Physiological Responses of Jurkat Lymphocytes to Simulated Microgravity Conditions


 ,
,
Abstract
:1. Introduction
2. Results
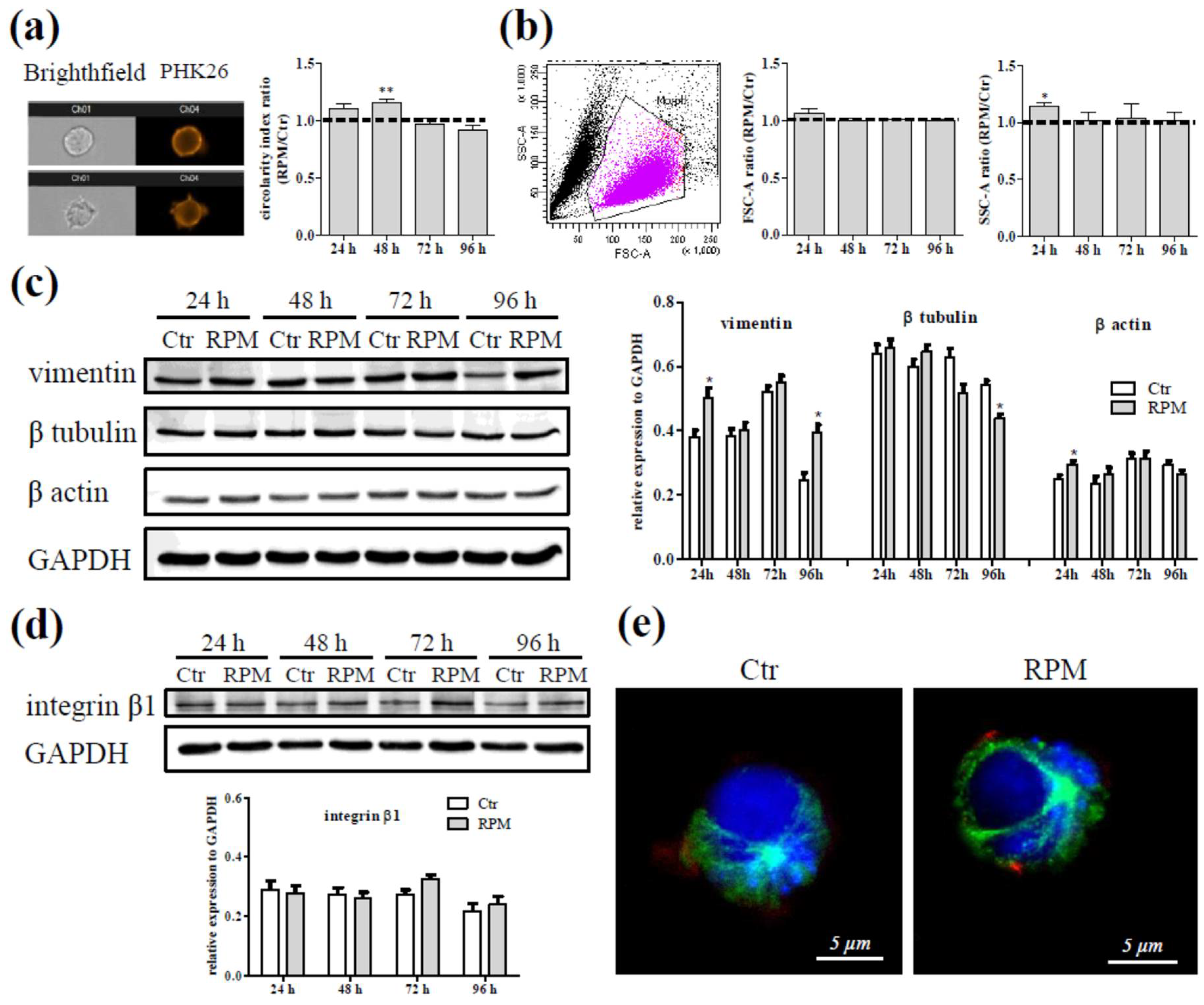
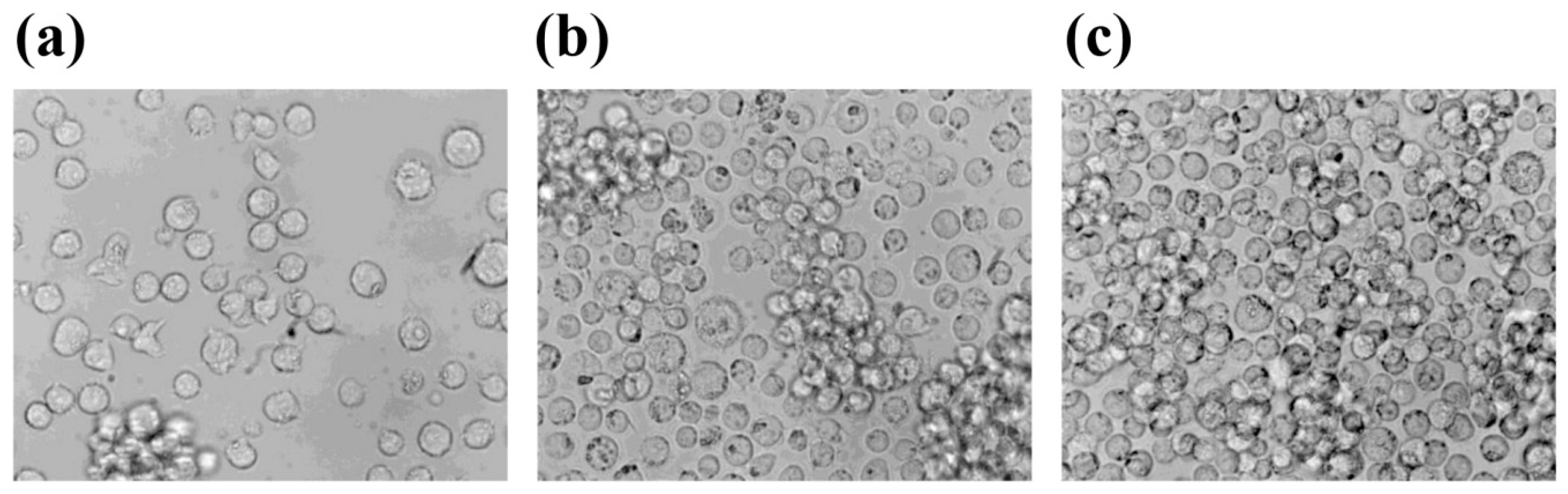
2.1. Cell Shape Aspects Affected by S-Microgravity
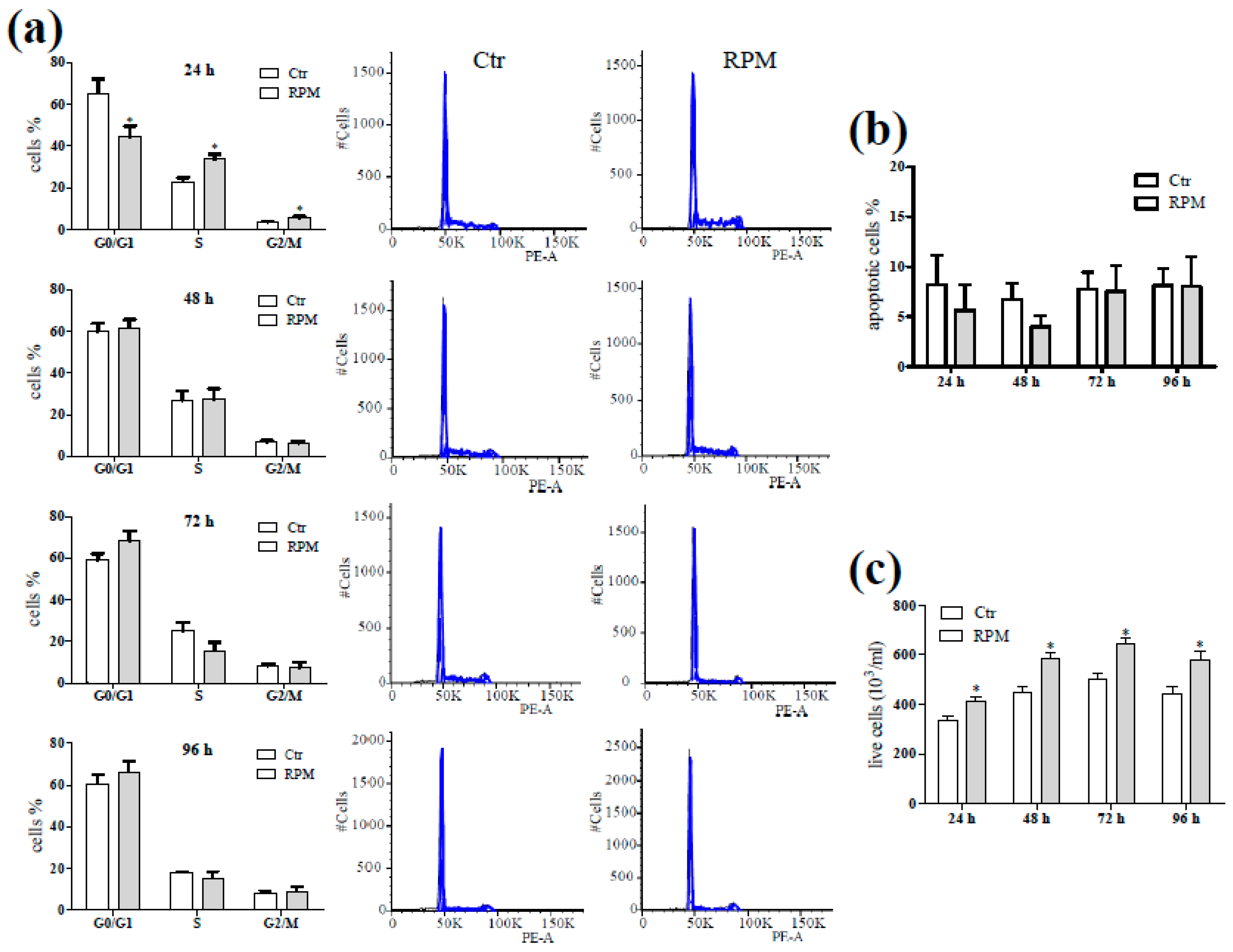
2.2. Biological Features in Presence of S-Microgravity
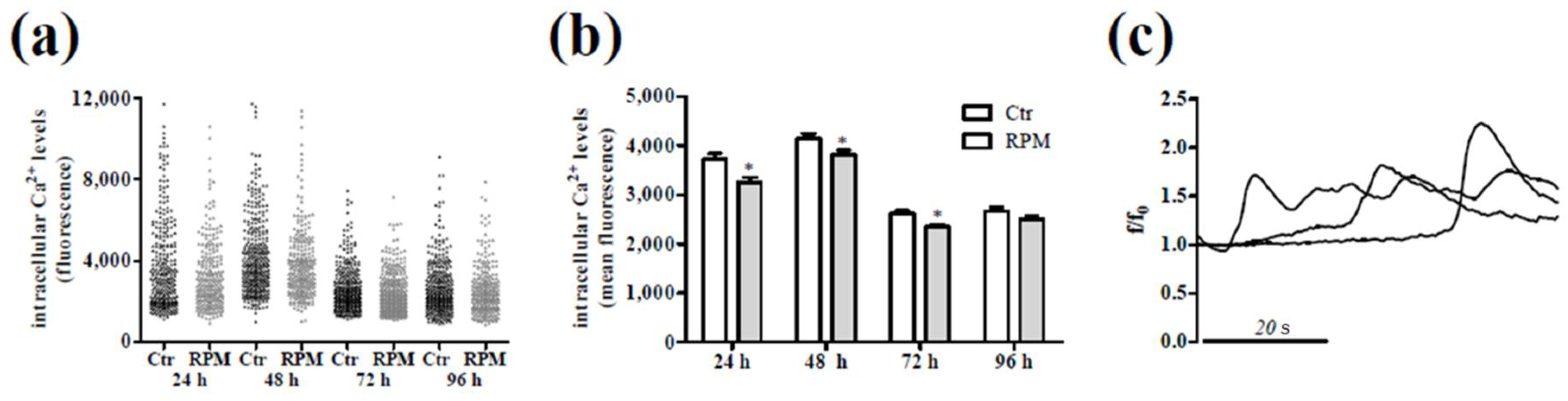
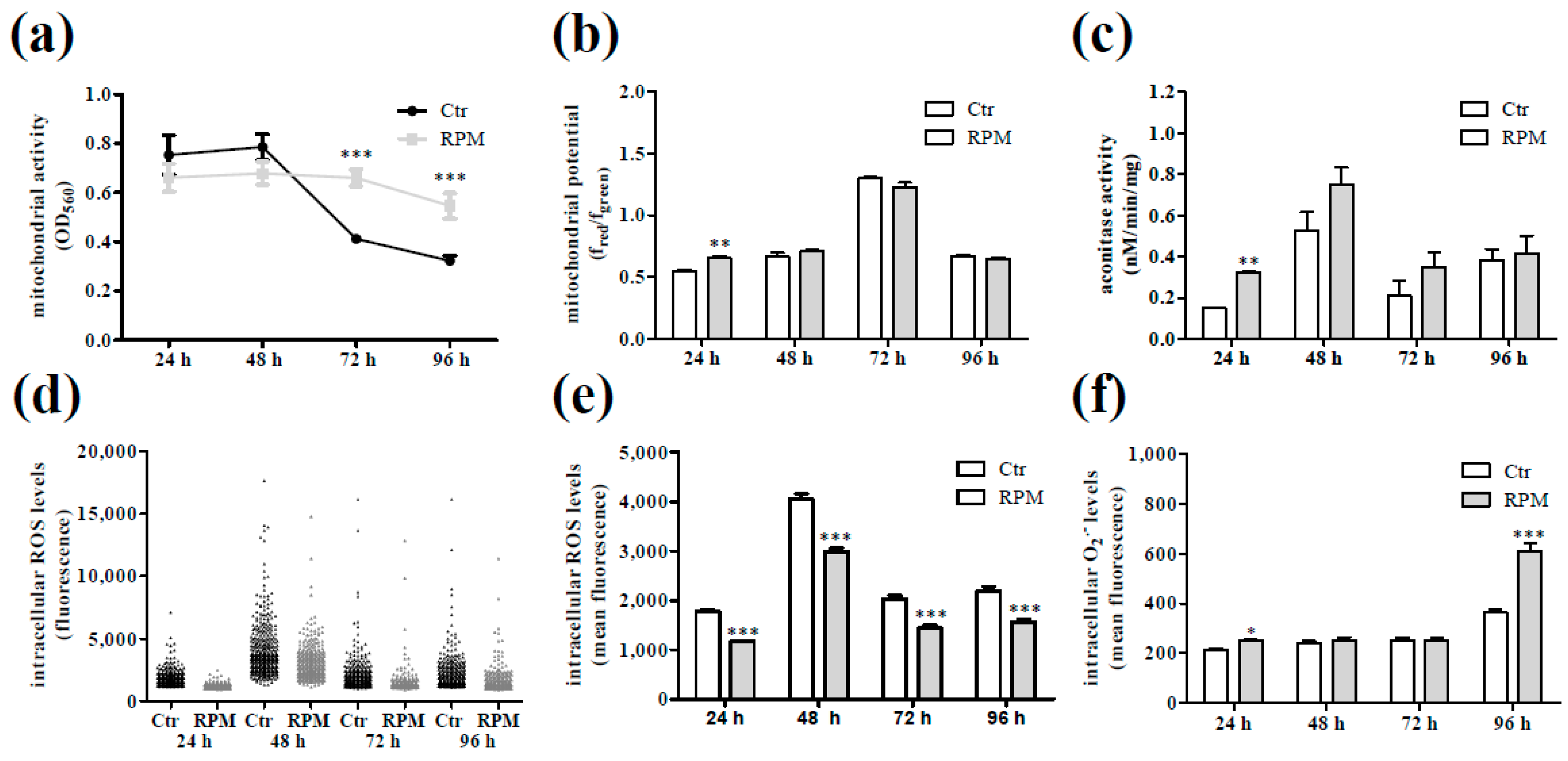
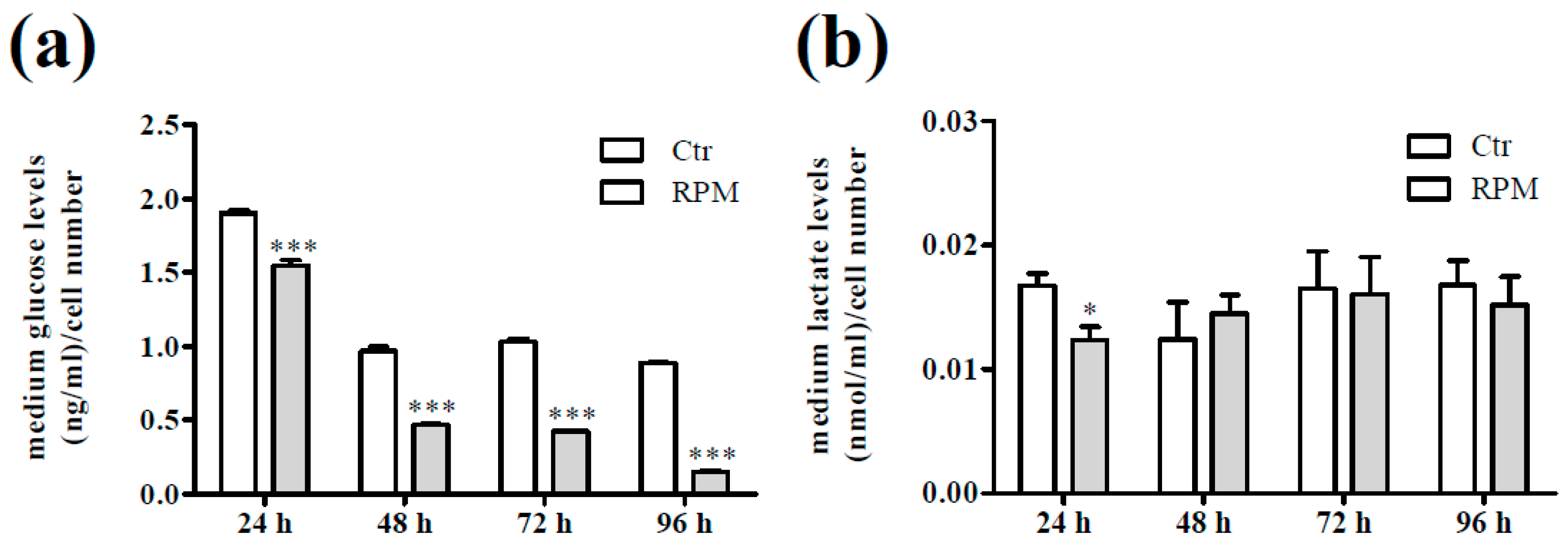
2.3. Intracellular Ca2+ Dynamics and Cell Metabolic Features in Response to S-Microgravity Exposure
3. Discussion
4. Materials and Methods
4.1. Equipment and Cell Exposure Parameters
4.2. Chemicals and Materials
4.3. Cell Cultures
4.4. Viability and Proliferation Assays
4.5. Flow Cytometry and Cell Cycle Analysis
4.6. Cytokine Levels
4.7. Immunofluorescent Cytoskeleton Detection
4.8. ImageStream Analysis of Cell Shape
4.9. Western Blotting
4.10. Measurements of Glucose and Lactate Levels in Cell Culture Medium
4.11. Fluorescence Analyses on Single Cell
4.12. Spectrofluorimetric Measurements
4.13. MTT Assay
4.14. Aconitase Assay
4.15. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ross, M.D. The influence of gravity on structure and function of animals. Adv. Space Res. 1984, 4, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Pietsch, J.; Bauer, J.; Egli, M.; Infanger, M.; Wise, P.; Ulbrich, C.; Grimm, D. The effects of weightlessness on the human organism and mammalian cells. Curr. Mol. Med. 2011, 11, 350–364. [Google Scholar] [CrossRef] [PubMed]
- White, R.J.; Averner, M. Humans in space. Nature 2001, 409, 1115–1118. [Google Scholar] [CrossRef]
- Williams, D.; Kuipers, A.; Mukai, C.; Thirsk, R. Acclimation during space flight: Effects on human physiology. CMAJ 2009, 180, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Hauschild, S.; Tauber, S.; Lauber, B.; Thiel, C.S.; Layer, L.E.; Ullrich, O. T cell regulation in microgravity—The current knowledge from in vitro experiments conducted in space, parabolic flights and ground-based facilities. Acta Astronaut. 2014, 104, 365–377. [Google Scholar] [CrossRef]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.; Lebert, M.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef]
- Hashemi, B.B.; Penkala, J.E.; Vens, C.; Huls, H.; Cubbage, M.; Sams, C.F. T cell activation responses are differentially regulated during clinorotation and in spaceflight. FASEB J. 1999, 13, 2071–2082. [Google Scholar] [CrossRef]
- Martinez, E.M.; Yoshida, M.C.; Candelario, T.L.; Hughes-Fulford, M. Spaceflight and simulated microgravity cause a significant reduction of key gene expression in early t-cell activation. Am. J Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R480–R488. [Google Scholar] [CrossRef]
- Lewis, M.L. The cytoskeleton, apoptosis, and gene expression in t lymphocytes and other mammalian cells exposed to altered gravity. Adv. Space Biol. Med. 2002, 8, 77–128. [Google Scholar] [PubMed]
- Mangala, L.S.; Zhang, Y.; He, Z.; Emami, K.; Ramesh, G.T.; Story, M.; Rohde, L.H.; Wu, H. Effects of simulated microgravity on expression profile of microrna in human lymphoblastoid cells. J. Biol. Chem. 2011, 286, 32483–32490. [Google Scholar] [CrossRef] [PubMed]
- Thiel, C.S.; Huge, A.; Hauschild, S.; Tauber, S.; Lauber, B.A.; Polzer, J.; Paulsen, K.; Lier, H.; Engelmann, F.; Schmitz, B.; et al. Stability of gene expression in human t cells in different gravity environments is clustered in chromosomal region 11p15.4. NPJ Microgravity 2017, 3, 22. [Google Scholar] [CrossRef]
- Cogoli-Greuter, M.; Lovis, P.; Vadrucci, S. Signal transduction in t cells: An overview. J. Gravit. Physiol. 2004, 11, P53–P56. [Google Scholar] [PubMed]
- Fletcher, D.A.; Mullins, R.D. Cell mechanics and the cytoskeleton. Nature 2010, 463, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Valitutti, S.; Dessing, M.; Aktories, K.; Gallati, H.; Lanzavecchia, A. Sustained signaling leading to t cell activation results from prolonged t cell receptor occupancy. Role of t cell actin cytoskeleton. J. Exp. Med. 1995, 181, 577–584. [Google Scholar] [CrossRef]
- Ferranti, F.; Caruso, M.; Cammarota, M.; Masiello, M.G.; Corano Scheri, K.; Fabrizi, C.; Fumagalli, L.; Schiraldi, C.; Cucina, A.; Catizone, A.; et al. Cytoskeleton modifications and autophagy induction in tcam-2 seminoma cells exposed to simulated microgravity. Biomed. Res. Int. 2014, 2014, 904396. [Google Scholar] [CrossRef]
- Vorselen, D.; Roos, W.H.; MacKintosh, F.C.; Wuite, G.J.; van Loon, J.J. The role of the cytoskeleton in sensing changes in gravity by nonspecialized cells. FASEB J. 2014, 28, 536–547. [Google Scholar] [CrossRef] [Green Version]
- Sciola, L.; Cogoli-Greuter, M.; Cogoli, A.; Spano, A.; Pippia, P. Influence of microgravity on mitogen binding and cytoskeleton in jurkat cells. Adv. Space Res. 1999, 24, 801–805. [Google Scholar] [CrossRef]
- Lewis, M.L.; Cubano, L.A.; Zhao, B.; Dinh, H.K.; Pabalan, J.G.; Piepmeier, E.H.; Bowman, P.D. Cdna microarray reveals altered cytoskeletal gene expression in space-flown leukemic t lymphocytes (jurkat). FASEB J. 2001, 15, 1783–1785. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.L.; Reynolds, J.L.; Cubano, L.A.; Hatton, J.P.; Lawless, B.D.; Piepmeier, E.H. Spaceflight alters microtubules and increases apoptosis in human lymphocytes (jurkat). FASEB J. 1998, 12, 1007–1018. [Google Scholar] [CrossRef]
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated microgravity: Critical review on the use of random positioning machines for mammalian cell culture. Biomed. Res. Int. 2015, 2015, 971474. [Google Scholar] [CrossRef]
- Denk, S.; Taylor, R.P.; Wiegner, R.; Cook, E.M.; Lindorfer, M.A.; Pfeiffer, K.; Paschke, S.; Eiseler, T.; Weiss, M.; Barth, E.; et al. Complement c5a-induced changes in neutrophil morphology during inflammation. Scand. J. Immunol. 2017, 86, 143–155. [Google Scholar] [CrossRef]
- Basiji, D.A.; Ortyn, W.E.; Liang, L.; Venkatachalam, V.; Morrissey, P. Cellular image analysis and imaging by flow cytometry. Clin. Lab. Med. 2007, 27, 653–670. [Google Scholar] [CrossRef]
- Cohrs, R.J.; Mehta, S.K.; Schmid, D.S.; Gilden, D.H.; Pierson, D.L. Asymptomatic reactivation and shed of infectious varicella zoster virus in astronauts. J. Med. Virol. 2008, 80, 1116–1122. [Google Scholar] [CrossRef]
- Sonnenfeld, G.; Butel, J.S.; Shearer, W.T. Effects of the space flight environment on the immune system. Rev. Environ. Health 2003, 18, 1–17. [Google Scholar] [CrossRef]
- Sonnenfeld, G.; Shearer, W.T. Immune function during space flight. Nutrition 2002, 18, 899–903. [Google Scholar] [CrossRef]
- Cervantes, J.L.; Hong, B.Y. Dysbiosis and immune dysregulation in outer space. Int. Rev. Immunol. 2016, 35, 67–82. [Google Scholar] [CrossRef]
- Kim, Y.J.; Jeong, A.J.; Kim, M.; Lee, C.; Ye, S.K.; Kim, S. Time-averaged simulated microgravity (tasmg) inhibits proliferation of lymphoma cells, l-540 and hdlm-2, using a 3d clinostat. Biomed. Eng. Online. 2017, 16, 48. [Google Scholar] [CrossRef]
- Morabito, C.; Guarnieri, S.; Catizone, A.; Schiraldi, C.; Ricci, G.; Mariggio, M.A. Transient increases in intracellular calcium and reactive oxygen species levels in tcam-2 cells exposed to microgravity. Sci. Rep. 2017, 7, 15648. [Google Scholar] [CrossRef]
- Hauslage, J.; Cevik, V.; Hemmersbach, R. Pyrocystis noctiluca represents an excellent bioassay for shear forces induced in ground-based microgravity simulators (clinostat and random positioning machine). NPJ Microgravity 2018, 3, 12. [Google Scholar] [CrossRef]
- Wuest, S.L.; Stern, P.; Casartelli, E.; Egli, M. Fluid dynamics appearing during simulated microgravity using random positioning machines. PLoS ONE 2017, 12, e0170826. [Google Scholar] [CrossRef]
- Balakrishnan, S.; Rao, S.B. Cytogenetic analysis of peripheral blood lymphocytes of occupational workers exposed to low levels of ionising radiation. Mutat. Res. 1999, 442, 37–42. [Google Scholar] [CrossRef]
- Blakely, W.F.; Miller, A.C.; Grace, M.B.; McLeland, C.B.; Luo, L.; Muderhwa, J.M.; Miner, V.L.; Prasanna, P.G. Radiation biodosimetry: Applications for spaceflight. Adv. Space Res. 2003, 31, 1487–1493. [Google Scholar] [CrossRef]
- Mariggio, M.A.; Falone, S.; Morabito, C.; Guarnieri, S.; Mirabilio, A.; Pilla, R.; Bucciarelli, T.; Verratti, V.; Amicarelli, F. Peripheral blood lymphocytes: A model for monitoring physiological adaptation to high altitude. High Alt. Med. Biol. 2010, 11, 333–342. [Google Scholar] [CrossRef]
- Morabito, C.; Bosco, G.; Pilla, R.; Corona, C.; Mancinelli, R.; Yang, Z.; Camporesi, E.M.; Fano, G.; Mariggio, M.A. Effect of pre-breathing oxygen at different depth on oxidative status and calcium concentration in lymphocytes of scuba divers. Acta Physiol. (Oxf.) 2011, 202, 69–78. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, Y.; Wang, F. Effects of a 28-day “living high--training low” on t-lymphocyte subsets in soccer players. Int. J. Sports Med. 2007, 28, 354–358. [Google Scholar] [CrossRef]
- Aleshcheva, G.; Wehland, M.; Sahana, J.; Bauer, J.; Corydon, T.J.; Hemmersbach, R.; Frett, T.; Egli, M.; Infanger, M.; Grosse, J.; et al. Moderate alterations of the cytoskeleton in human chondrocytes after short-term microgravity produced by parabolic flight maneuvers could be prevented by up-regulation of bmp-2 and sox-9. FASEB J. 2015, 29, 2303–2314. [Google Scholar] [CrossRef]
- Dominguez, R.; Holmes, K.C. Actin structure and function. Annu. Rev. Biophys. 2011, 40, 169–186. [Google Scholar] [CrossRef]
- Steward, A.J.; Wagner, D.R.; Kelly, D.J. The pericellular environment regulates cytoskeletal development and the differentiation of mesenchymal stem cells and determines their response to hydrostatic pressure. Eur. Cell Mater. 2013, 25, 167–178. [Google Scholar] [CrossRef]
- Bizzarri, M.; Masiello, M.G.; Giuliani, A.; Cucina, A. Gravity constraints drive biological systems toward specific organization patterns: Commitment of cell specification is constrained by physical cues. Bioessays 2018, 40. [Google Scholar] [CrossRef]
- Hellweg, C.E.; Dilruba, S.; Adrian, A.; Feles, S.; Schmitz, C.; Berger, T.; Przybyla, B.; Briganti, L.; Franz, M.; Segerer, J.; et al. Space experiment “cellular responses to radiation in space (cellrad)”: Hardware and biological system tests. Life Sci. Space Res. (Amst) 2015, 7, 73–89. [Google Scholar] [CrossRef]
- Pippia, P.; Galleri, G.; Meloni, M.A.; Saba, A.; Pani, G.; Cogoli-Greuter, M.; Cogoli, A. Effects of microgravity and cosmic radiations on human t lymphocytes. J. Biol. Res. 2009, 82, 19–21. [Google Scholar] [CrossRef]
- Bergqvist, A.; Sundstrom, S.; Dimberg, L.Y.; Gylfe, E.; Masucci, M.G. The hepatitis c virus core protein modulates t cell responses by inducing spontaneous and altering t-cell receptor-triggered Ca2+ oscillations. J. Biol. Chem. 2003, 278, 18877–18883. [Google Scholar] [CrossRef]
- Wacholtz, M.C.; Lipsky, P.E. Anti-cd3-stimulated Ca2+ signal in individual human peripheral t cells. Activation correlates with a sustained increase in intracellular Ca2+. J. Immunol. 1993, 150, 5338–5349. [Google Scholar]
- Babich, A.; Burkhardt, J.K. Coordinate control of cytoskeletal remodeling and calcium mobilization during t-cell activation. Immunol. Rev. 2013, 256, 80–94. [Google Scholar] [CrossRef]
- Joseph, N.; Reicher, B.; Barda-Saad, M. The calcium feedback loop and t cell activation: How cytoskeleton networks control intracellular calcium flux. Biochim. Biophys. Acta 2014, 1838, 557–568. [Google Scholar] [CrossRef]
- Quintana, A.; Hoth, M. Mitochondrial dynamics and their impact on t cell function. Cell Calcium. 2012, 52, 57–63. [Google Scholar] [CrossRef]
- Tauber, S.; Lauber, B.A.; Paulsen, K.; Layer, L.E.; Lehmann, M.; Hauschild, S.; Shepherd, N.R.; Polzer, J.; Segerer, J.; Thiel, C.S.; et al. Cytoskeletal stability and metabolic alterations in primary human macrophages in long-term microgravity. PLoS ONE 2017, 12, e0175599. [Google Scholar] [CrossRef]
- Trepat, X.; Sahai, E. Mesoscale physical principles of collective cell organization. Nat. Phys. 2018, 14, 671–682. [Google Scholar] [CrossRef]
- Borst, A.G.; van Loon, J.J.W.A. Technology and developments for the random positioning machine, rpm. Microgravity Sci. Technol. 2009, 21, 287–292. [Google Scholar] [CrossRef]
- Guarnieri, S.; Pilla, R.; Morabito, C.; Sacchetti, S.; Mancinelli, R.; Fano, G.; Mariggio, M.A. Extracellular guanosine and gtp promote expression of differentiation markers and induce s-phase cell-cycle arrest in human sh-sy5y neuroblastoma cells. Int. J. Dev. Neurosci. 2009, 27, 135–147. [Google Scholar] [CrossRef]
- Lanuti, P.; Santilli, F.; Marchisio, M.; Pierdomenico, L.; Vitacolonna, E.; Santavenere, E.; Iacone, A.; Davi, G.; Romano, M.; Miscia, S. A novel flow cytometric approach to distinguish circulating endothelial cells from endothelial microparticles: Relevance for the evaluation of endothelial dysfunction. J. Immunol. Methods 2012, 380, 16–22. [Google Scholar] [CrossRef]
- Morabito, C.; Lanuti, P.; Caprara, G.A.; Guarnieri, S.; Verratti, V.; Ricci, G.; Catizone, A.; Marchisio, M.; Fano-Illic, G.; Mariggio, M.A. Responses of peripheral blood mononuclear cells to moderate exercise and hypoxia. Scand. J. Med. Sci. Sports 2016, 26, 1188–1199. [Google Scholar] [CrossRef]
- Caprara, G.A.; Morabito, C.; Perni, S.; Navarra, R.; Guarnieri, S.; Mariggio, M.A. Evidence for altered Ca2+ handling in growth associated protein 43-knockout skeletal muscle. Front. Physiol. 2016, 7, 493. [Google Scholar] [CrossRef]
- Yan, L.J.; Levine, R.L.; Sohal, R.S. Oxidative damage during aging targets mitochondrial aconitase. Proc. Natl. Acad. Sci. USA 1997, 94, 11168–11172. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Oscillating Cells (% ± SEM) | Frequency of Ca2+ Oscillations (Peaks/min ± SEM) | Amplitude of Ca2+ Oscillations (Ratio Peakmax/Baseline) | Tested Cells (Number) | ||||
|---|---|---|---|---|---|---|---|---|
| Ctr | RPM | Ctr | RPM | Ctr | RPM | Ctr | RPM | |
| 24 h | 49.3 ± 13.7 | 49.3 ± 9.4 | 1.06 ± 0.02 | 1.03 ± 0.02 | 1.83 ± 0.07 | 1.45 ± 0.04 *** | 312 | 254 |
| 48 h | 43.0 ± 8.0 | 41.3 ± 3.3 | 1.06 ± 0.02 | 1.05 ± 0.02 | 1.40 ± 0.03 | 1.28 ± 0.02 ** | 364 | 277 |
| 72 h | 55.3 ± 6.1 | 41.3 ± 7.5 | 1.15 ± 0.03 | 1.15 ± 0.03 | 1.31 ± 0.02 | 1.25 ± 0.02 * | 357 | 406 |
| 96 h | 53.1 ± 5.5 | 55.3 ± 7.7 | 1.12 ± 0.01 | 1.12 ± 0.03 | 1.29 ± 0.02 | 1.25 ± 0.02 | 346 | 294 |
| Probe | Excitation (nm) | Emission (nm) | Analyses |
|---|---|---|---|
| Fluo4-AM 5 µM | 488 | 520 | Intracellular Ca2+ levels |
| H2-DCFDA 10 µM | 488 | 520 | Intracellular ROS levels |
| MitoSox RED 5 µM | 510 | 580 | Mitochondrial O2− levels |
| JC1 5 µg/mL | 488 | 520/590 | Mitochondrial membrane potential |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morabito, C.; Lanuti, P.; Caprara, G.A.; Marchisio, M.; Bizzarri, M.; Guarnieri, S.; Mariggiò, M.A. Physiological Responses of Jurkat Lymphocytes to Simulated Microgravity Conditions. Int. J. Mol. Sci. 2019, 20, 1892. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20081892
Morabito C, Lanuti P, Caprara GA, Marchisio M, Bizzarri M, Guarnieri S, Mariggiò MA. Physiological Responses of Jurkat Lymphocytes to Simulated Microgravity Conditions. International Journal of Molecular Sciences. 2019; 20(8):1892. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20081892
Chicago/Turabian StyleMorabito, Caterina, Paola Lanuti, Giusy A. Caprara, Marco Marchisio, Mariano Bizzarri, Simone Guarnieri, and Maria A. Mariggiò. 2019. "Physiological Responses of Jurkat Lymphocytes to Simulated Microgravity Conditions" International Journal of Molecular Sciences 20, no. 8: 1892. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20081892