6. RNA Isolation and cDNA Synthesis
4.7. Analysis of Gene Expression
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Adachi, M.; Yano, S. Ant-mediated indirect negative effects of aphids on spider mites living on the same plant. Exp. Appl. Acarol. 2017, 72, 15–21. [Google Scholar] [CrossRef]
- Ataide, L.M.S.; Pappas, M.L.; Schimmel, B.C.J.; Lopez-Orenes, A.; Alba, J.M.; Duarte, M.V.A.; Pallini, A.; Schuurink, R.C.; Kant, M.R. Induced plant-defenses suppress herbivore reproduction but also constrain predation of their offspring. Plant Sci. 2016, 252, 300–310. [Google Scholar] [CrossRef] [Green Version]
- Errard, A.; Ulrichs, C.; Kühne, S.; Mewis, I.; Drungowski, M.; Schreiner, M.; Baldermann, S. Single- versus Multiple-Pest Infestation Affects Differently the Biochemistry of Tomato (Solanum lycopersicum “Ailsa Craig”). J. Agric. Food Chem. 2015, 63, 10103–10111. [Google Scholar] [CrossRef] [PubMed]
- Hol, W.H.; De Boer, W.; Termorshuizen, A.J.; Meyer, K.M.; Schneider, J.H.; Van Der Putten, W.H.; Van Dam, N.M. Heterodera schachtii Nematodes Interfere with Aphid-Plant Relations on Brassica oleracea. J. Chem. Ecol. 2013, 39, 1193–203. [Google Scholar] [CrossRef] [PubMed]
- Hol, W.H.G.; Raaijmakers, C.E.; Mons, I.; Meyer, K.M.; van Dam, N.M. Root-Lesion Nematodes Suppress Cabbage Aphid Population Development by Reducing Aphid Daily Reproduction. Front. Plant Sci. 2016, 7, 111. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, S.D.; Novak, N.G.; Ju, C.J.-T.; Cooke, J.E.K. Potato, Solanum tuberosum, defense against Colorado potato beetle, Leptinotarsa decemlineata (Say): Microarray gene expression profiling of potato by Colorado potato beetle regurgitant treatment of wounded leaves. J. Chem. Ecol. 2008, 34, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Saona, C.R.; Musser, R.O.; Vogel, H.; Hum-Musser, S.M.; Thaler, J.S. Molecular, Biochemical, and Organismal Analyses of Tomato Plants Simultaneously Attacked by Herbivores from Two Feeding Guilds. J. Chem. Ecol. 2010, 36, 1043–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dam, N.M.; Witjes, L.; Svatoš, A. Interactions between aboveground and belowground induction of glucosinolates in two wild Brassica species. New Phytol. 2004, 161, 801–810. [Google Scholar] [CrossRef]
- Erb, M.; Meldau, S.; Howe, G.A. Role of phytohormones in insect-specific plant reactions. Trends Plant Sci. 2012, 17, 250–259. [Google Scholar] [CrossRef] [Green Version]
- Mithöfer, A.; Boland, W. Recognition of Herbivory-Associated Molecular Patterns. Plant Physiol. 2008, 146, 825–831. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, F.E.; Rivera-Vega, L.J.; Chung, S.H.; Ray, S.; Felton, G.W. Cues from chewing insects—The intersection of DAMPs, HAMPs, MAMPs and effectors. Curr. Opin. Plant Biol. 2015, 26, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Ton, J.; van der Ent, S.; van Hulten, M.H.A.; Pozo, M.; van Oosten, V.; van Loon, L.C.; Mauch-Mani, B.; Turlings, T.C.J.; Pieterse, C.M.J. Priming as a mechanism behind induced resistance against pathogens, insects and abiotic stress. IOBC/wprs Bull. 2009, 44, 3–13. [Google Scholar]
- Alba, J.M.; Schimmel, B.C.; Glas, J.J.; Ataide, L.M.; Pappas, M.L.; Villarroel, C.A.; Schuurink, R.C.; Sabelis, M.W.; Kant, M.R. Spider mites suppress tomato defenses downstream of jasmonate and salicylate independently of hormonal crosstalk. New Phytol. 2015, 205, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Glas, J.J.; Alba, J.M.; Simoni, S.; Villarroel, C.A.; Stoops, M.; Schimmel, B.C.; Schuurink, R.C.; Sabelis, M.W.; Kant, M.R. Defense suppression benefits herbivores that have a monopoly on their feeding site but can backfire within natural communities. BMC Biol. 2014, 12, 98. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.R.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.C.; Villarroel, C.A.; Ataide, L.M.; Dermauw, W.; Glas, J.J.; et al. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, C.M.; Clement, S.L. Molecular bases of plant resistance to arthropods. Annu. Rev. Entomol. 2012, 57, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Kerchev, P.I.; Karpińska, B.; Morris, J.A.; Hussain, A.; Verrall, S.R.; Hedley, P.E.; Fenton, B.; Foyer, C.H.; Hancock, R.D. Vitamin C and the Abscisic Acid-Insensitive 4 Transcription Factor Are Important Determinants of Aphid Resistance in Arabidopsis. Antioxid. Redox Signal. 2013, 18, 2091–2105. [Google Scholar] [CrossRef]
- Bensoussan, N.; Santamaria, M.E.; Zhurov, V.; Diaz, I.; Grbić, M.; Grbić, V. Plant-Herbivore Interaction: Dissection of the Cellular Pattern of Tetranychus urticae Feeding on the Host Plant. Front. Plant Sci. 2016, 7, 1105. [Google Scholar] [CrossRef]
- Miles, P.W. Aphid saliva. Biol. Rev. 1999, 74, 41–85. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Salivary secretions by aphids interacting with proteins of phloem wound responses. J. Exp. Bot. 2006, 57, 739–745. [Google Scholar] [CrossRef] [Green Version]
- Dermauw, W.; Wybouw, N.; Rombauts, S.; Menten, B.; Vontas, J.; Grbic, M.; Clark, R.M.; Feyereisen, R.; Van Leeuwen, T. A link between host plant adaptation and pesticide resistance in the polyphagous spider mite Tetranychus urticae. Proc. Natl. Acad. Sci. USA 2013, 110, e113–e122. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Dermauw, W. The Molecular Evolution of Xenobiotic Metabolism and Resistance in Chelicerate Mites. Annu. Rev. Entomol. 2016, 61, 475–498. [Google Scholar] [CrossRef] [PubMed]
- Jonckheere, W.; Dermauw, W.; Zhurov, V.; Wybouw, N.; Van den Bulcke, J.; Villarroel, C.A.; Greenhalgh, R.; Grbić, M.; Schuurink, R.C.; Tirry, L.; et al. The salivary protein repertoire of the polyphagous spider mite Tetranychus urticae: A quest for effectors. Mol. Cell. Proteom. 2016, 15, 3594–3613. [Google Scholar] [CrossRef] [PubMed]
- Santamaría, M.E.; González-Cabrera, J.; Martínez, M.; Grbic, V.; Castañera, P.; Díaz, L.; Ortego, F. Digestive proteases in bodies, and faeces of the two-spotted spider mite, Tetranychus urticae. J. Insect Physiol. 2015, 78, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Agut, B.; Gamir, J.; Jaques, J.A.; Flors, V. Systemic resistance in citrus to Tetranychus urticae induced by conspecifics is transmitted by grafting and mediated by mobile amino acids. J. Exp. Bot. 2016, 67, 5711–5723. [Google Scholar] [CrossRef] [PubMed]
- Ament, K.; Kant, M.R.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Jasmonic acid is a key regulator of spider mite-induced volatile terpenoid and methyl salicylate emission in tomato. Plant Physiol. 2004, 135, 2025–2037. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.; Ozawa, R.; Horiuchi, J.; Nishioka, T.; Takabayashi, J. Plant–plant interactions mediated by volatiles emitted from plants infested by spider mites. Biochem. Syst. Ecol. 2001, 29, 1049–1061. [Google Scholar] [CrossRef]
- Barczak-Brzyżek, A.; Kiełkiewicz, M.; Górecka, M.; Kot, K.; Karpińska, B.; Filipecki, M. Abscisic Acid Insensitive 4 transcription factor is an important player in the response of Arabidopsis thaliana to two-spotted spider mite (Tetranychus urticae) feeding. Exp. Appl. Acarol. 2017, 73, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Kant, M.R.; Ament, K.; Sabelis, M.W.; Haring, M.A.; Schuurink, R.C. Differential Timing of Spider Mite-Induced Direct and Indirect Defenses in Tomato Plants. Plant Physiol. 2004, 135, 483–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Williams, M.M.; Loh, Y.-T.; Lee, G.I.; Howe, G.A. Resistance of Cultivated Tomato to Cell Content-Feeding Herbivores Is Regulated by the Octadecanoid-Signaling Pathway. Plant Physiol. 2002, 130, 494–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarroel, C.A.; Jonckheere, W.; Alba, J.M.; Glas, J.J.; Dermauw, W.; Haring, M.A.; Van Leeuwen, T.; Schuurink, R.C.; Kant, M.R. Salivary proteins of spider mites suppress defenses in Nicotiana benthamiana and promote mite reproduction. Plant J. Cell Mol. Biol. 2016, 86, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Zhurov, V.; Navarro, M.; Bruinsma, K.A.; Arbona, V.; Santamaria, M.E.; Cazaux, M.; Wybouw, N.; Osborne, E.J.; Ens, C.; Rioja, C.; et al. Reciprocal responses in the interaction between Arabidopsis and the cell-content-feeding chelicerate herbivore spider mite. Plant Physiol. 2014, 164, 384–399. [Google Scholar] [CrossRef] [PubMed]
- De Vos, M.; Van Oosten, V.R.; Van Poecke, R.M.; Van Pelt, J.A.; Pozo, M.J.; Mueller, M.J.; Buchala, A.J.; Métraux, J.P.; Van Loon, L.C.; Dicke, M.; et al. Signal Signature and Transcriptome Changes of Arabidopsis During Pathogen and Insect Attack. Mol. Plant Microbe Interact. 2005, 18, 923–937. [Google Scholar] [CrossRef] [PubMed]
- Moran, P.J.; Thompson, G.A. Molecular Responses to Aphid Feeding in Arabidopsis in Relation to Plant Defense Pathways. Plant Physiol. 2001, 125, 1074–1085. [Google Scholar] [CrossRef] [PubMed]
- Barczak-Brzyżek, A.K.; Kiełkiewicz, M.; Gawroński, P.; Kot, K.; Filipecki, M.; Karpińska, B. Cross-talk between high light stress and plant defence to the two-spotted spider mite in Arabidopsis thaliana. Exp. Appl. Acarol. 2017, 73, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Jaouannet, M.; Rodriguez, P.A.; Thorpe, P.; Lenoir, C.J.G.; MacLeod, R.; Escudero-Martinez, C.; Bos, J.I.B. Plant immunity in plant–aphid interactions. Front. Plant Sci. 2014, 5, 663. [Google Scholar] [CrossRef]
- Birkenbihl, R.P.; Diezel, C.; Somssich, I.E. Arabidopsis WRKY33 is a key transcriptional regulator of hormonal and metabolic responses toward Botrytis cinerea infection. Plant Physiol. 2012, 159, 266–285. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Qamar, S.A.; Chen, Z.; Mengiste, T. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. Plant J. Cell Mol. Biol. 2006, 48, 592–605. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.-S.; Kim, J.S. Understanding of MYB Transcription Factors Involved in Glucosinolate Biosynthesis in Brassicaceae. Molecules 2017, 22, 1549. [Google Scholar]
- Rioja, C.; Zhurov, V.; Bruinsma, K.; Grbic, M.; Grbic, V. Plant-herbivore interactions: A case of an extreme generalist, the two-spotted spider mite, Tetranychus urticae. Mol Plant-Microbe Interact. 2017, 30, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A.; Halitschke, R.; Baldwin, I.T. Silencing the Jasmonate Cascade: Induced Plant Defenses and Insect Populations. Science 2004, 305, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Kammerhofer, N.; Egger, B.; Dobrev, P.; Vankova, R.; Hofmann, J.; Schausberger, P.; Wieczorek, K. Systemic above- and belowground cross talk: Hormone-based responses triggered by Heterodera schachtii and shoot herbivores in Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 7005–7017. [Google Scholar] [CrossRef] [PubMed]
- Breen, S.; Williams, S.J.; Outram, M.; Kobe, B.; Solomon, P.S. Emerging Insights into the Functions of Pathogenesis-Related Protein 1. Trends Plant Sci. 2017, 22, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, V.; Schneiter, R. Pathogen-Related Yeast (PRY) proteins and members of the CAP superfamily are secreted sterol-binding proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 16882–16887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-L.; Lee, C.-Y.; Cheng, K.-T.; Chang, W.-H.; Huang, R.-N.; Nam, H.G.; Chen, Y.R. Quantitative peptidomics study reveals that a wound-induced peptide from PR-1 regulates immune signaling in tomato. Plant Cell 2014, 26, 4135–4148. [Google Scholar] [CrossRef] [PubMed]
- Badenes-Perez, F.R.; Reichelt, M.; Gershenzon, J.; Heckel, D.G. Using plant chemistry and insect preference to study the potential of Barbarea (Brassicaceae) as a dead-end trap crop for diamondback moth (Lepidoptera: Plutellidae). Phytochemistry 2014, 98, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Wittstock, U.; Halkier, B.A. Glucosinolate research in the Arabidopsis era. Trends Plant Sci. 2002, 7, 263–270. [Google Scholar] [CrossRef]
- Gawrońska, H.; Kiełkiewicz, M. Effect of the carmine spider mite (Acarida: Tetranychidae) infestation and mechanical injury on the level of ABA in tomato plants. Acta Physiol. Plant. 1999, 21, 297–303. [Google Scholar] [CrossRef]
- Vos, I.A.; Verhage, A.; Schuurink, R.C.; Watt, L.G.; Pieterse, C.M.J.; Van Wees, S.C.M. Onset of herbivore-induced resistance in systemic tissue primed for jasmonate-dependent defenses is activated by abscisic acid. Front. Plant Sci. 2013, 4, 539. [Google Scholar] [CrossRef] [PubMed]
- Keogh, R.C.; Deverall, B.J.; McLeod, S. Comparison of histological and physiological responses to Phakopsora pachyrhizi in resistant and susceptible soybean. Trans. Br. Mycol. Soc. 1980, 74, 329–333. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiełkiewicz, M.; Barczak-Brzyżek, A.; Karpińska, B.; Filipecki, M. Unravelling the Complexity of Plant Defense Induced by a Simultaneous and Sequential Mite and Aphid Infestation. Int. J. Mol. Sci. 2019, 20, 806. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040806
Kiełkiewicz M, Barczak-Brzyżek A, Karpińska B, Filipecki M. Unravelling the Complexity of Plant Defense Induced by a Simultaneous and Sequential Mite and Aphid Infestation. International Journal of Molecular Sciences. 2019; 20(4):806. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040806
Chicago/Turabian StyleKiełkiewicz, Małgorzata, Anna Barczak-Brzyżek, Barbara Karpińska, and Marcin Filipecki. 2019. "Unravelling the Complexity of Plant Defense Induced by a Simultaneous and Sequential Mite and Aphid Infestation" International Journal of Molecular Sciences 20, no. 4: 806. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20040806