Effects of 2,4-Dichlorophenoxyacetic Acid on Cucumber Fruit Development and Metabolism


Abstract
:1. Introduction
2. Results
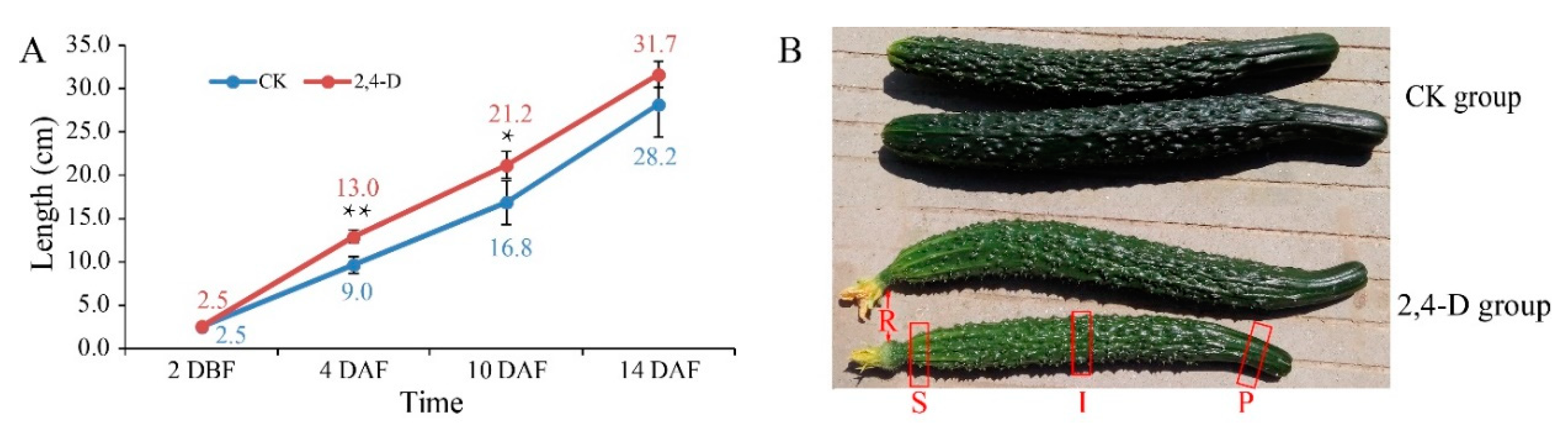
2.1. Effects of 2,4-D on Cucumber Fruit Development
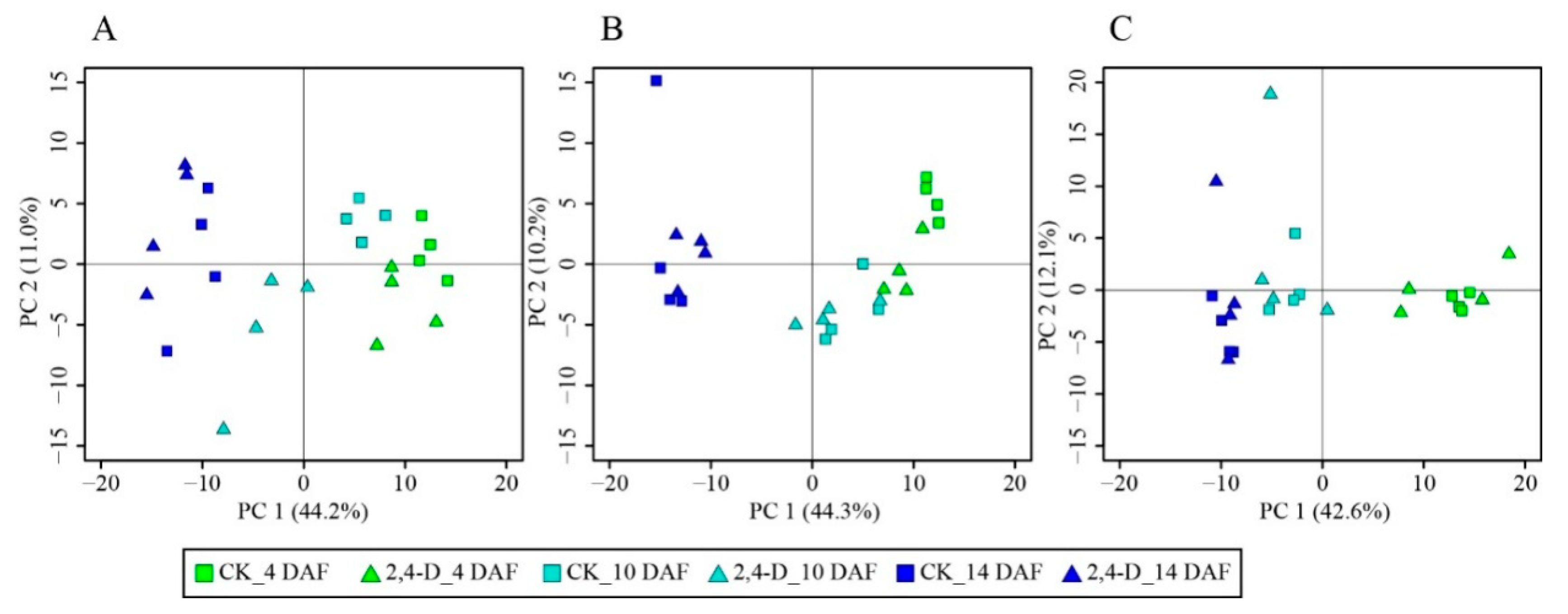
2.2. Kinetic Metabolic Change Patterns of Cucumber Fruit Were Similar between 2,4-D and CK Groups
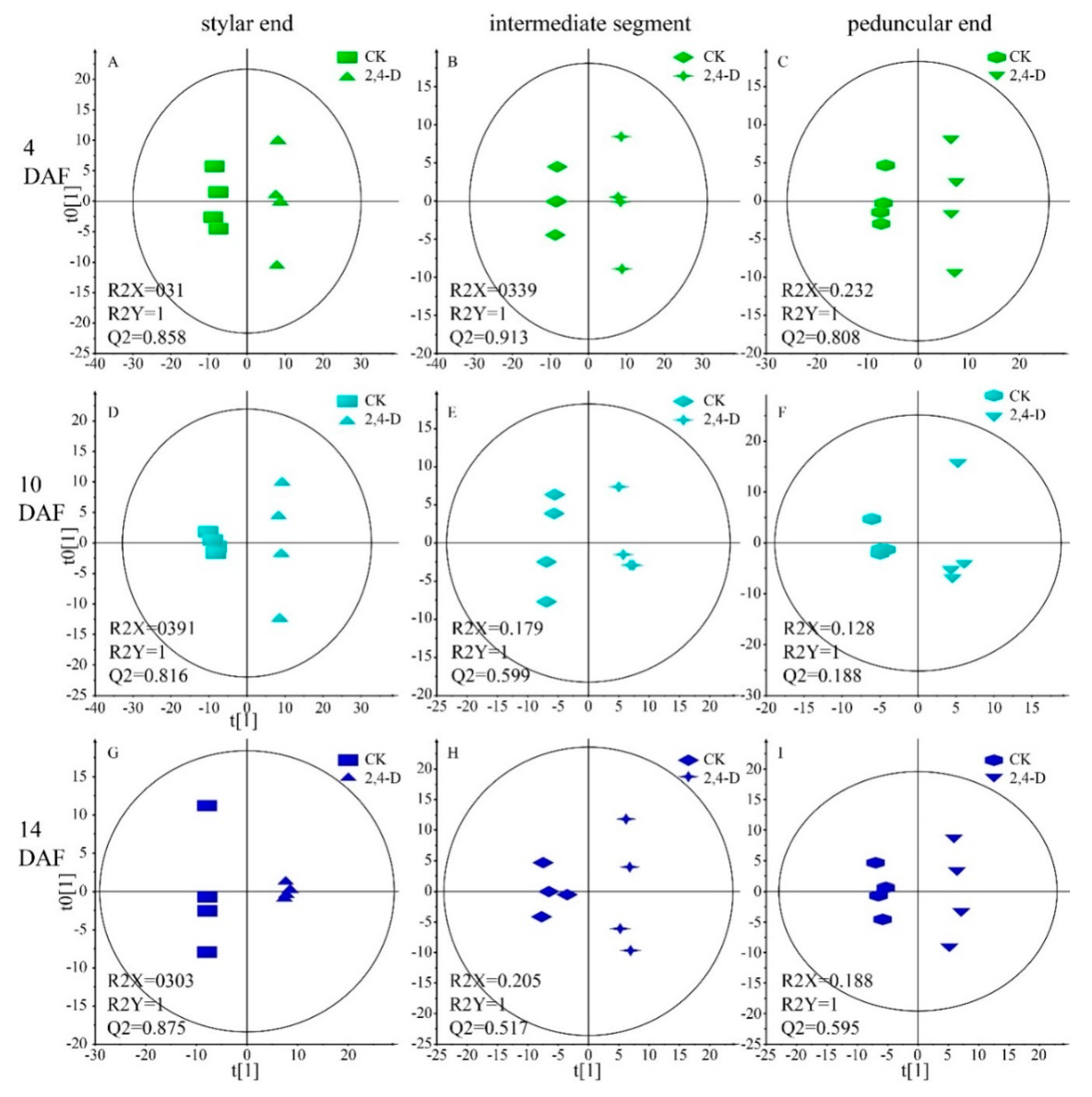
2.3. Metabolic Pathways Affected by 2,4-D at 4 DAF
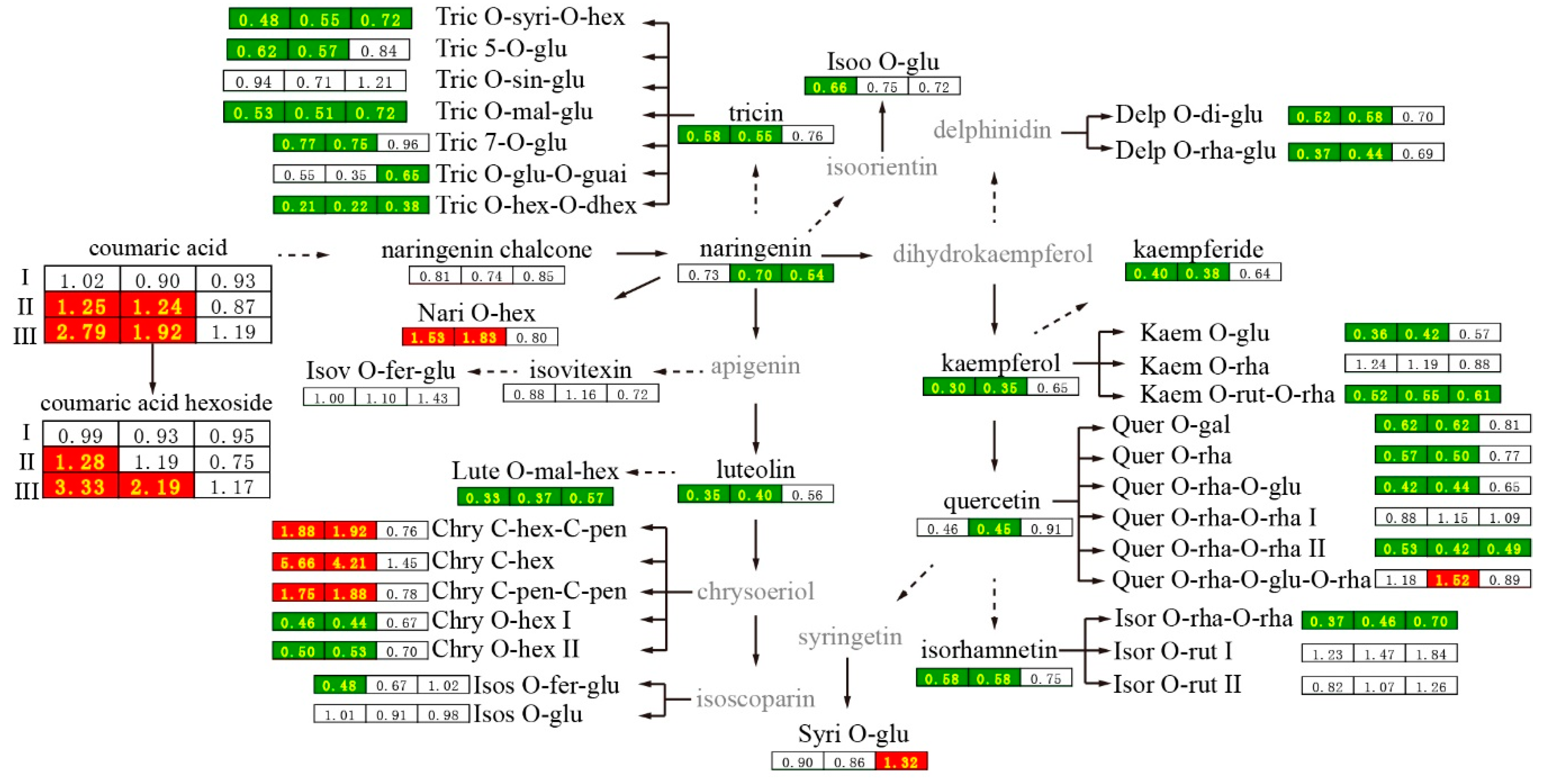
2.4. Metabolic Differences Generated by 2,4-D at 10 DAF
2.5. Metabolic Difference between the 2,4-D and CK Groups at the Harvest Stage
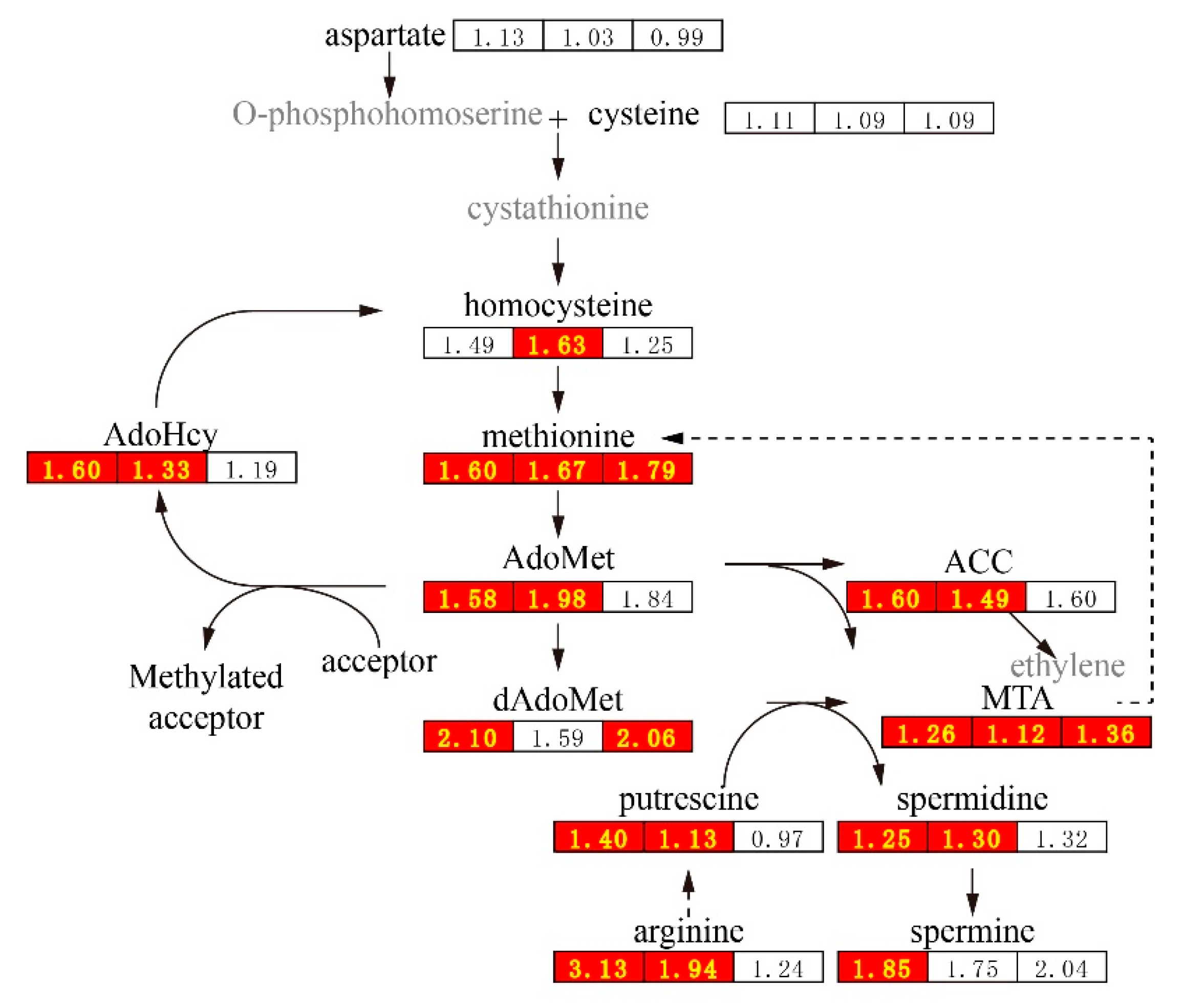
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Metabolite Profiling
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 2,4-D | 2,4-dichlorophenoxyacetic acid |
| ACC | 1-aminocyclopropane-1-carboxylate |
| AdoMet | S-adenosylmethionine |
| CID | Collision-induced dissociation |
| CK | Black controls |
| CPPU | N-(2-chloro-4-pyridyl)-N′-phenylurea |
| dAdoMet | S-adenosylmethioninamine |
| DAF | Days after flowering |
| DBF | Days before flowering |
| ESI | Electrospray ionization |
| HOTrE | hydroxyl-octadecatrienoate |
| LysoPC | lysophosphocholine |
| MTA | 5-methylthioadenosine |
| NADH | Reduced nicotinamide adenine dinucleotide |
| NAD+ | Nicotinamide adenine dinucleotide |
| NADP+ | Nicotinamide adenine dinucleotide phosphate |
| OPLS-DA | Orthogonal partial least squares projection to latent structures-discriminant analysis |
| PCA | Principle component analysis |
| PCD/PCDL | Personal compound database and library |
| PGR | Plant growth regulators |
| TCA | Tricarboxylic acid |
| UHPLC-qTOF-MS | Ultra-high performance liquid chromatography-quadrupole time of flight mass spectrometry |
| UV | Ultraviolet ray |
| VIP | Variable importance in projection |
References
- Elotmani, M.; Coggins, C.W., Jr.; Agustí, M.; Lovatt, C.J. Plant growth regulators in citriculture: World current uses. Crit. Rev. Plant Sci. 2000, 19, 395–447. [Google Scholar] [CrossRef]
- Prajapati, S.; Jamkar, T.; Singh, O.; Raypuriya, N.; Mandloi, R.; Jain, P. Plant growth regulators in vegetable production: An overview. Plant Arch. 2015, 15, 619–626. [Google Scholar]
- Ben-Arie, R.; Sarig, P.; Cohen-Ahdut, Y.; Zutkhi, Y.; Sonego, L.; Kapulonov, T.; Lisker, N. CPPU and GA3 effects on pre-and post-harvest quality of seedless and seeded grapes. Acta Hortic. 1997, 463, 349–358. [Google Scholar] [CrossRef]
- Han, D.; Lee, C. effects of GA3, CPPU and ABA applications on the quality of kyoho (Vitis vinifera L. x Labrusca L.) grape. Acta Hortic. 2004, 653, 193–197. [Google Scholar] [CrossRef]
- Qian, C.; Ren, N.; Wang, J.; Xu, Q.; Chen, X.; Qi, X.; Qian, C.; Ren, N.; Wang, J.; Xu, Q. Effects of exogenous application of CPPU, NAA and GA4+7 on parthenocarpy and fruit quality in cucumber (Cucumis sativus L.). Food Chem. 2017, 243, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y. Auxin biosynthesis and its role in plant development. Annu. Rev. Plant Biol. 2010, 61, 49–64. [Google Scholar] [CrossRef] [PubMed]
- Steffen, V.; Jirí, F. Auxin: A trigger for change in plant development. Cell 2009, 136, 1005–1016. [Google Scholar]
- Petrášek, J.; Simon, S. Why plants need more than one type of auxin. Plant Sci. 2011, 180, 454–460. [Google Scholar]
- Bhalerao, R.P.; Bennett, M.J. The case for morphogens in plants. Nat. Cell Biol. 2003, 5, 939–943. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Zhao, H.; Wang, W.; Xu, M.; Shi, J.; Nie, X.; Yang, G. Identification of conserved and diverse metabolic shift of the stylar, intermediate and peduncular segments of cucumber fruit during development. Int. J. Mol. Sci. 2018, 19, 135. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Tahir, F.M.; Rajwana, I.A.; Raza, S.A.; Asad, H.U. Comparative evaluation of plant growth regulators for preventing premature fruit drop and improving fruit quality parameters in ‘Dusehri’ Mango. Int. J. Fruit Sci. 2012, 12, 372–389. [Google Scholar] [CrossRef]
- Goren, R.; Huberman, M. Effects of ethylene and 2,4-D on the activity of cellulase isoenzymes in abscission zones of the developing orange fruit. Physiol. Plantarum 2010, 37, 123–130. [Google Scholar]
- Maroto, J.V.; Miguel, A.; Lopez-Galarza, S.; Bautista, A.S.; Pascual, B.; Alagarda, J.; Guardiola, J.L. Parthenocarpic fruit set in triploid watermelon induced by CPPUand 2,4-D applications. Plant Growth Regul. 2005, 45, 209–213. [Google Scholar] [CrossRef]
- Medeiros, Ê.C.D.; Siqueira, D.L.D.; Salomão, L.C.C.; Neves, J.C.L.; Pereira, W.E. Use of 2,4-D and GA3 to control ‘Hamlin’ orange fruit drop. Revista Ceres 2000, 47, 287–301. [Google Scholar]
- Vendrell, M. Dual effect of 2, 4-D on ethylene production and ripening of tomato fruit tissue. Physiol. Plantarum 2010, 64, 559–563. [Google Scholar] [CrossRef]
- Fu, Q.; Mo, J.; Xian, S.; Zhou, Y.; Ruan, L.; Li, W. Effects of 2,4-D on major appearance commodity traits and eating quality of cucumber. Changjiang Veg. 2018, 2, 60–62. [Google Scholar]
- Zhao, H.; Yang, G.; Liang, S.; Huang, Q.; Wang, Q.; Dai, W.; Zhang, Z.; Wang, W.; Song, W.; Cai, Z. The dissipation and risk assessment of 2,4-D sodium, a preharvest anti-fruit-drop plant hormone in bayberries. Environ. Sci. Pollut. Res. Int. 2017, 24, 24327–24332. [Google Scholar] [CrossRef] [PubMed]
- Umetrics, M. User Guide to SIMCA; MKS Umetrics AB: Malmö, Sweden, 2013. [Google Scholar]
- Ravanel, S.; Gakière, B.; Job, D.; Douce, R. The specific features of methionine biosynthesis and metabolism in plants. Proc. Natl. Acad. Sci. USA 1998, 95, 7805–7812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovanelli, J.; Mudd, S.H.; Datko, A.H. Quantitative analysis of pathways of methionine metabolism and their regulation in Lemna. Plant Physiol. 1985, 78, 555–560. [Google Scholar] [PubMed]
- Fuso, A.; Seminara, L.; Cavallaro, R.A.; D’Anselmi, F.; Scarpa, S. S-adenosylmethionine/homocysteine cycle alterations modify DNA methylation status with consequent deregulation of PS1 and BACE and beta-amyloid production. Mol. Cell. Neurosci. 2005, 28, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Chiang, P.K.; Gordon, R.K.; Tal, J.; Zeng, G.C.; Doctor, B.P.; Pardhasaradhi, K.; Mccann, P.P. S-Adenosylmethionine and methylation. FASEB J. 1996, 10, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Bürstenbinder, K.; Waduwara, I.; Schoor, S.; Moffatt, B.A.; Wirtz, M.; Minocha, S.C.; Oppermann, Y.; Bouchereau, A.; Hell, R.; Sauter, M. Inhibition of 5’-methylthioadenosine metabolism in the Yang cycle alters polyamine levels, and impairs seedling growth and reproduction in Arabidopsis. Plant J. 2010, 62, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Kusano, T.; Berberich, T.; Tateda, C.; Takahashi, Y. Polyamines: Essential factors for growth and survival. Planta 2008, 228, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Cronan, J., Jr.; Laporte, D. Tricarboxylic acid cycle and glyoxylate bypass. EcoSal Plus 2006, 1, 206–216. [Google Scholar]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, T.B.; Basset, G.J.; Borel, P.; Carrari, F.; Dellapenna, D.; Fraser, P.D.; Hellmann, H.; Osorio, S.; Rothan, C.; Valpuesta, V. Vitamin deficiencies in humans: Can plant science help? Plant Cell 2012, 24, 395–414. [Google Scholar] [CrossRef] [PubMed]
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New roles for old molecules. J. Integr. Plant Biol. 2010, 52, 98–111. [Google Scholar] [PubMed]
- Agati, G.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Pollastri, S.; Tattini, M. Functional roles of flavonoids in photoprotection: New evidence, lessons from the past. Plant Physiol. Biochem. 2013, 72, 35–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cronjé, P.J.R.; Crouch, E.M.; Huysamer, M. Postharvest calyx retention of citrus fruit. Acta Hortic. 2005, 682, 369–376. [Google Scholar] [CrossRef]
- Modise, D.M.; Likuku, A.S.; Thuma, M.; Phuti, R. The influence of exogenously applied 2,4-dichlorophenoxyacetic acid on fruit drop and quality of navel oranges (Citrus sinensis L.). Afr. J. Biotechnol. 2009, 8, 2131–2137. [Google Scholar]
- Kurnlawati, B.; Hamim. Physiological responses and fruit retention of carambola fruit (Averrhoa carambola L.) induced by 2,4-D and GA3. Hayati J. Biosci. 2009, 16, 9–14. [Google Scholar] [CrossRef]
- Ma, Q.; Ding, Y.; Chang, J.; Sun, X.; Zhang, L.; Wei, Q.; Cheng, Y.; Chen, L.; Xu, J.; Deng, X. Comprehensive insights on how 2,4-dichlorophenoxyacetic acid retards senescence in post-harvest citrus fruits using transcriptomic and proteomic approaches. J. Exp. Bot. 2014, 65, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Tittle, F.L.; Goudey, J.S.; Spencer, M.S. Effect of 2,4-dichlorophenoxyacetic acid on endogenous cyanide, beta-cyanoalanine synthase activity, and ethylene evolution in seedlings of soybean and barley. Plant Physiol. 1990, 94, 1143–1148. [Google Scholar] [CrossRef] [PubMed]
- Morgan, P.W.; Hall, W.C. Effect of 2,4-dichlorophenoxyacetic acid on the production of ethylene by cotton and grain sorghum. Physiol. Plantarum 2010, 15, 420–427. [Google Scholar] [CrossRef]
- Pinfield, N.J.; Tillberg, E. The regulation by 2,4-dichlorophenoxyacetic acid of endogenous abscisic Acid levels in seedlings of Cucurbita pepo L. J. Plant Physiol. 1987, 129, 211–217. [Google Scholar] [CrossRef]
- Henderson, J.H.; Deese, D.C. Correlation between endogenous auxin and its destruction in vivo by 2:4-dichlorophenoxyacetic acid in plants. Nature 1954, 174, 967. [Google Scholar] [CrossRef] [PubMed]
- Skelton, J.J.; Rong, M.; Riechers, D.E. Waterhemp (Amaranthus tuberculatus) control under drought stress with 2,4-dichlorophenoxyacetic acid and glyphosate. Weed Biol. Manag. 2016, 16, 34–41. [Google Scholar] [CrossRef]
- Mansfeld, B.N.; Colle, M.; Kang, Y.; Jones, A.D.; Grumet, R. Transcriptomic and metabolomic analyses of cucumber fruit peels reveal a developmental increase in terpenoid glycosides associated with age-related resistance to Phytophthora capsici. Hortic. Res. 2017, 4, 17022. [Google Scholar] [CrossRef] [PubMed]
- Klemš, M.; Truksa, M.; Macháccaron, I.; ková; Eder, J.; Procházka, S. Uptake, transport and metabolism of 14C-2,4-dichlorophenoxyacetic acid (14C-2,4-D) in cucumber (Cucumis sativus L.) explants. Plant Growth Regul. 1998, 26, 195–202. [Google Scholar]
- Slife, F.W.; Key, J.L.; Yamaguchi, S.; Crafts, A.S. Penetration, Translocation, and Metabolism of 2,4-D and 2,4,5-T in Wild and Cultivated Cucumber Plants. Weeds 1962, 10, 29–35. [Google Scholar] [CrossRef]
- Raghavan, C.; Ong, E.K.; Dalling, M.J.; Stevenson, T.W. Regulation of genes associated with auxin, ethylene and ABA pathways by 2, 4-dichlorophenoxyacetic acid in Arabidopsis. Funct. Integr. Genom. 2006, 6, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Pasquer, F.; Ochsner, U.; Zarn, J.; Keller, B. Common and distinct gene expression patterns induced by the herbicides 2,4-dichlorophenoxyacetic acid, cinidon-ethyl and tribenuron-methyl in wheat. Pest Manag. Sci. 2006, 62, 1155. [Google Scholar] [CrossRef] [PubMed]
- Karuppanapandian, T.; Wang, H.W.; Prabakaran, N.; Jeyalakshmi, K.; Kwon, M.; Manoharan, K.; Kim, W. 2,4-dichlorophenoxyacetic acid-induced leaf senescence in mung bean (Vigna radiata L. Wilczek) and senescence inhibition by co-treatment with silver nanoparticles. Plant Physiol. Biochem. 2011, 49, 168. [Google Scholar] [CrossRef] [PubMed]
- Pazmiño, D.M.; Rodríguez-Serrano, M.; Romero-Puertas, M.C.; Archilla-Ruiz, A.; Río, L.A.D.; Sandalio, L.M. Differential response of young and adult leaves to herbicide 2,4-dichlorophenoxyacetic acid in pea plants: Role of reactive oxygen species. Plant Cell Environ. 2011, 34, 1874–1889. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.W.; Marth, P.C. Effects of 2,4-dichlorophenoxyacetic acid on the ripening of detached fruit. Bot. Gazette 1944, 106, 199–207. [Google Scholar] [CrossRef]
- Barry, C.S.; Giovannoni, J.J. Ethylene and fruit ripening. Physiol. Plantarum 1997, 26, 143. [Google Scholar] [CrossRef]
- Wang, B.; Wang, J.; Hao, L.; Jianyong, Y.I.; Zhang, J.; Lin, L.; Wu, Y.U.; Feng, X.; Cao, J.; Jiang, W. Reduced chilling injury in mango fruit by 2,4-dichlorophenoxyacetic acid and the antioxidant response. Postharvest Biol. Technol. 2008, 48, 172–181. [Google Scholar] [CrossRef]
- Nassar, A.M.K.; Adss, I.A.A. 2,4-Dichlorophenoxy acetic acid, abscisic acid, and hydrogen peroxide induced resistance-related components against potato early blight (Alternaria solani, Sorauer). Ann. Agric. Sci. 2016, 61, 15–23. [Google Scholar] [CrossRef]
- Xu, E.; Brosché, M. Salicylic acid signaling inhibits apoplastic reactive oxygen species signaling. BMC Plant Biol. 2014, 14, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, W.Y.; Yang, H.C.; Shao, H.B.; Zheng, A.Z.; Brestic, M. The alleviative effects of salicylic acid on the activities of catalase and superoxide dismutase in malting barley (Hordeum uhulgare L.) seedling leaves stressed by heavy metals. Acta Hydrochim. Hydrobiol. 2014, 42, 88–97. [Google Scholar]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Ionkova, I. Optimization of flavonoid production in cell cultures of Astragalus missouriensis Nutt. (Fabaceae). Pharmacogn. Mag. 2009, 4, 92–97. [Google Scholar]
- Ozeki, Y.; Komamine, A. Changes in activities of enzymes involved in general phenylpropanoid metabolism during the induction and reduction of anthocyanin synthesis in a carrot suspension Culture as regulated by 2,4-D. Physiol. Plantarum 2010, 69, 123–128. [Google Scholar] [CrossRef]
- Ban, T.; Ishimaru, M.; Kobayashi, S.; Shiozaki, S.; Goto-Yamamoto, N.; Horiuchi, S. Abscisic acid and 2,4-dichlorophenoxyacetic acid affect the expression of anthocyanin biosynthetic pathway genes in ‘Kyoho’ grape berries. J. Pomol. Hortic. Sci. 2003, 78, 586–589. [Google Scholar] [CrossRef]
- Schuler, M.A.; Werckreichhart, D. Functional genomics of P450s. Annu. Rev. Plant Biol. 2003, 54, 629–667. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Tohge, T.; Chan, S.A.; Song, Y.; Rao, J.; Cui, B.; Lin, H.; Wang, L.; Fernie, A.R.; Zhang, D. Identification of conserved and diverse metabolic shifts during rice grain development. Sci. Rep. 2016, 6, 20942. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN: A metabolite mass spectral database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Class a | Ratio b | p-Value c | VIP d |
|---|---|---|---|---|
| salicylic acid | Ben | 0.28 | 4.25 × 10−3 | 1.44 |
| salicylic acid O-glucoside | Ben | 0.25 | 3.50 × 10−3 | 1.39 |
| citraconate | Car | 0.65 | 3.76 × 10−4 | 1.54 |
| aconitic acid | Car | 0.41 | 3.56 × 10−3 | 1.41 |
| ascorbate | Cof | 0.04 | 6.72 × 10−4 | 1.54 |
| naringenin | Fla | 0.08 | 9.07 × 10−4 | 1.58 |
| isoorientin 2′-O-glucoside | Fla | 0.11 | 1.10 × 10−3 | 1.56 |
| quercetin-3-O-rhamnosyl(1-2)-glucoside-7-O-rhamnoside | Fla | 0.08 | 2.01 × 10−3 | 1.56 |
| kaempferol-3-O-rhamnoside | Fla | 0.02 | 2.55 × 10−4 | 1.54 |
| tricin-O-hexoside-deoxyhexoside | Fla | 0.04 | 1.17 × 10−6 | 1.51 |
| isovitexin | Fla | 0.03 | 2.03 × 10−4 | 1.49 |
| Naringenin O-hexoside | Fla | 0.03 | 1.43 × 10−3 | 1.48 |
| kaempferol-3-O-glucoside | Fla | 0.27 | 0.014 | 1.46 |
| quercetin-3-O-rhamnoside-7-O-glucosid | Fla | 0.28 | 0.013 | 1.45 |
| tricin 7-O-(6′-O-malonyl)-β-d-glucopyranoside | Fla | 0.31 | 0.015 | 1.44 |
| astilbin | Fla | 0.02 | 8.47 × 10−4 | 1.44 |
| luteolin | Fla | 0.27 | 0.017 | 1.44 |
| chrysoeriol C-pentoside-C-pentoside | Fla | 0.11 | 2.40 × 10−3 | 1.41 |
| kaempferol | Fla | 0.29 | 2.10 × 10−3 | 1.41 |
| luteolin-O-malonylhexoside | Fla | 0.29 | 8.71 × 10−3 | 1.41 |
| chrysoeriol-C-hexoside-C-pentoside | Fla | 0.1 | 2.73 × 10−3 | 1.4 |
| caffeic acid hexoside II | Hyd | 0.02 | 7.54 × 10−5 | 1.59 |
| ferulic acid hexoside II | Hyd | 0.08 | 3.75 × 10−5 | 1.53 |
| coumaric acid II | Hyd | 0.11 | 6.40 × 10−4 | 1.51 |
| coumaric acid hexoside II | Hyd | 0.07 | 7.32 × 10−4 | 1.49 |
| feruloyl quinic acid I | Hyd | 0.49 | 6.77 × 10−3 | 1.47 |
| feruloyl quinic acid II | Hyd | 0.28 | 8.36 × 10−3 | 1.46 |
| coumaroyl quinic acid | Hyd | 0.26 | 5.02 × 10−3 | 1.44 |
| 2-hydroxyadipate | Lip | 0.56 | 0.013 | 1.43 |
| terpenyl-pentosyl-glucoside I | Mon | 0.06 | 1.43 × 10−3 | 1.41 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, C.; Zhao, H.; Shi, J.; Li, J.; Nie, X.; Yang, G. Effects of 2,4-Dichlorophenoxyacetic Acid on Cucumber Fruit Development and Metabolism. Int. J. Mol. Sci. 2019, 20, 1126. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051126
Hu C, Zhao H, Shi J, Li J, Nie X, Yang G. Effects of 2,4-Dichlorophenoxyacetic Acid on Cucumber Fruit Development and Metabolism. International Journal of Molecular Sciences. 2019; 20(5):1126. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051126
Chicago/Turabian StyleHu, Chaoyang, Huiyu Zhao, Jianxin Shi, Jian Li, Xiangbo Nie, and Guiling Yang. 2019. "Effects of 2,4-Dichlorophenoxyacetic Acid on Cucumber Fruit Development and Metabolism" International Journal of Molecular Sciences 20, no. 5: 1126. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20051126