Instability of Alien Chromosome Introgressions in Wheat Associated with Improper Positioning in the Nucleus

 , , ,
, , ,  and
and
Abstract
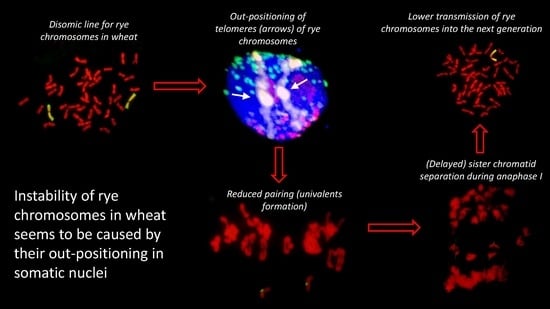
:
1. Introduction
2. Results
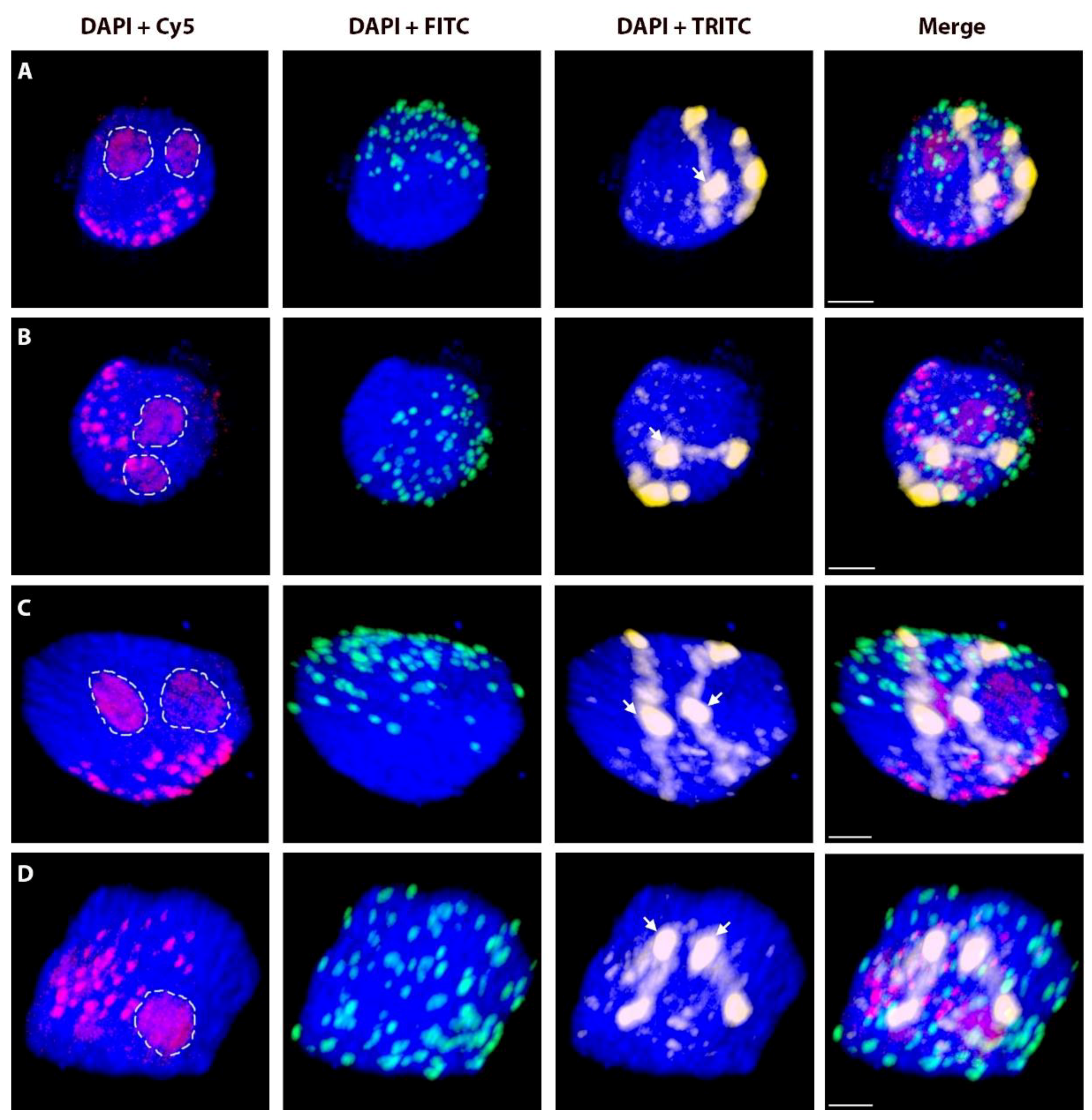
2.1. Morphometrical Characteristics of G1 Interphase Nuclei of Wheat-Rye Introgression Lines
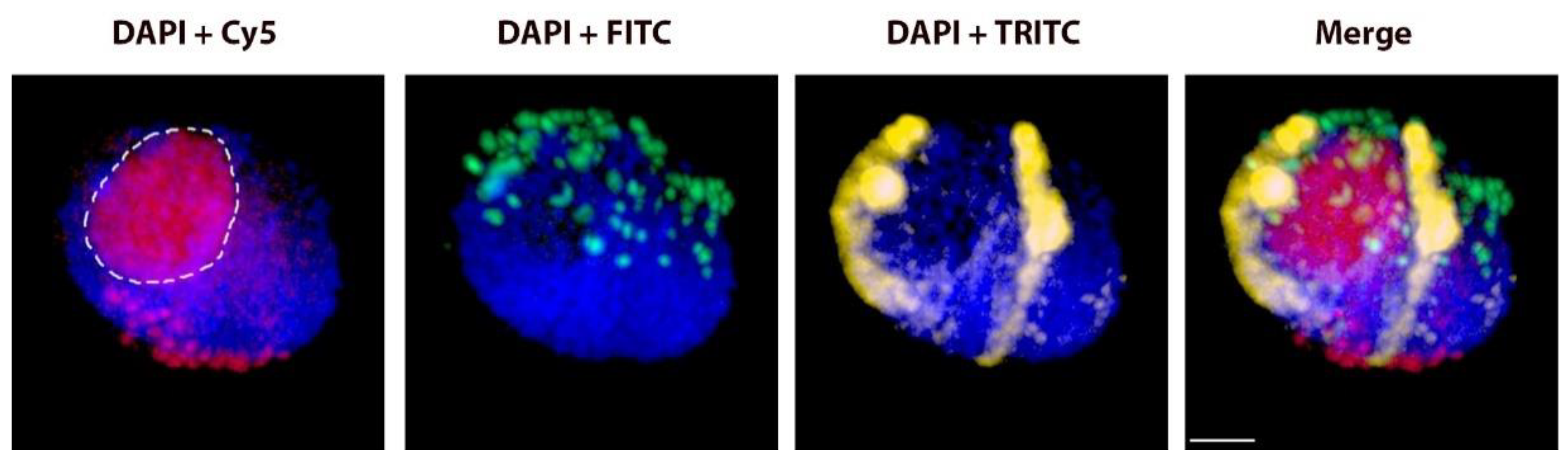
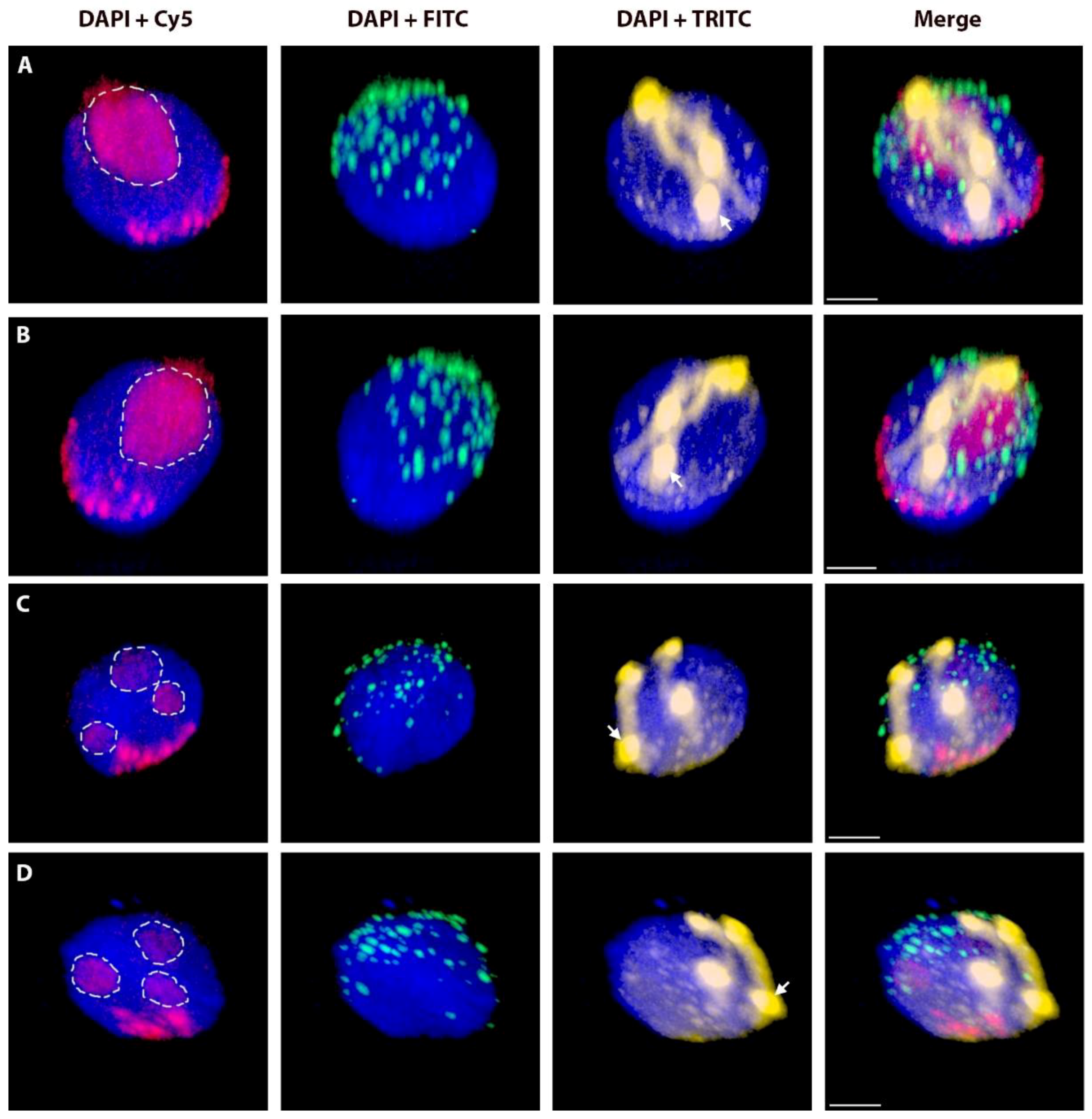
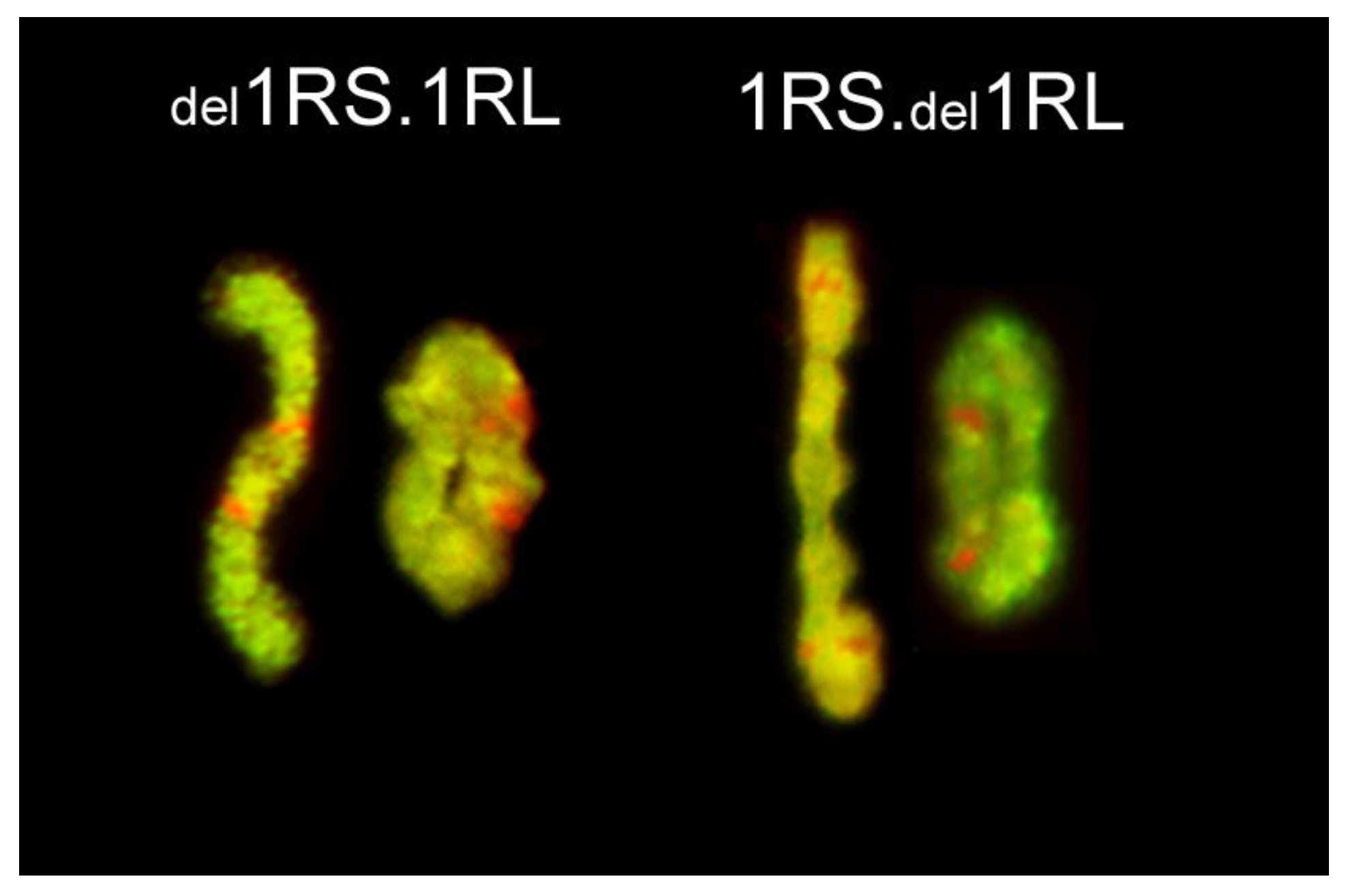
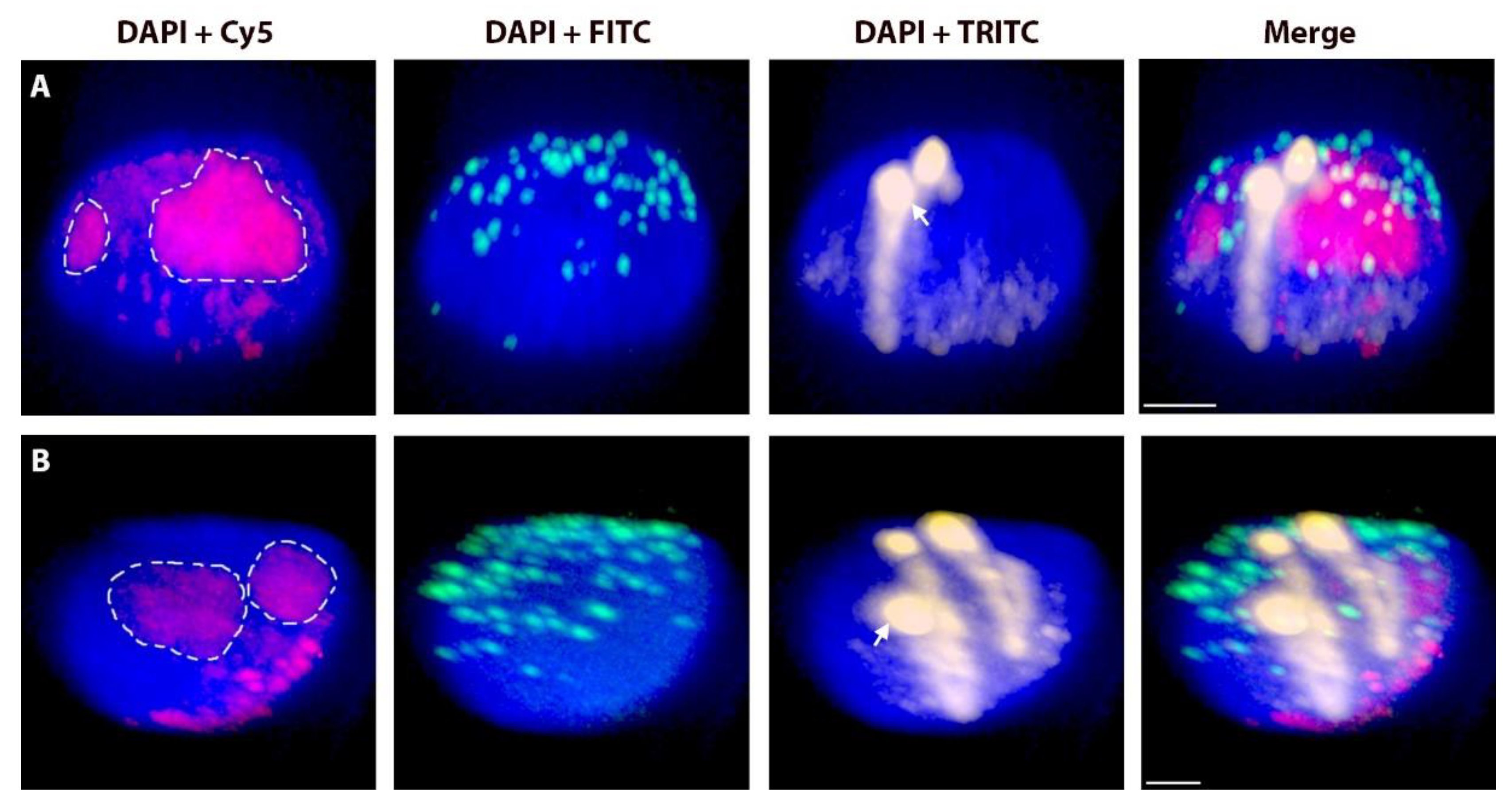
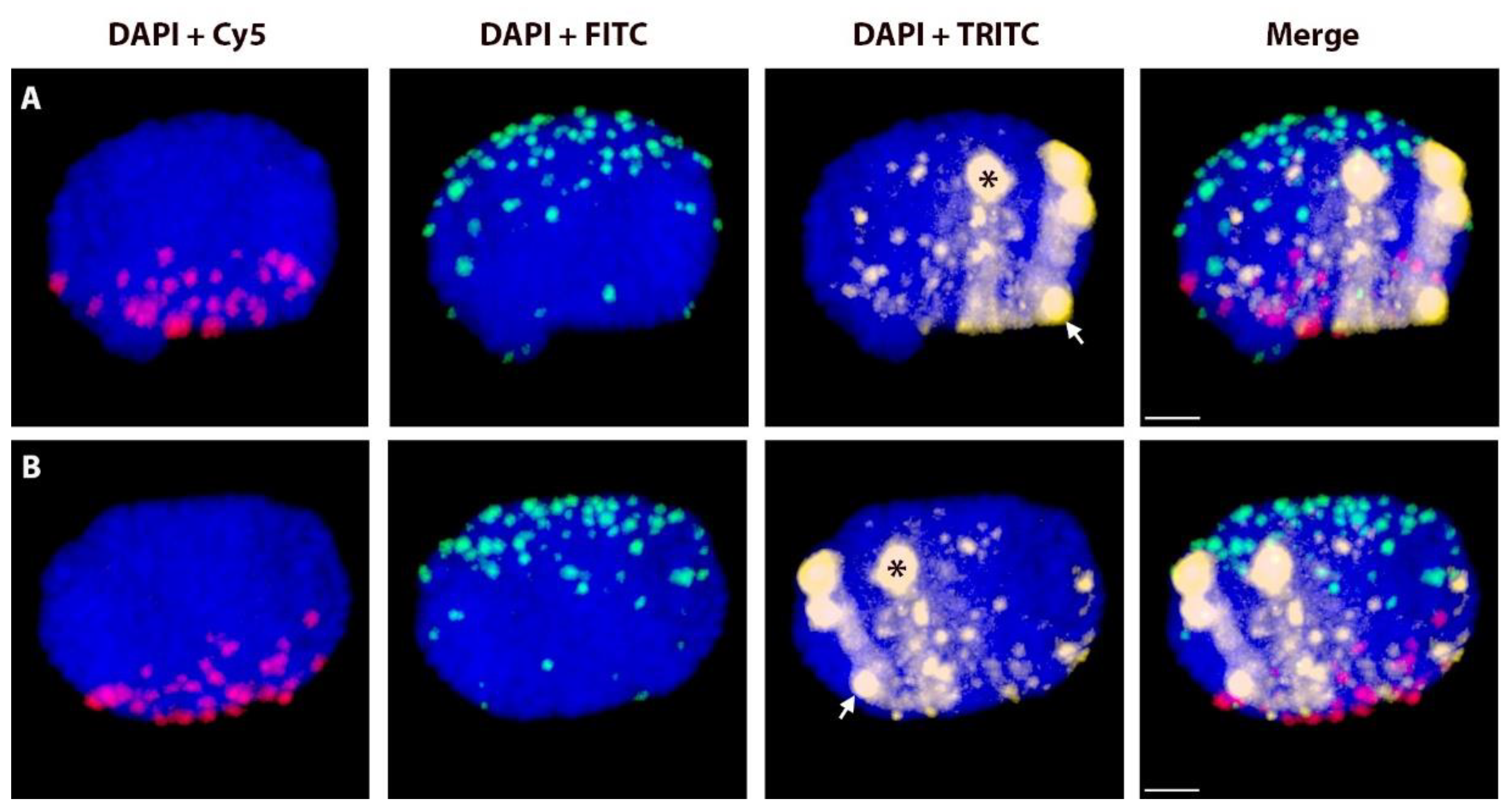
2.2. Positions of Rye Telomeres and Centromeres Relative to the Nuclear Envelope and Positioning of Rye Telomeres in the Telomere Cluster
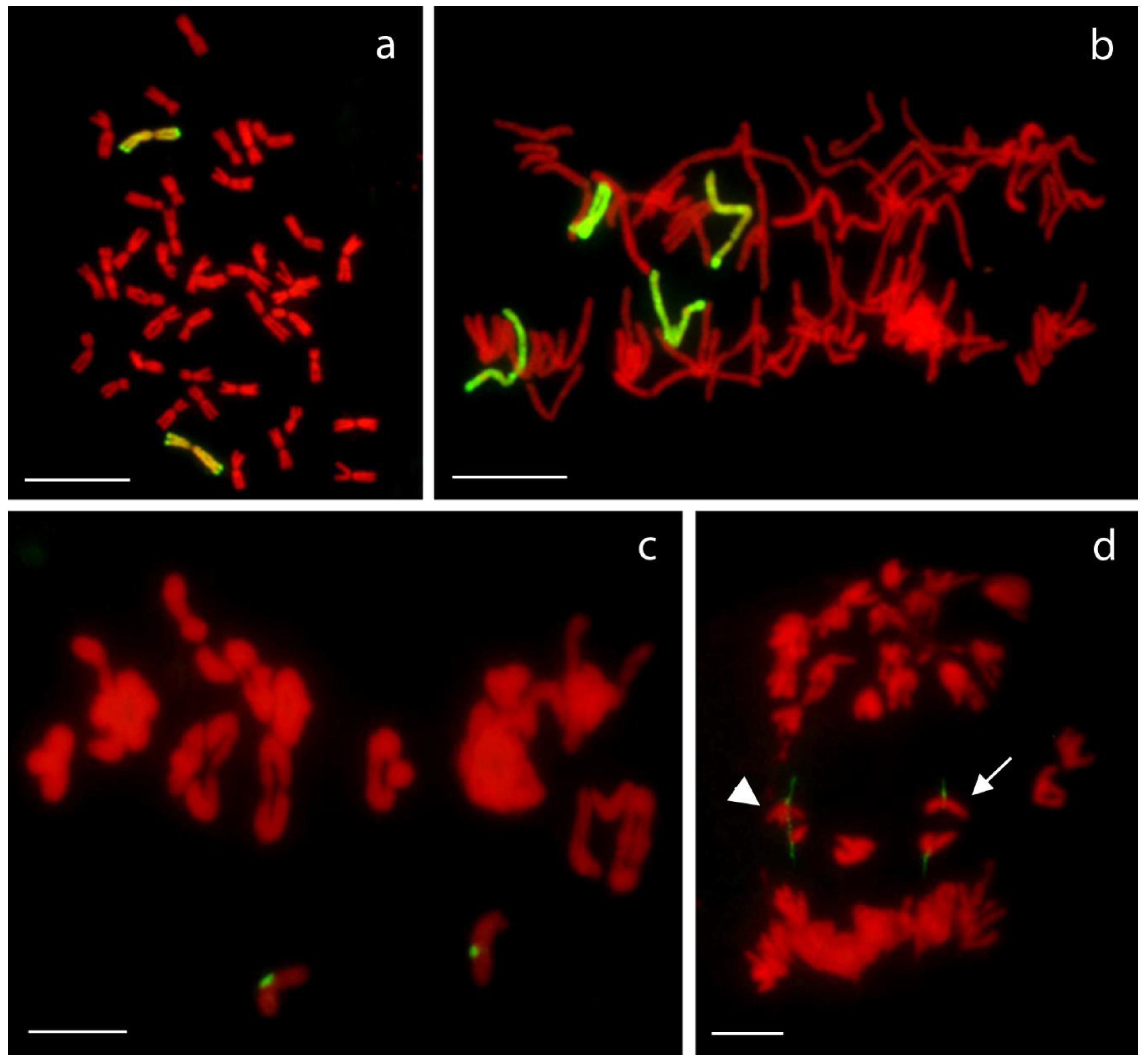
2.3. Chromosome Pairing and Transmission Rate
3. Discussion
4. Materials and Methods
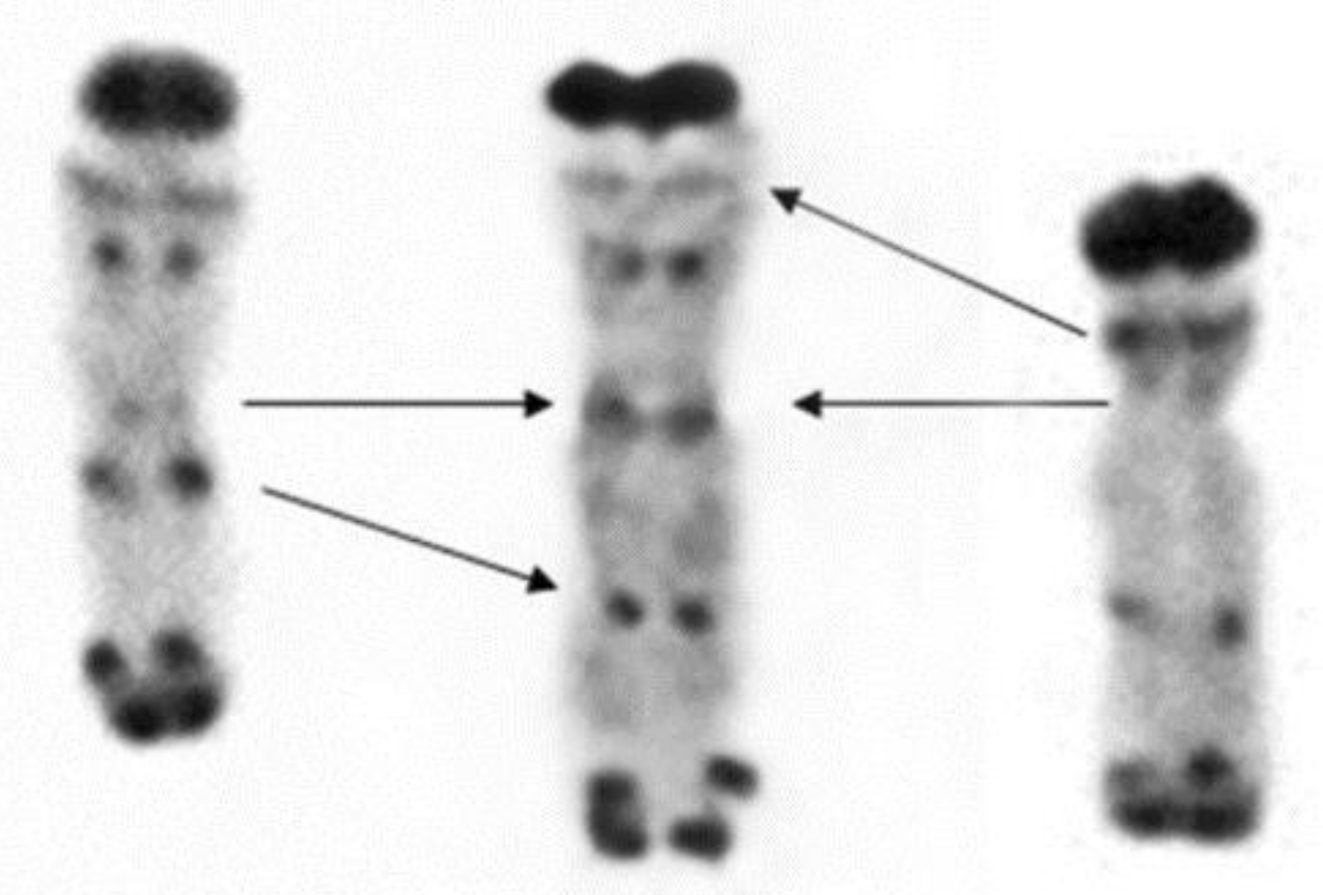
4.1. Plant Material
4.2. Isolation of Nuclei and Flow Sorting
4.3. Probe Preparation and 3D-FISH
4.4. Image Acquisition and Analysis
4.5. Chromosome Pairing and Transmission Rate
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 3D-FISH | Three-dimensional fluorescent in situ hybridization |
| CN | Centre of the nucleus |
| CT | Chromosome territory |
| GISH | Genomic in situ hybridization |
| NE | Nuclear envelope |
| NOR | Nucleolar organizing region |
| NP | Nuclear periphery |
| PMC | Pollen mother cell |
Appendix A


References
- Cremer, T.; Cremer, C. Chromosome territories, nuclear architecture and gene regulation in mammalian cells. Nat. Rev. Genet. 2001, 2, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Fritz, A.J.; Barutcu, A.R.; Martin-Buley, L.; van Wijnen, A.J.; Zaidi, S.K.; Imbalzano, A.N.; Lian, J.B.; Stein, J.L. Chromosomes at Work: Organization of Chromosome Territories in the Interphase Nucleus. J. Cell. Biochem. 2016, 117, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Fransz, P.; de Jong, J.H.; Lysak, M.; Castiglione, M.R.; Schubert, I. Interphase chromosomes in Arabidopsis are organized as well defined chromocenters from which euchromatin loops emanate. Proc. Natl. Acad. Sci. USA 2002, 9, 14584–14589. [Google Scholar] [CrossRef] [PubMed]
- Tiang, C.L.; He, Y.; Pawlowski, W.P. Chromosome Organization and Dynamics during Interphase, Mitosis, and Meiosis in Plants. Plant Physiol. 2012, 158, 26–34. [Google Scholar] [CrossRef]
- Rabl, C. Über Zellteilung. Morph. Jahrb. 1885, 10, 214–330. [Google Scholar]
- Dong, F.G.; Jiang, J.M. Non-Rabl patterns of centromere and telomere distribution in the interphase nuclei of plant cells. Chromosome Res. 1998, 6, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Dawe, R.K. Meiotic chromosome organization and segregation in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 371–395. [Google Scholar] [CrossRef] [PubMed]
- Moens, P.B.; Bernei-Moens, C.; Spyropoulos, B. Chromosome core attachment to the meiotic nuclear envelope regulates synapsis in Chloealtis (Orthoptera). Genome 1989, 32, 601–610. [Google Scholar] [CrossRef]
- Curtis, C.A.; Lukaszewski, A.J.; Chrzastek, M. Metaphase I pairing of deficient chromosomes and genetic mapping of deficiency breakpoints in common wheat. Genome 1991, 34, 553–560. [Google Scholar] [CrossRef]
- Mujeeb-Kazi, A. Intergeneric crosses: Hybrid production and utilization. In Utilizing Wild Grass Biodiversity in Wheat Improvement, 15 Years of Wide cross Research at CIMMYT; Mujeeb-Kazi, A., Hettel, G.P., Eds.; CIMMYT: Texcoco, Mexico, 1995; p. 140. [Google Scholar]
- Friebe, B.; Jiang, J.; Raupp, J.W.; Mclntosh, R.A.; Gill, B.S. Characterization of wheat-alien translocations conferring resistance to diseases and pests: Current status. Euphytica 1996, 91, 59–87. [Google Scholar] [CrossRef]
- Molnár-Láng, M. The crossability of wheat with rye and other related species. In Alien Introgression in Wheat: Cytogenetics, Molecular Biology, and Genomics; Molnár-Láng, M., Ceoloni, C., Doležel, J., Eds.; Springer International Publishing: Chem, Switzerland, 2015; pp. 103–120. [Google Scholar]
- Nishiyama, I. Cytogenetic studies in Avena, IX New synthetic oats in the progenies of induced decaploid interspecific hybrids. Jpn. J. Genet. 1962, 37, 118–130. [Google Scholar] [CrossRef]
- Tsunewaki, K. Genetic studies on a 6x derivative from an 8x triticale. Can. J. Genet. Cytol. 1964, 6, 1–11. [Google Scholar] [CrossRef]
- Lukaszewski, A.J.; Apolinarska, B.; Gustafson, J.P.; Krolow, K.D. Chromosome-pairing and aneuploidy in tetraploid triticale 1. Stabilized karyotypes. Genome 1987, 29, 554–561. [Google Scholar] [CrossRef]
- Orellana, J.; Cermeno, M.C.; Lacadena, J.R. Meiotic pairing in wheat-rye addition and substitution lines. Can. J. Genet. Cytol. 1984, 26, 25–33. [Google Scholar] [CrossRef]
- Lukaszewski, A.J.; Gustafson, J.P. Cytogenetics of Triticale. Plant Breed. Rev. 1987, 5, 41–93. [Google Scholar]
- Murphy, S.P.; Bass, H.W. The maize (Zea mays) desynaptic (dy) mutation defines a pathway for meiotic chromosome segregation, linking nuclear morphology, telomere distribution and synapsis. J. Cell Sci. 2012, 125, 3681–3690. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, T. Dynamics of Rye Telomeres in a Wheat Background during Early Meiosis. Cytogenet. Genome Res. 2014, 143, 60–68. [Google Scholar] [CrossRef]
- Pernickova, K.; Linc, G.; Gaal, E.; Kopecky, D.; Samajova, O.; Lukaszewski, A.J. Out-of-position telomeres in meiotic leptotene appear responsible for chiasmate pairing in an inversion heterozygote in wheat (Triticum aestivum L.). Chromosoma 2019, 128, 31–39. [Google Scholar] [CrossRef]
- Schlegel, R.; Melz, G.; Nestrowicz, R. A universal reference karyotype in rye, Secale cereale L. Theor. Appl. Genet. 1987, 74, 820–826. [Google Scholar] [CrossRef]
- Naranjo, T. Variable Patterning of Chromatin Remodeling, Telomere Positioning, Synapsis, and Chiasma Formation of Individual Rye Chromosomes in Meiosis of Wheat-Rye Additions. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Gill, B.S.; Friebe, B.; Endo, T.R. Standard karyotype and nomenclature system for description of chromosome bands and structural abberations in wheat (Triticum aestivum). Genome 1991, 34, 830–839. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Lucretti, S.; Meister, A.; Lysák, M.A.; Nardi, L.; Obermayer, R. Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann. Bot. 1998, 82, 17–26. [Google Scholar] [CrossRef]
- Lukaszewski, A.J. Unexpected behavior of an inverted rye chromosome arm in wheat. Chromosoma 2008, 117, 569–578. [Google Scholar] [CrossRef]
- Kopecky, D.; Loureiro, J.; Zwierzykowski, Z.; Ghesquiere, M.; Dolezel, J. Genome constitution and evolution in Lolium x Festuca hybrid cultivars (Festulolium). Theor. Appl. Genet. 2006, 113, 731–742. [Google Scholar] [CrossRef]
- Zwierzykowski, Z.; Kosmala, A.; Zwierzykowska, E.; Jones, N.; Joks, W.; Bocianowski, J. Genome balance in six successive generations of the allotetraploid Festuca pratensis x Lolium perenne. Theor. Appl. Genet. 2006, 113, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Alleva, B.; Smolikove, S. Moving and stopping: Regulation of chromosome movement to promote meiotic chromosome pairing and synapsis. Cell 2017, 8, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, R.; Mettin, D. Studies on intraindividual and interindividual variation of chromosome-pairing in diploid and tetraploid populations. 2. interindividual variation. Biol. Zbl. 1975, 94, 703–715. [Google Scholar]
- Schlegel, R. Rye: Genetics, Breeding, and Cultivation; CRC Press: Boca Raton, FL, USA, 2014; p. 359. [Google Scholar]
- Naranjo, T.; Valenzuela, N.T.; Perera, E. Chiasma Frequency Is Region Specific and Chromosome Conformation Dependent in a Rye Chromosome Added to Wheat. Cytogenet. Genome Res. 2010, 129, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Lukaszewski, A.J. Behavior of centromeres in univalents and centric misdivision in wheat. Cytogenet. Genome Res. 2010, 129, 97–109. [Google Scholar] [CrossRef]
- Bridges, C.B. Deficiency. Genetics 1917, 2, 445–465. [Google Scholar]
- Vrana, J.; Simkova, H.; Kubalakova, M.; Cihalikova, J.; Dolezel, J. Flow cytometric chromosome sorting in plants: The next generation. Methods 2012, 57, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Nasuda, S.; Endo, T.R. A direct repeat sequence associated with the centromeric retrotransposons in wheat. Genome 2004, 47, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.; Nibau, C.; Ramsay, L.; Waugh, R.; Jenkins, G. Development of a Molecular Cytogenetic Recombination Assay for Barley. Cytogenet. Genome Res. 2010, 129, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Howe, E.S.; Murphy, S.P.; Bass, H.W. Three-Dimensional Acrylamide Fluorescence in Situ Hybridization for Plant Cells. In Plant Meiosis. Methods and Protocols; Pawlowski, W., Grelon, M., Armstrong, S., Eds.; Springer International Publishing AG: Dordrecht, The Netherlands, 2014; pp. 53–66. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Introgression | Nuclear Volume (µm3) | Arm Length (µm) | C-C (µm) | T-T (µm) |
|---|---|---|---|---|
| 1AS.1RL | 1545 ± 224 | 7.73 ± 1.48 | 4.01 ± 1.98 | 4.88 ± 1.71 |
| 1RS.1BL | 1501 ± 362 | 8.21 ± 1.69 | 3.49 ± 1.50 | 4.18 ± 1.66 |
| 1RS.1DL | 1998 ± 285 | 6.86 ± 1.31 | 2.83 ± 1.15 | 3.71 ± 1.74 |
| 2RS.2BL | 1597 ± 490 | 8.51 ± 2.58 | 3.38 ± 1.70 | 4.09 ± 1.95 |
| 2BS.2RL | 1510 ± 357 | 8.96 ± 1.62 | 4.05 ± 1.80 | 4.99 ± 2.25 |
| 5RS.5BL | 1656 ± 329 | 6.95 ± 1.54 | 3.06 ± 1.29 | 3.97 ± 1.48 |
| del1RS.1RL | 1986 ± 390 | S 1: 4.04 ± 1.03 | 2.88 ± 1.37 | 3.86 ± 1.79 |
| L 2: 7.56 ± 1.67 | 3.56 ± 1.87 | |||
| 1RS. del1RL | 2006 ± 500 | S 1: 6.18 ± 1.58 | 3.57 ± 1.69 | 4.20 ± 1.92 |
| L 2: 5.21 ± 1.41 | 4.12 ± 1.53 |
| Introgression | Rye Chromosome arm Length (Mb) 3 | Chromosome Length (Mb) | Arm Ratio | Number of Nuclei | Telomere Proper Positioning (%) | Telomere in Contact with the NP (%) | Centromere in Contact with the NP (%) |
|---|---|---|---|---|---|---|---|
| 1AS.1RL | 626 | 902 | 2.27 | 25 | 98.00 | 98.00 | 84.00 |
| 1RS.1BL | 423 | 959 | 1.27 | 40 | 98.75 | 98.75 | 100.00 |
| 1RS.1DL | 423 | 804 | 0.90 | 40 | 97.50 | 95.00 | 98.75 |
| 2RS.2BL | 595 | 1102 | 0.85 | 40 | 98.75 | 96.25 | 98.75 |
| 2BS.2RL | 693 | 1116 | 1.64 | 40 | 98.75 | 100.00 | 100.00 |
| 5RS.5BL | 346 | 928 | 1.68 | 40 | 93.75 | 87.50 | 96.25 |
| del1RS.1RL | short arm: 271 1 long arm: 626 | 897 | 2.31 | 25 | 1RS: 82.00 1RL: 100.00 | 1RS: 46.00 1RL: 100.00 | 90.00 |
| 1RS.del1RL | short arm: 423 long arm: 438 2 | 861 | 1.04 | 21 | 1RS: 92.90; 1RL: 100.00 | 1RS: 90.50 1RL: 97.60 | 88.10 |
| 1R(1A) | short arm: 423 long arm: 626 | 1049 | 1.48 | 22 | 1RS: 100.00 1RL: 100.00 | 1RS: 93.20; 1RL: 100.00 | 90.90 |
| t1RS | 423 | 423 | - | 22 | 100.00 | 93.20 | 79.50 |
| Introgression | Number of PMC | Chromosome Pairing | Number of Progeny | Transmission (%) |
|---|---|---|---|---|
| 1AS.1RL | 63 | 100.0 | ||
| 1RS.1BL | 50 | 1RS: 90.0; 1BL: 94.0 | 32 | 100.0 |
| 1RS.1DL | 30 | 100.0 | ||
| 2RS.2BL | 38 | 100.0 | ||
| 2BS.2RL | 49 | 2BS: 95.9; 2RL: 93.9 | 76 | 100.0 |
| 5RS.5BL | 71 | 98.6 | ||
| del1RS.1RL | 60 | 1RS: 90.0; 1RL: 100.0 | 80 | 98.7 |
| 1RS.del1RL | 59 | 1RS: 89.8; 1RL: 94.9 | 80 | 91.9 |
| 1R(1A) | 289 1 | 1RS: 80.8; 1RL: 94.4 | 80 | 99.4 |
| t1RS | 74 | 87.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perničková, K.; Koláčková, V.; Lukaszewski, A.J.; Fan, C.; Vrána, J.; Duchoslav, M.; Jenkins, G.; Phillips, D.; Šamajová, O.; Sedlářová, M.; et al. Instability of Alien Chromosome Introgressions in Wheat Associated with Improper Positioning in the Nucleus. Int. J. Mol. Sci. 2019, 20, 1448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061448
Perničková K, Koláčková V, Lukaszewski AJ, Fan C, Vrána J, Duchoslav M, Jenkins G, Phillips D, Šamajová O, Sedlářová M, et al. Instability of Alien Chromosome Introgressions in Wheat Associated with Improper Positioning in the Nucleus. International Journal of Molecular Sciences. 2019; 20(6):1448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061448
Chicago/Turabian StylePerničková, Kateřina, Veronika Koláčková, Adam J. Lukaszewski, Chaolan Fan, Jan Vrána, Martin Duchoslav, Glyn Jenkins, Dylan Phillips, Olga Šamajová, Michaela Sedlářová, and et al. 2019. "Instability of Alien Chromosome Introgressions in Wheat Associated with Improper Positioning in the Nucleus" International Journal of Molecular Sciences 20, no. 6: 1448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20061448