BvcZR3 and BvHs1pro-1 Genes Pyramiding Enhanced Beet Cyst Nematode (Heterodera schachtii Schm.) Resistance in Oilseed Rape (Brassica napus L.)


Abstract
:1. Introduction
2. Results
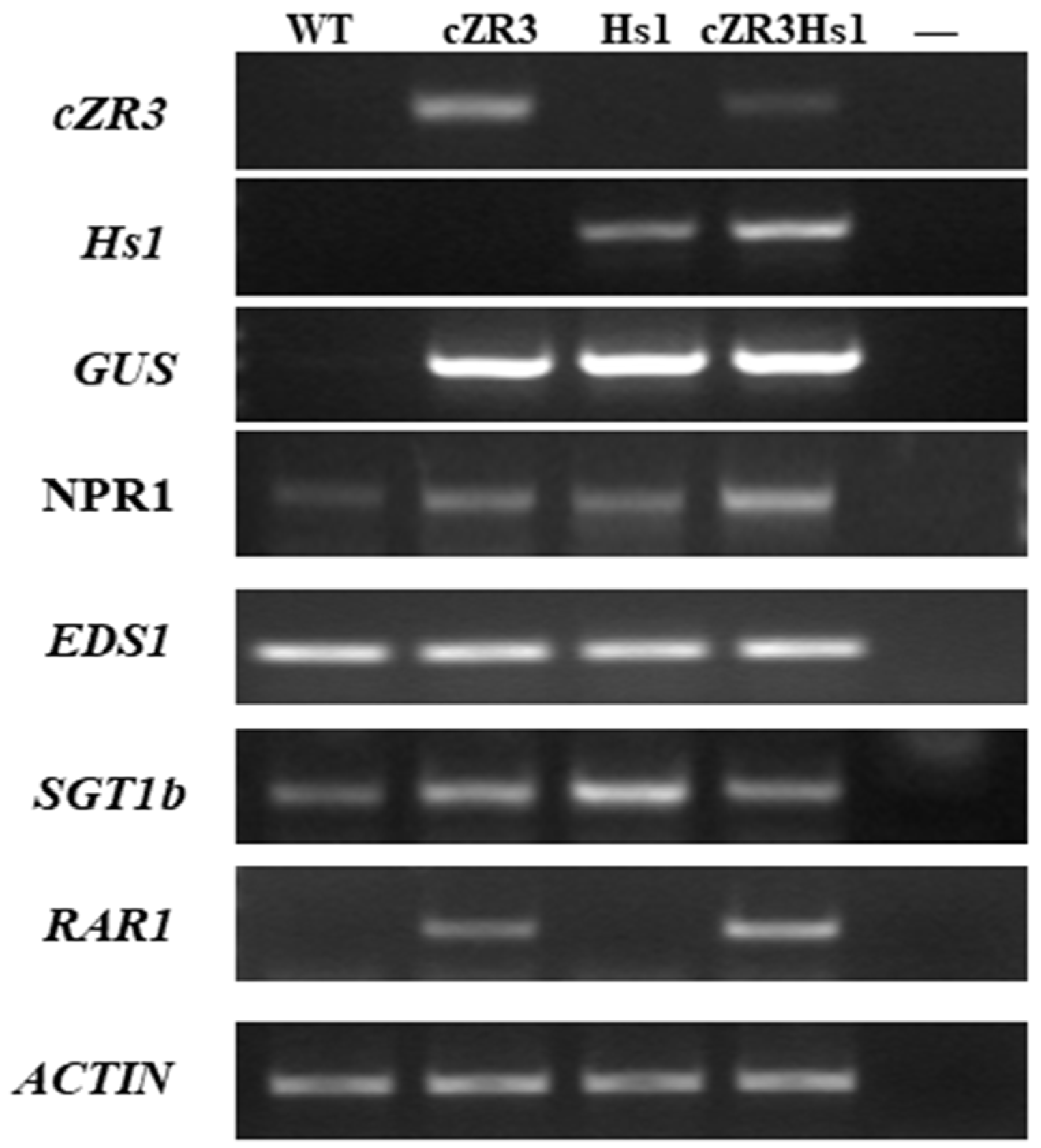
2.1. Generation of Transgenic Plants and Identification
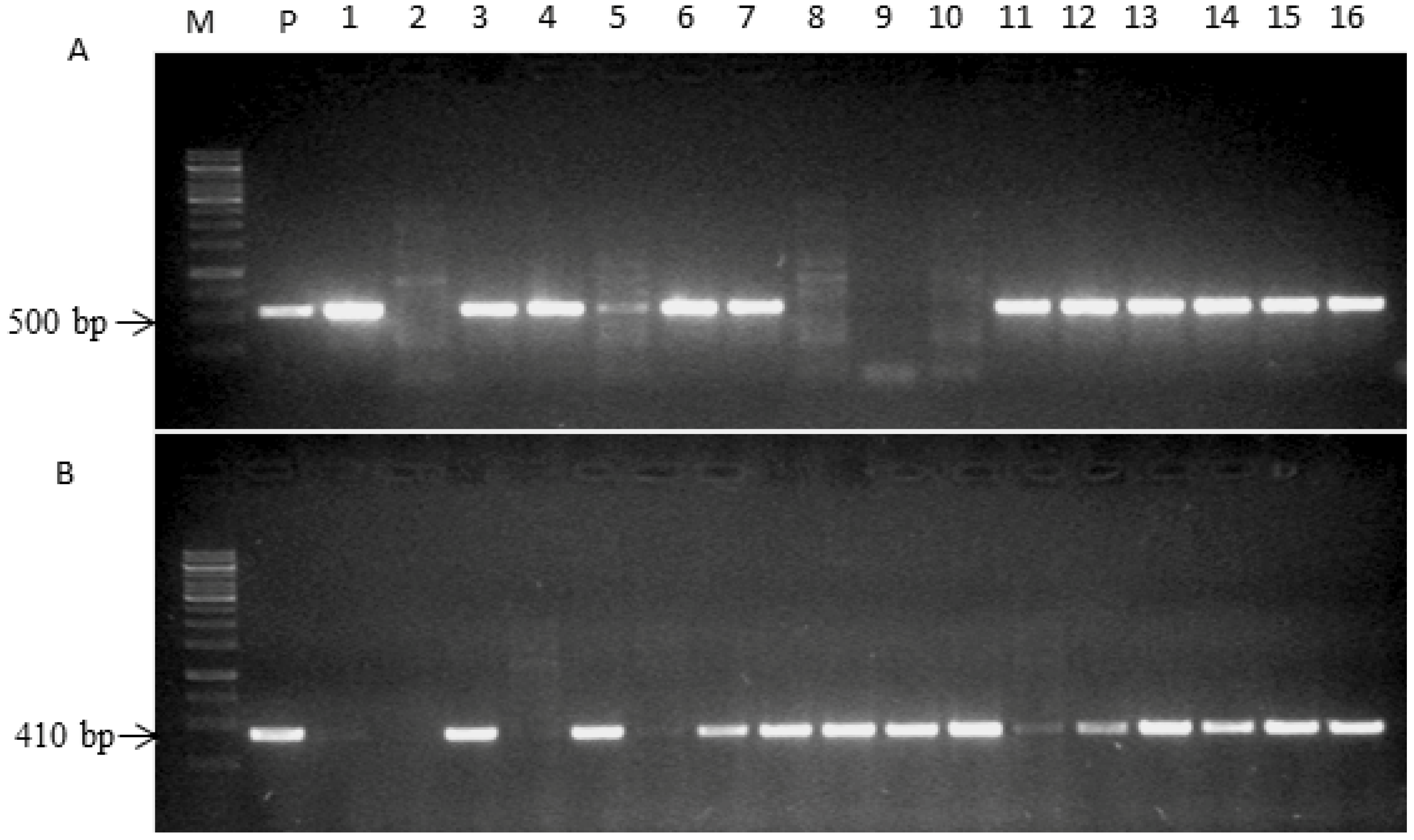
2.2. Generation of cZR3Hs1pro-1 Gene Pyramiding Plants
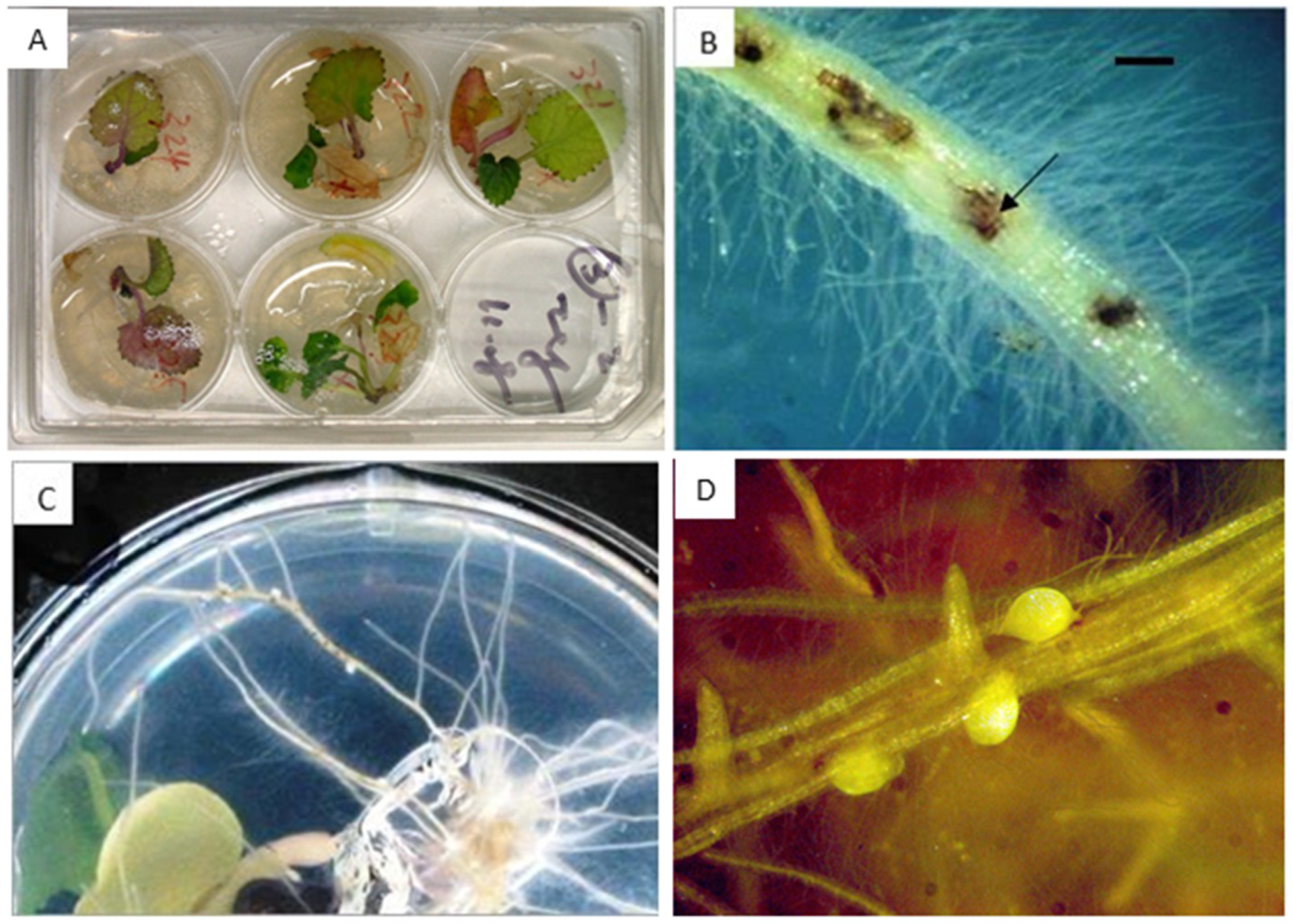
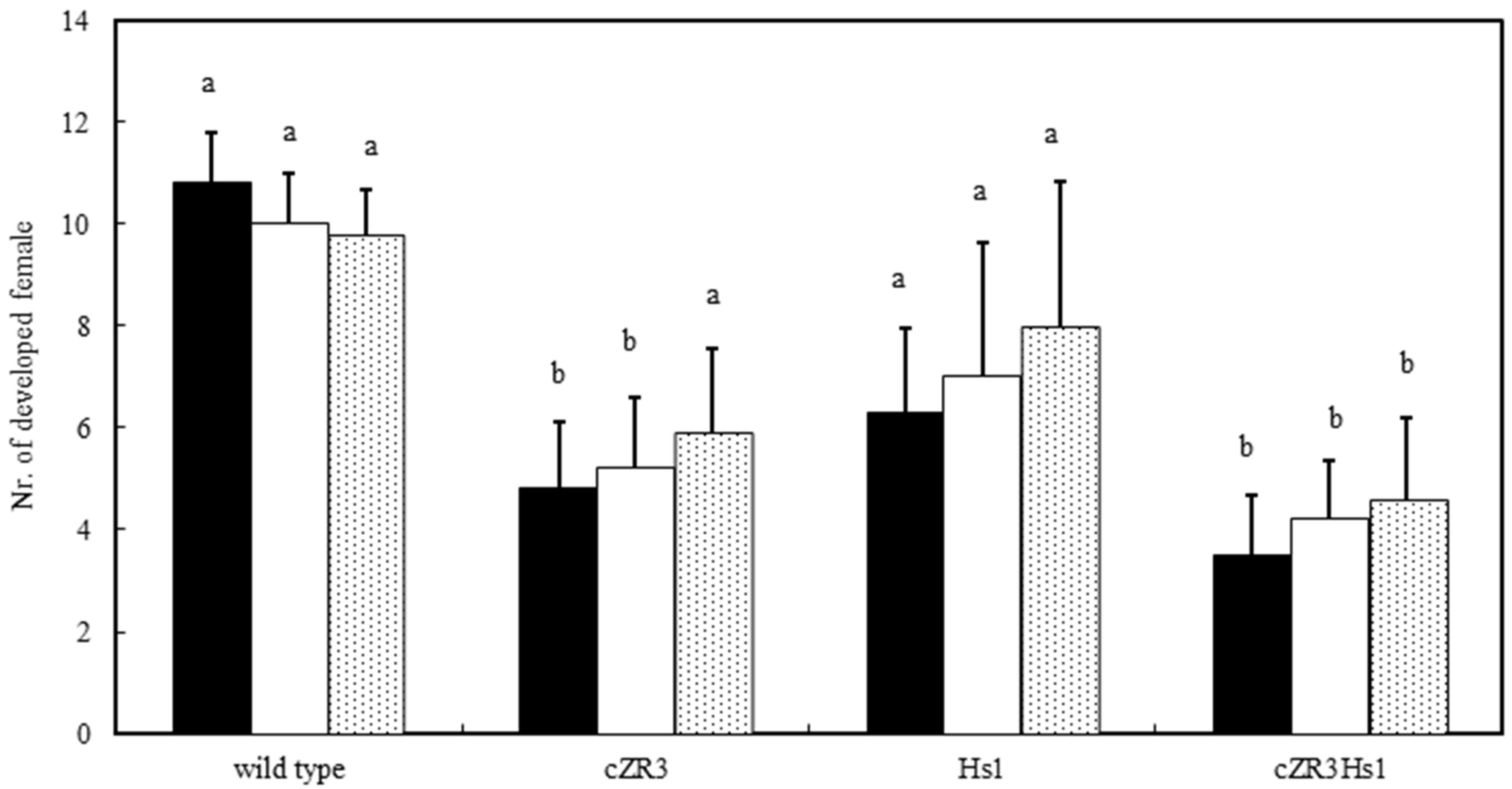
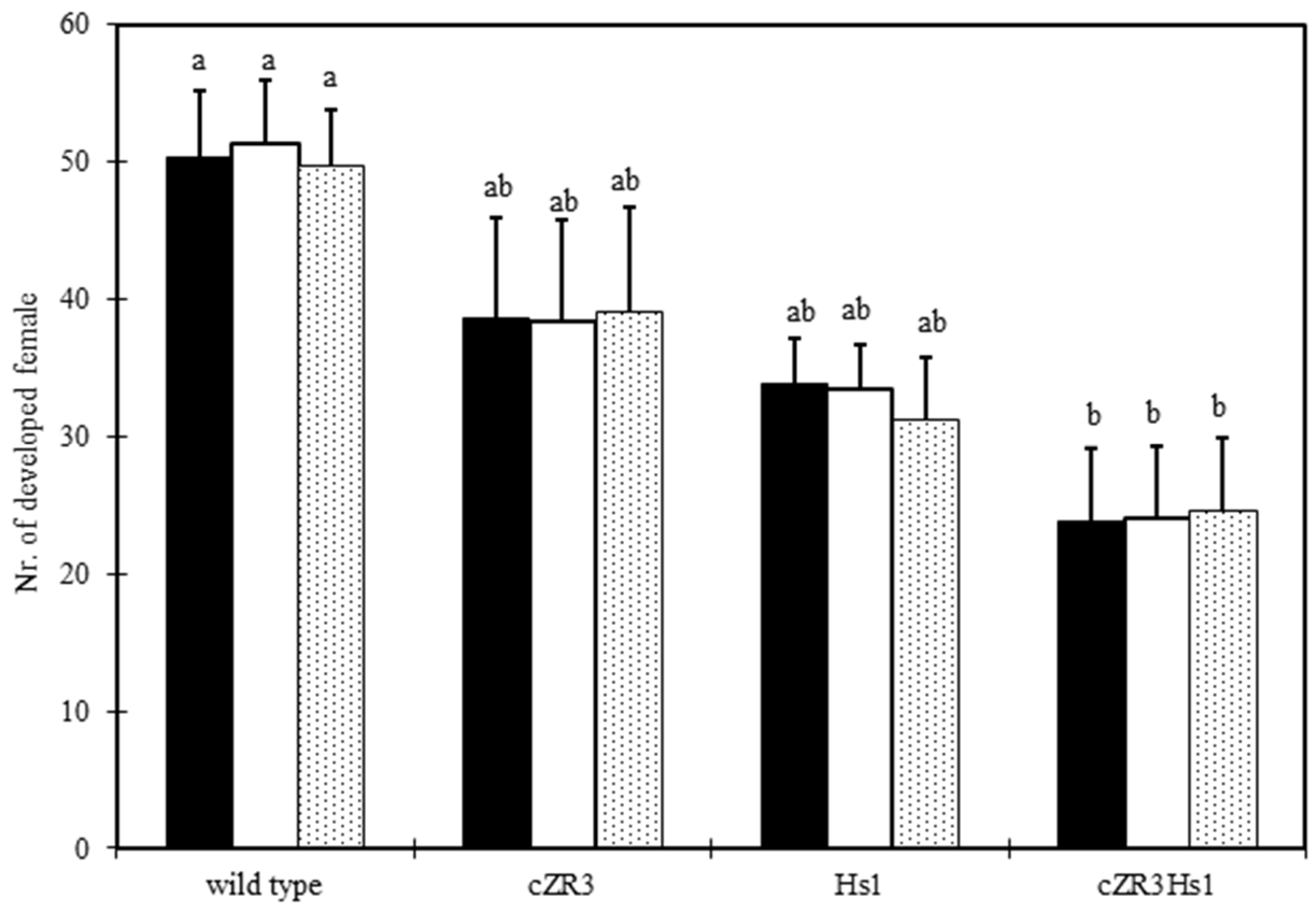
2.3. Beet Cyst Nematode Test In Vitro and In Vivo
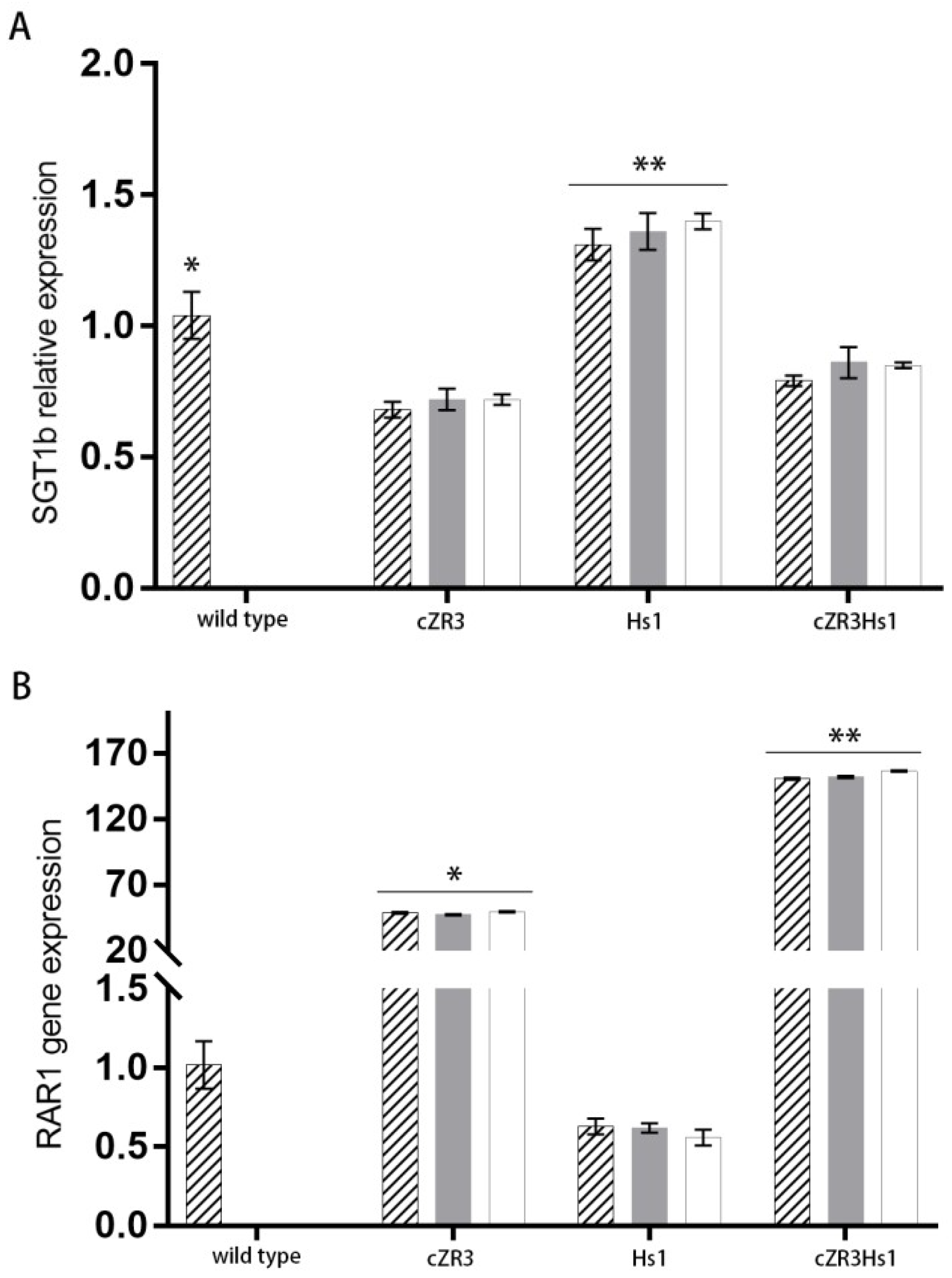
2.4. Determination of RGA-Mediated Signaling Pathways
3. Discussion
4. Materials and Methods
4.1. Plant Transformation
4.2. PCR Analysis
4.3. Southern Blot Analysis
4.4. Histochemical GUS Assays
4.5. Genes Pyramiding and Progeny Analysis
4.6. Propagation of H. Schactii
4.7. Nematode Resistance Assay In Vitro and In Vivo
4.8. Semi Real Time PCR Analysis
4.9. Data Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BCN | beet cyst nematode |
| RGAs | resistance gene analogs |
| CIM | callus induction medium |
| SIM | shoot induced media |
| SEM | shoot elongation media |
| BAP | N-6-benzylaminopurine |
| NAA | Naphthaleneacetic acid |
| AgNO3 | silver nitrate |
| Carb | carbenicillin |
| Kan | kanamycin |
References
- Lilley, C.J.; Atkinson, H.J.; Urwin, P.E. Molecular aspects of cysts nematodes. Mol. Plant Pathol. 2005, 6, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Hemayati, S.S.; Akbar, J.E.; Ghaemi, A.R.; Fasahat, P. Efficiency of white mustard and oilseed radish trap plants against sugar beet cyst nematode. Appl. Soil Ecol. 2017, 119, 192–196. [Google Scholar] [CrossRef]
- Jung, C.; Wyss, U. New approaches to control plant parasitic nematodes. Appl. Microbiol. Biotechnol. 1999, 51, 439–446. [Google Scholar] [CrossRef]
- Lelivelt, C.L.C.; Lange, W. Intergeneric crosses for the transfer of resistance to the beet cyst nematode from Raphanussativus to Brassica napus. Euphytica 1993, 68, 111–120. [Google Scholar] [CrossRef]
- Nielsen, E.L.; Baltenspencer, D.P.; Kerr, E.D.; Rife, C.L. Host suitability of rapeseed for Heterodera schachtii. J. Nematol. 2003, 35, 35–38. [Google Scholar] [PubMed]
- Peterka, H.; Budahn, H.; Schrader, O.; Ahne, R.; Schütze, W. Transfer of resistance against the beet cyst nematode from radish (Raphanussativus) to rape (Brassica napus) by monosomic chromosome addition. Appl. Genet. 2004, 109, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Peterka, H.; Budahn, H.; Zhang, S.S.; Li, J.B. Nematode resistance of rape-radish chromosome addition lines. Nematology 2010, 12, 269–275. [Google Scholar]
- Lelivelt, C.L.C.; Leunissen, E.H.M.; Frederiks, H.J.; Helsper, J.P.F.G.; Krens, F.A. Transfer of resistance to the beet cyst nematode (Heterodera schachtii Schm.) from Sinapis alba L. (white mustard) to the Brassica napus L. gene pool by means of sexual and somatic hybridization. Appl. Genet. 1993, 85, 688–696. [Google Scholar] [CrossRef]
- Brown, J.; Brown, A.P.; Davis, J.B.; Erickson, D. Intergeneric hybridization between Sinapis alba and Brassica napus. Euphytica 1997, 93, 163–168. [Google Scholar] [CrossRef]
- Zhang, S.S.; Peterka, H.; Budahn, H.; Schrader, O.; Li, C.Y. Chromosomal localization of resistance gene in radish against beet cyst nematode and the stability of additional oil radish chromosomes in rape-radish addition lines. Sci. Agric. Sin. 2008, 4, 93–101. [Google Scholar]
- Cai, D.; Kleine, M.; Kifle, S.; Harloff, H.J.; Sandal, N.N.; Marcker, K.A.; Klein-Lankhorst, R.M.; Salentijn, E.M.; Lange, W.; Stiekema, W.J.; et al. Positional cloning of a gene for nematode resistance in sugar beet. Science 1997, 275, 832–834. [Google Scholar] [CrossRef]
- Milligan, S.B.; Bodeau, J.; Yaghoobi, J.; Kaloshian, I.; Zabel, P.; Williamson, V.M. The root knot nematode resistance gene Mi from tomato is a member of the leucine zipper, nucleotide binding, leucine-rich family of plant genes. Plant Cell 1998, 10, 1307–1319. [Google Scholar] [CrossRef]
- Van Der Vossen, E.A.; Van Der Voort, J.N.; Kanyuka, K.; Bendahmane, A.; Sandbrink, H.; Baulcombe, D.C.; Bakker, J.; Stiekema, W.J.; Klein-Lankhorst, R.M. Homologues of a single resistance-gene cluster in potato confer resistance to distinct pathogens: A virus and a nematode. Plant J. 2000, 23, 567–576. [Google Scholar] [CrossRef]
- Ernst, K.; Kumar, A.; Kriseleit, D.; Kloos, D.U.; Phillips, M.S.; Ganal, M.W. The broad-spectrum potato cyst nematode resistance gene (Hero) from tomato is the only member of a large gene family of NBS-LRR genes with an annual amino acid repeat in the LRR region. Plant J. 2002, 31, 127–136. [Google Scholar] [CrossRef]
- Thurau, T.; Kifle, S.; Jung, C.; Cai, D. The promoter of the nematode resistance gene Hs1pro-1 activates a nematode-responsive and feeding site specific gene expression in sugar beet (Beta vulgaris L.) and Arabidopsis thaliana. Plant Mol. Biol. 2003, 52, 643–660. [Google Scholar] [CrossRef]
- McLean, M.D.; Hoover, G.J.; Bancroft, B.; Makhmoudova, A.; Clark, S.M.; Welacky, T.; Simmonds, D.H.; Shelp, B.J. Identification of the full-length Hs1pro-1 coding sequence and preliminary evaluation of soybean cyst nematode resistance in soybean transformed with Hs1pro-1 cDNA. Can. J. Bot. 2007, 85, 437–441. [Google Scholar] [CrossRef]
- Hunger, S.; Gaspero, G.D.; Mohring, S.; Bellin, D.; Schafer-Pregl, R.; Borchardt, D.C.; Durel, C.E.; Werber, M.; Weisshaar, B.; Salamini, F.; et al. Isolation and linkage analysis of expressed disease-resistance gene analogues of sugar beet (Beta vulgaris L.). Genome 2003, 46, 70–82. [Google Scholar] [CrossRef]
- Leister, D.; Kurth, J.; Laurie, D.A.; Yano, M.; Sasaki, T.; Devos, K.; Graner, A.; Schulze-Lefert, P. Rapid reorganization of resistance gene homologues in cereal genomes. Proc. Natl. Acad. Sci. USA 1998, 95, 370–375. [Google Scholar] [CrossRef] [Green Version]
- Timmerman-Vaughan, G.M.; Frew, T.J.; Weeden, N. Characterization and linkage mapping of R-gene analogous DNA sequences in pea (Pisum sativum L.). Appl. Genet. 2000, 101, 241–247. [Google Scholar] [CrossRef]
- Dangl, J.L.; Jones, J.D. Plant pathogens and integrated defence responses to infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef]
- Tian, Y.Y.; Fan, L.J.; Thurau, T.; Jung, C.; Cai, D. The absence of TIR-type resistance gene analogues in the sugar beet (Beta vulgaris L.) genome. Mol. Evol. 2004, 58, 40–53. [Google Scholar] [CrossRef]
- Lein, J.C.; Asbach, K.; Tian, Y.; Schulte, D.; Li, C.; Koch, G.; Jung, C.; Cai, D.G. Resistance gene analogues are clustered on chromosome 3 of sugar beet and cosegregate with QTL for rhizomania resistance. Genome 2007, 50, 61–71. [Google Scholar] [CrossRef]
- Ori, N.; Eshed, Y.; Paran, I.; Presting, G.; Aviv, D.; Tanksley, S.; Zamir, D.; Fluhr, R. The I2C family from the wilt disease resistance locus I2 belongs to the nucleotide binding, leucine-rich repeat superfamily of plant resistance genes. Plant Cell 1997, 9, 521–532. [Google Scholar] [CrossRef]
- Yoshimura, S.; Yamanouchi, U.; Katayose, Y.; Toki, S.; Wang, Z.X.; Kono, I.; Kurata, N.; Yano, M.; Iwata, N.; Sasaki, T. Expression of Xa1, a bacterial blight-resistance gene in rice, is induced by bacterial inoculation. Proc. Natl. Acad. Sci. USA 1998, 95, 1663–1668. [Google Scholar] [CrossRef] [Green Version]
- Lagudah, E.S.; Moullet, O.; Appels, R. Map-based cloning of a gene sequence encoding a nucleotide binding domain and a leucine-rich region at the Cre3 nematode resistance locus of wheat. Genome 1997, 40, 659–665. [Google Scholar] [CrossRef]
- Whitham, S.; Dinesh-Kumar, S.P.; Choi, D.; Hehl, R.; Corr, C.; Baker, B. The product of the tobacco mosaic virus resistance gene N: Similarity to toll and the interleukin-1 receptor. Cell 1994, 78, 1101–1115. [Google Scholar] [CrossRef]
- Dinesh-Kumar, S.P.; Baker, B.J. Alternatively spliced N resistance gene transcripts: Their possible role in tobacco mosaic virus resistance. Proc. Natl. Acad. Sci. USA 2000, 97, 1908–1913. [Google Scholar] [CrossRef] [Green Version]
- Paal, J.; Henselewski, H.; Muth, J.; Meksem, K.; Menéndez, C.M.; Salamini, F.; Ballvora, A.; Gebhardt, C. Molecular cloning of the potato Gro1-4 gene conferring resistance to pathotype Ro1 of the root cyst nematode Globodera rostochiensis, based on a candidate gene approach. Plant. J. 2004, 38, 285–297. [Google Scholar] [CrossRef]
- Calenge, F.; Van der Linden, C.G.; Van de Weg, E.; Schouten, H.J.; Van Arkel, G.; Denance, C.; Durel, C.E. Resistance gene analogues identified through the NBS-profiling method map close to major genes and QTL for disease resistance in apple. Appl. Genet. 2005, 110, 660–668. [Google Scholar] [CrossRef] [Green Version]
- Schulte, D.; Cai, D.; Kleine, M.; Fan, L.; Wang, S.; Jung, C. A complete physical map of wild beet (Beta procumbens) translocation in sugar beet. Mol. Gen. Genom. 2006, 275, 504–511. [Google Scholar] [CrossRef]
- Hammond-Kosack, K.E.; Parker, J.E. Deciphering plant-pathogen communication: Fresh perspectives for molecular resistance breeding. Curr. Opin. Biotechnol. 2003, 14, 177–193. [Google Scholar] [CrossRef]
- Chitwood, D.J. Research on plant-parasitic nematode biology conducted by the United States Department of Agriculture–Agricultural Research Service. Pest. Manag. Sci. 2003, 59, 748–753. [Google Scholar] [CrossRef]
- Hauer, M.; Koch, H.J.; Krüssel, S.; Mittler, S.; Märländer, B. Integrated control of Heterodera schachtii Schmidt in Central Europe by trap crop cultivation, sugar beet variety choice and nematicide application. Appl. Soil Ecol. 2016, 99, 62–69. [Google Scholar] [CrossRef]
- Feechan, A.; Kocsis, M.; Riaz, S.; Zhang, W.; Gadoury, D.M.; Walker, M.A.; Dry, I.B.; Reisch, B.; Cadle-Davidson, L. Strategies for RUN1 deployment using RUN2 and REN2 to manage grapevine powdery mildew informed by studies of race specificity. Phytopathology 2015, 105, 1104–1113. [Google Scholar] [CrossRef]
- Abdeen, A.; Virgòs, A.; Olivella, E.; Villanueva, J.; Avilé, X.; Gabarra, R.; Prat, S. Multiple insect resistance in transgenic tomato plants over-expressing two families of plant proteinase inhibitors. Plant Mol. Biol. 2005, 57, 189–202. [Google Scholar] [CrossRef]
- Bharathia, Y.; Kumara, S.V.; Pasalub, I.C.; Balachandranb, S.M.; Reddya, V.D.; Raoa, K.V. Pyramided rice lines harbouring Allium sativum (asal) and Galanthus nivalis (gna) lectin genes impart enhanced resistance against major sap-sucking pests. J. Biotechnol. 2011, 152, 63–71. [Google Scholar] [CrossRef]
- Chan, Y.L.; He, Y.; Hsiao, T.T.; Wang, C.J.; Tian, Z.H.; Yeh, K.W. Pyramiding taro cystatin and fungal chitinase genes driven by a synthetic promoter enhances resistance in tomato to root-knot nematode Meloidogyne incognita. Plant Sci. 2015, 231, 74–81. [Google Scholar] [CrossRef]
- Urwin, P.E.; McPherson, M.J.; Atkinson, H.J. Enhanced transgenic plant resistance to nematodes by dual proteinase inhibitor constructs. Planta 1998, 204, 472–479. [Google Scholar] [CrossRef]
- Williamson, M.V.; Kumar, A. Nematode resistance in plants: The battle underground. Trends Genet. 2006, 22, 396–403. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Tang, G.X.; Kneck, K.; Yang, X.F.; Qin, Y.B.; Zhou, W.J.; Cai, D. A two-step protocol for shoot regeneration from hypocotyl explants of oilseed rape and its application for Agrobacterium-mediated transformation. Biol. Plant. 2011, 55, 21–26. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays in tobacco tissue culture. Physiol. Plant. 1962, 15, 474–493. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirement of suspensions cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151. [Google Scholar] [CrossRef]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol. Biol. 1985, 5, 69–76. [Google Scholar] [CrossRef]
- Jefferson, R.A. Assaying chimeric gene in plants: The GUS gene fusion system. Plant. Mol. Rep. 1987, 5, 387–405. [Google Scholar] [CrossRef]
- Reski, R.; Abel, W.O. Induction of budding on chloronemata and caulonemata of the moss, Physcomitrella patens, using isopentenyladenine. Planta 1985, 165, 354–358. [Google Scholar] [CrossRef]
- Steiner, A.A. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil 1961, 15, 134–154. [Google Scholar] [CrossRef] [Green Version]
- Sijmons, P.C.; Grundler, F.M.W.; Mende, N.; Burrows, P.R. Arabidopsis thaliana as a new model host for plant parasitic nematodes. Plant J. 2010, 1, 245–254. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and 2-△(△CP) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, D.; Chu, J. The Arabidopsis NPR1 protein is a receptor for the plant defense hormone salicylic acid. Cell Rep. 2012, 1, 639–647. [Google Scholar] [CrossRef]
- Youssef, R.M.; Macdonald, M.H.; Brewer, E.P. Ectopic expression of AtPAD4 broadens resistance of soybean to soybean cyst and root-knot nematodes. BMC Plant Biol. 2013, 13, 67. [Google Scholar] [CrossRef]
- Tornero, P. RAR1 and NDR1 contribute quantitatively to disease resistance in Arabidopsis, and their relative contributions are dependent on the R gene assayed. Plant Cell 2002, 14, 1005–1015. [Google Scholar] [CrossRef]
- Tör, M.; Gordon, P.; Cuzick, A. Arabidopsis SGT1b is required for defense signaling conferred by several downy mildew (Peronospora parasitica) resistance genes. Plant Cell 2002, 14, 993–1003. [Google Scholar] [CrossRef]









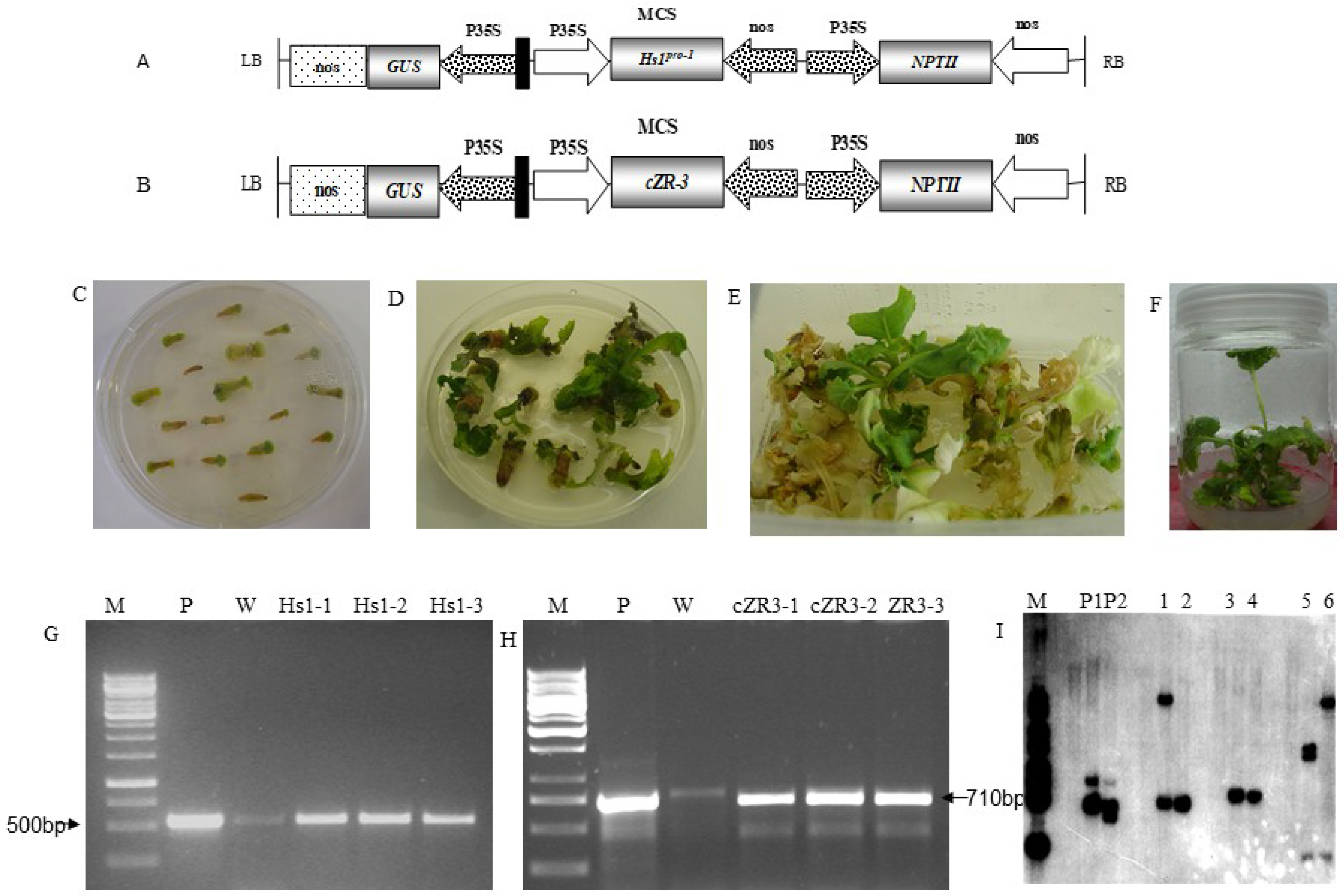{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lines | Number of Plants | Expected Ratio | p-Value | ||
|---|---|---|---|---|---|
| T1 | Hs1pro-1 or cZR3+ | Hs1pro-1 or cZR3− | |||
| Hs1pro-1-1 | 21 | 19 | 2 | 3:1 | p > 0.05 |
| Hs1pro-1-2 | 6 | 1 | 5 | 3:1 | p < 0.05 |
| Hs1pro-1-3 | 18 | 16 | 2 | 3:1 | p > 0.05 |
| cZR-3-1 | 8 | 6 | 2 | 3:1 | p > 0.05 |
| cZR-3-2 | 10 | 8 | 2 | 3:1 | p > 0.05 |
| cZR-3-3 | 37 | 27 | 10 | 3:1 | p > 0.05 |
| Cross Combination | Number of Plants | Expected Ratio | p-Value | ||||
|---|---|---|---|---|---|---|---|
| F2 | cZR3Hs1pro-1+ | cZR3 + | Hs1pro-1+ | cZR3Hs1pro-1− | |||
| cZR3-1 ×Hs1pro-1-2 | 20 | 10 | 3 | 4 | 3 | 9:3:3:1 | p > 0.05 |
| cZR3-2.1 ×Hs1pro-1-3 | 15 | 1 | 12 | 1 | 1 | 9:3:3:1 | p < 0.05 |
| cZR3-2.2 ×Hs1pro-1-3 | 27 | 16 | 4 | 4 | 3 | 9:3:3:1 | p > 0.05 |
| cZR3-3 ×Hs1pro-1-2 | 14 | 6 | 2 | 2 | 4 | 9:3:3:1 | p < 0.05 |
| cZR3-3 ×Hs1pro-1-3 | 35 | 15 | 8 | 9 | 3 | 9:3:3:1 | p > 0.05 |
| Target Gene | Accession Number | Primer Sequence (5′ → 3′) | References |
|---|---|---|---|
| NPR1 | AT1G64280.1 | TGAATTGAAGATGACGCTGCT AGGCCTTCTTTAGTGTCTCTTGTA | Wu et al. 2012 [50] |
| PAD4 | AT3G52430 | GGTCGACGCTGCCATACTCAAACT AGAGAGATTGGTTTCCGAGCAGAG | Youssef et al. 2013 [51] |
| RAR1 | AT5G51700 | CGGCTCCTACTTCATCTCCAG AACATCGCAACATTTCCACCCTCT | Tornero et al. 2002 [52] |
| SGT1b | AT4G11260 | CCCAAACCCAATGTCTCATCAG TCCACTTTCTTAGTCCCAACTTCT | Tör et al. 2002 [53] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, X.; Zhou, Q.; Cui, N.; Cai, D.; Tang, G. BvcZR3 and BvHs1pro-1 Genes Pyramiding Enhanced Beet Cyst Nematode (Heterodera schachtii Schm.) Resistance in Oilseed Rape (Brassica napus L.). Int. J. Mol. Sci. 2019, 20, 1740. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071740
Zhong X, Zhou Q, Cui N, Cai D, Tang G. BvcZR3 and BvHs1pro-1 Genes Pyramiding Enhanced Beet Cyst Nematode (Heterodera schachtii Schm.) Resistance in Oilseed Rape (Brassica napus L.). International Journal of Molecular Sciences. 2019; 20(7):1740. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071740
Chicago/Turabian StyleZhong, Xuanbo, Qizheng Zhou, Nan Cui, Daguang Cai, and Guixiang Tang. 2019. "BvcZR3 and BvHs1pro-1 Genes Pyramiding Enhanced Beet Cyst Nematode (Heterodera schachtii Schm.) Resistance in Oilseed Rape (Brassica napus L.)" International Journal of Molecular Sciences 20, no. 7: 1740. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20071740