HDAC5 Expression in Urothelial Carcinoma Cell Lines Inhibits Long-Term Proliferation but Can Promote Epithelial-to-Mesenchymal Transition

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
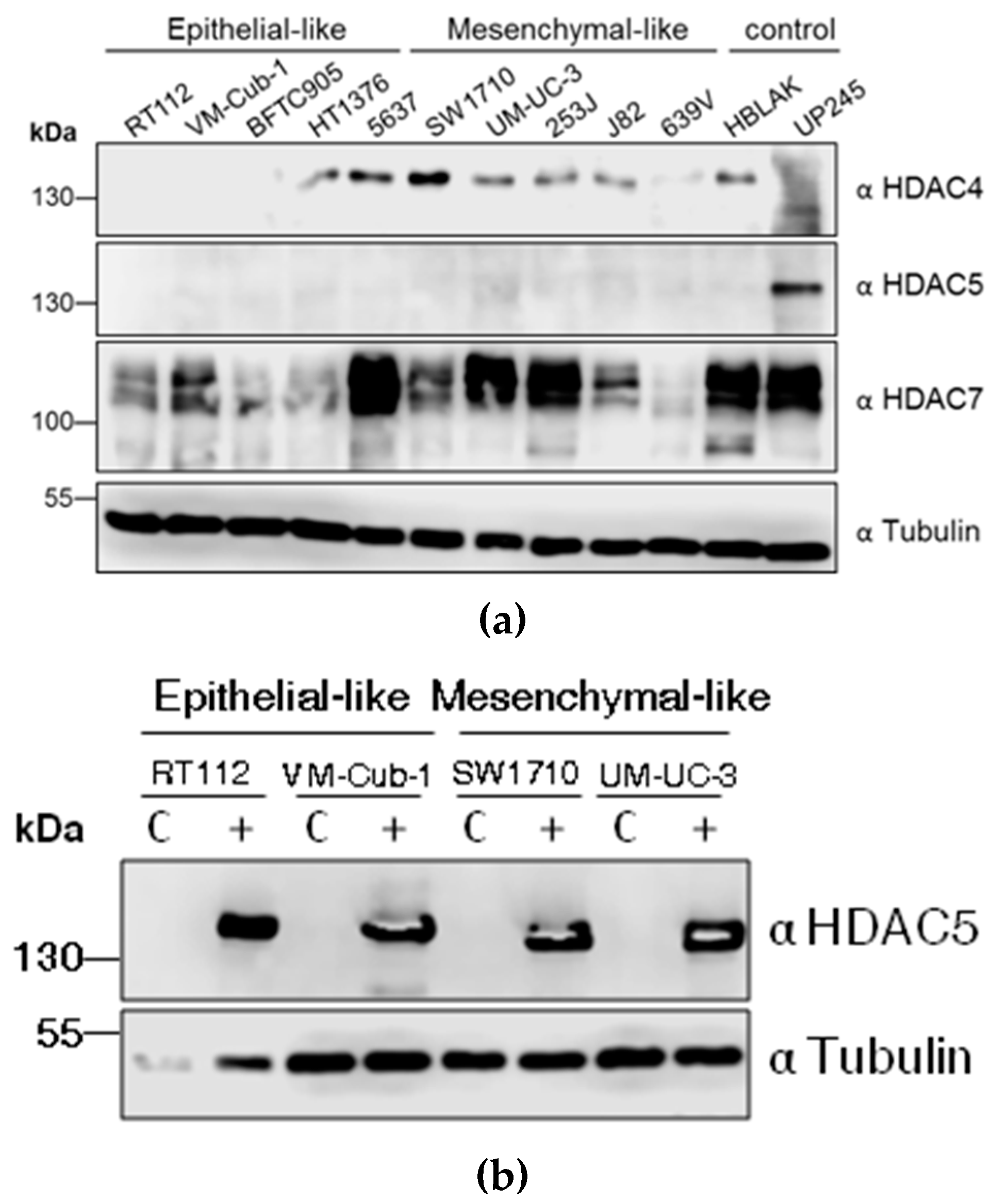
2.1. Consistent Downregulation of HDAC5 in Urothelial Carcinoma Cell Lines
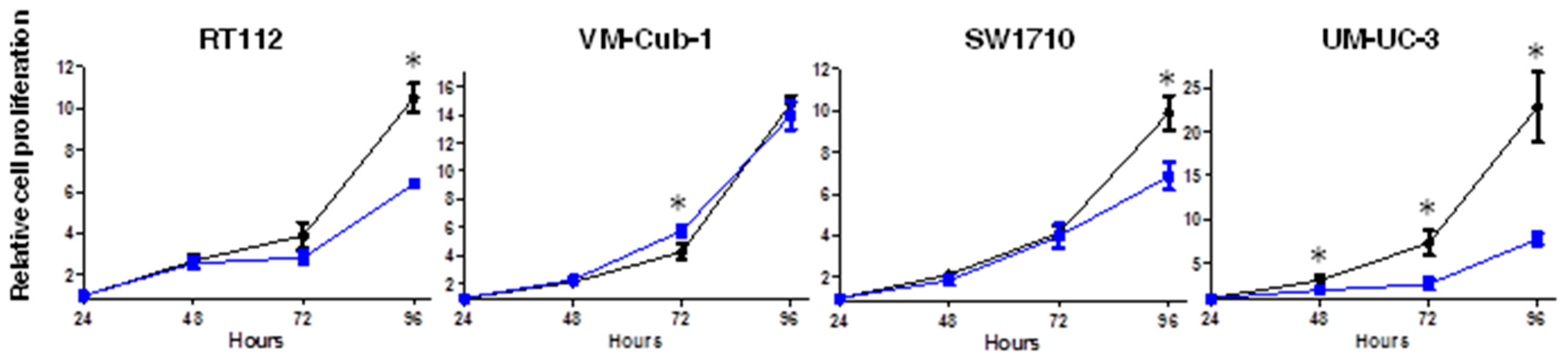
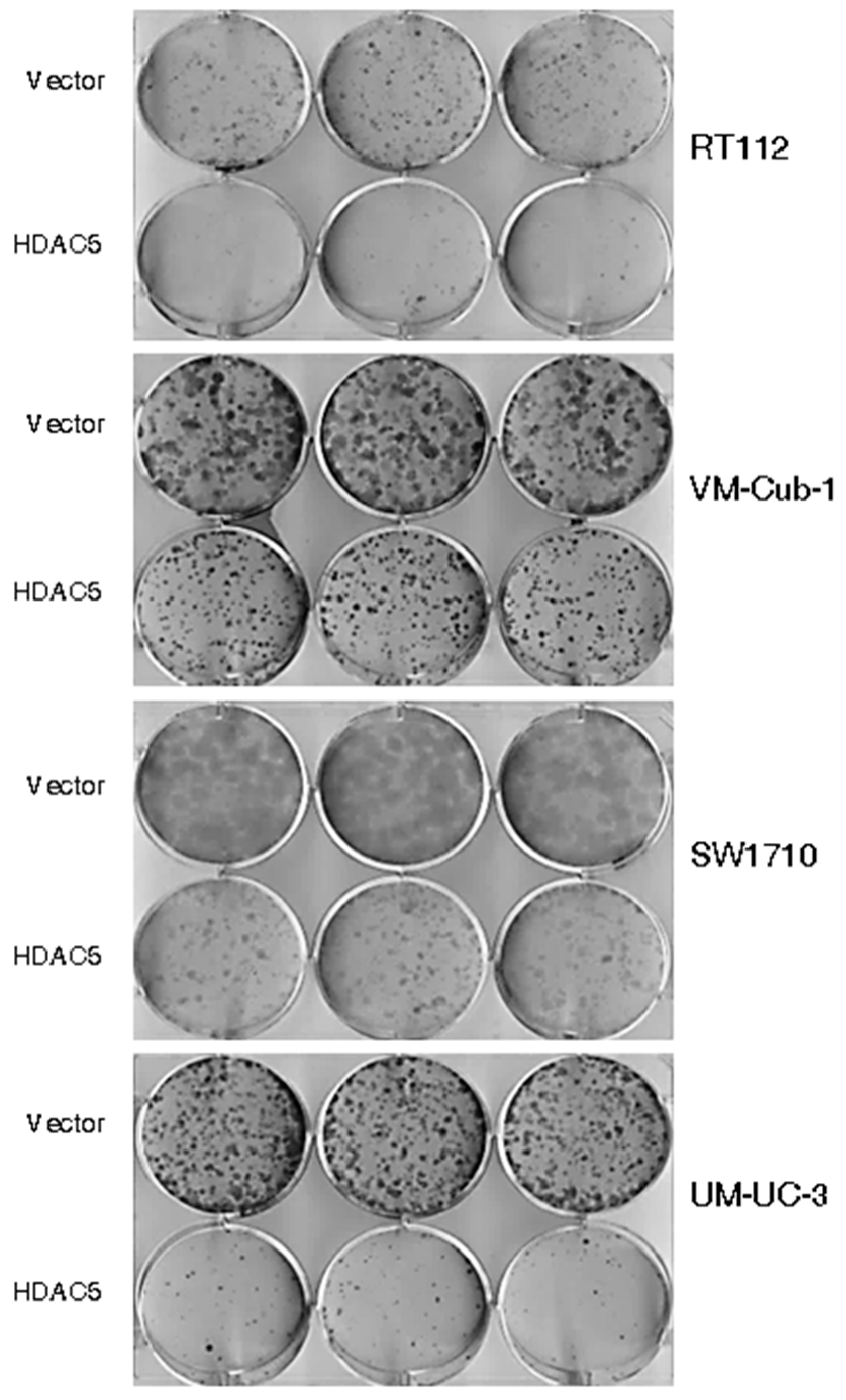
2.2. HDAC5 Expression Impairs Long-Term Proliferation and Clonogenic Potency of UCCs
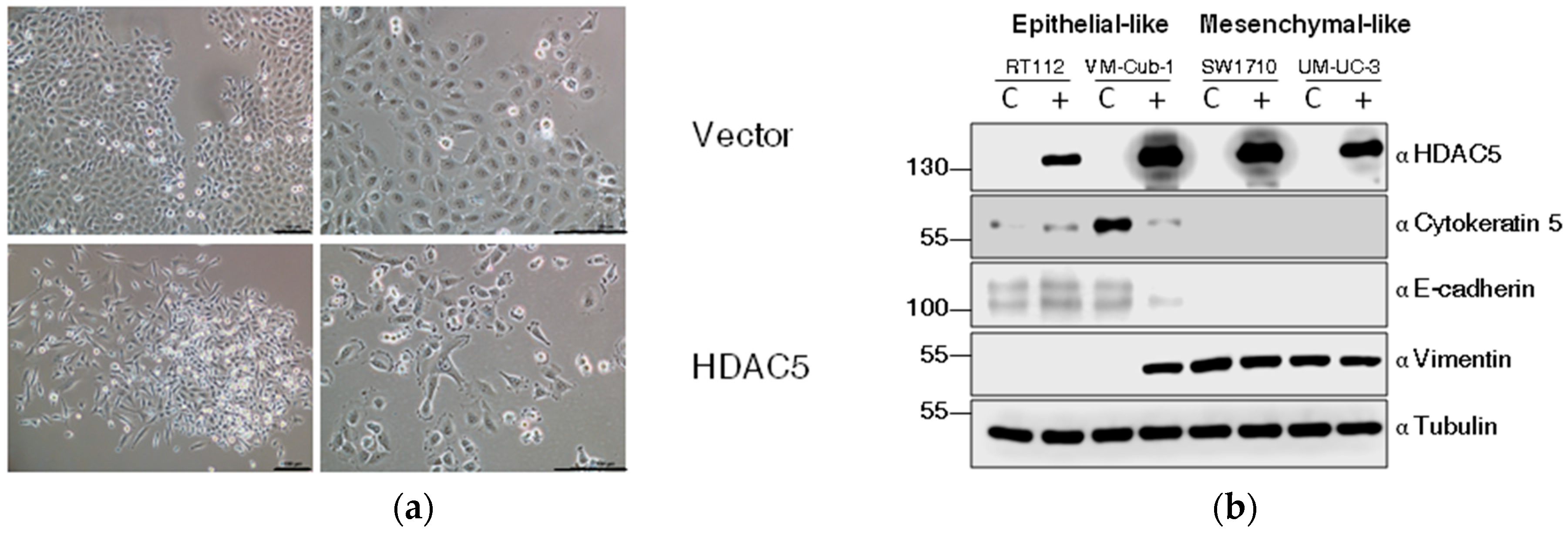
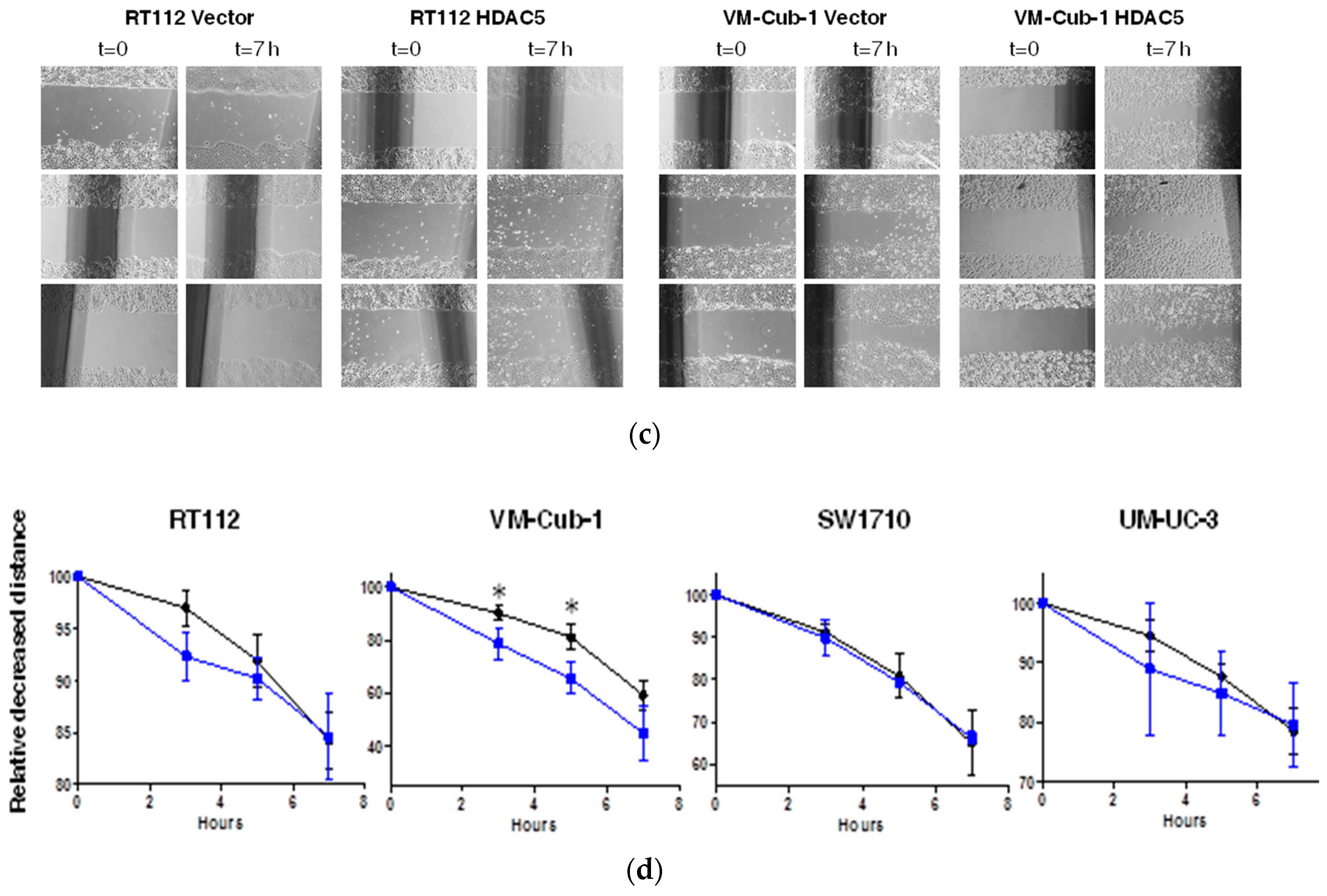
2.3. HDAC5 Induces an Epithelial-Mesenchymal Transition in VM-Cub-1 Cells
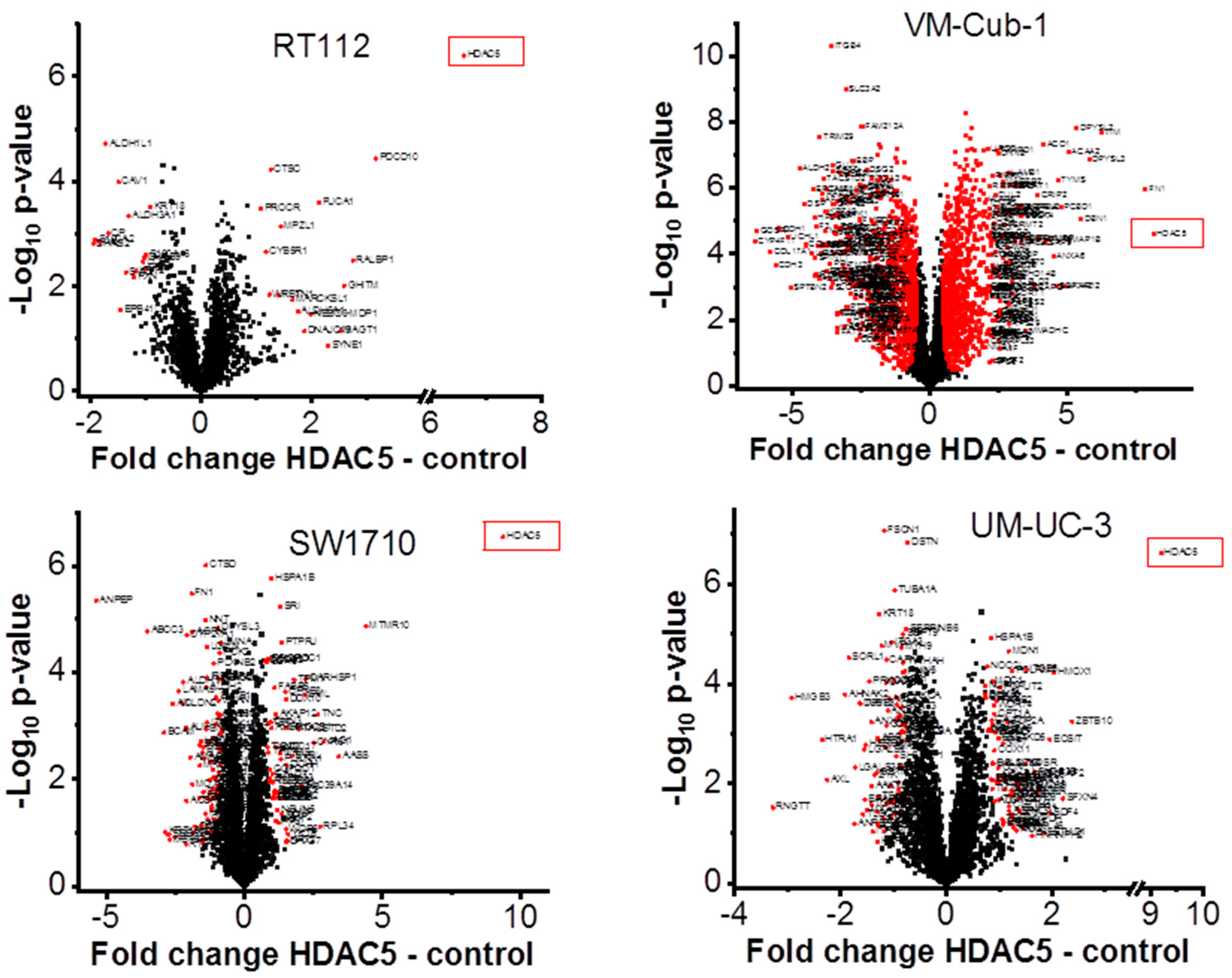
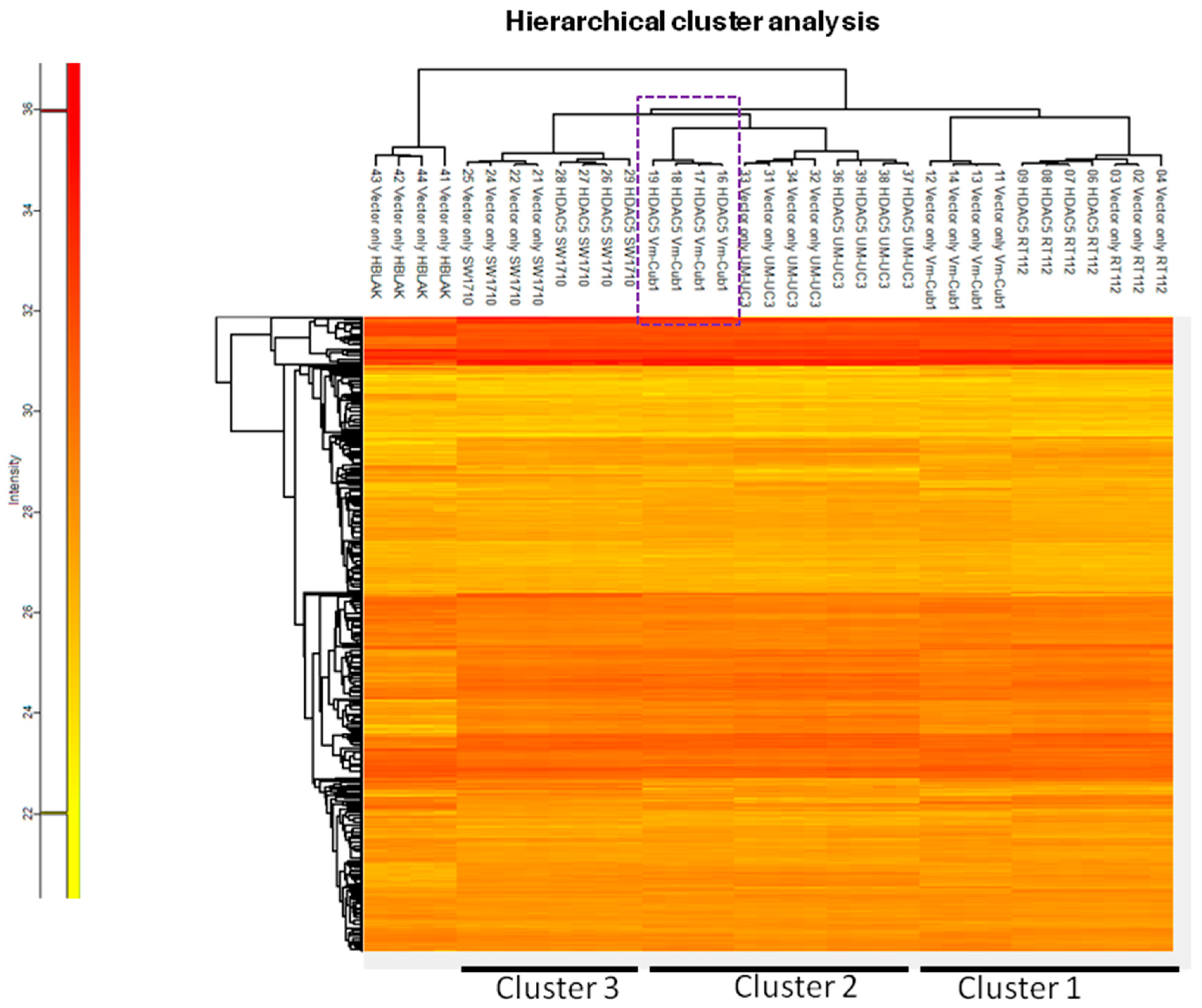
2.4. The Proteome of VM-Cub-1 Cells is Profoundly Altered by HDAC5
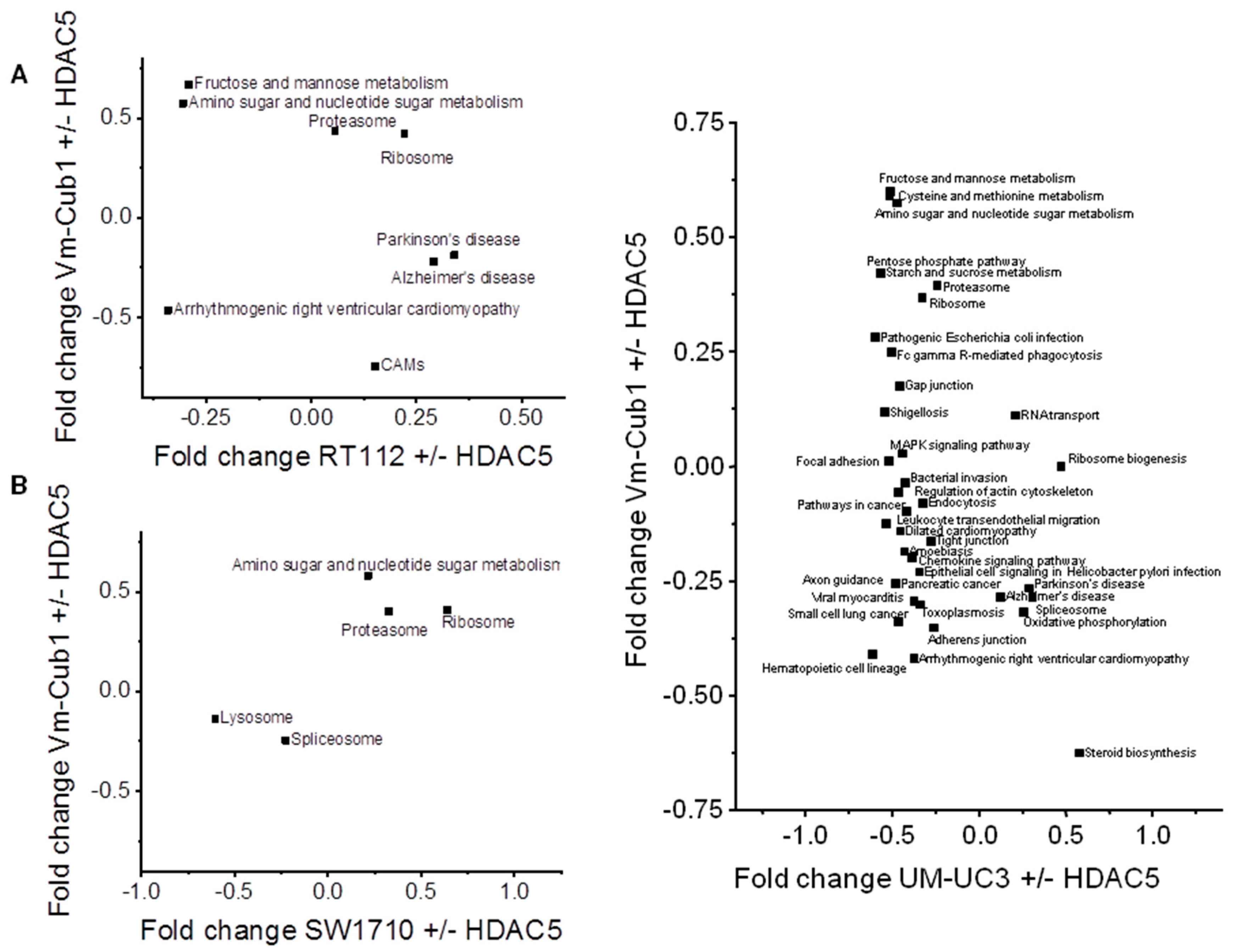
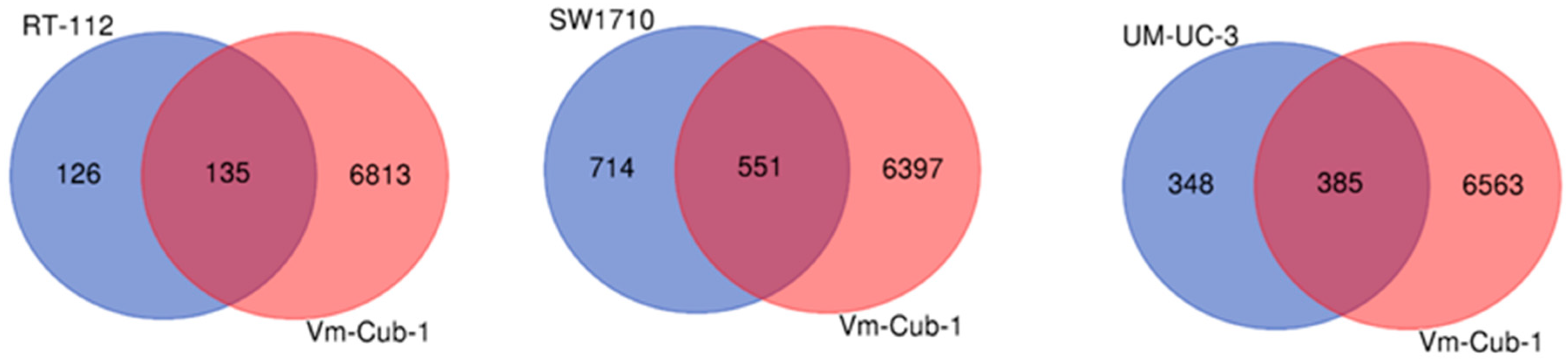
2.5. Transcriptome Changes by HDAC5 Are Most Extensive in VM-Cub-1 and Hint at an Involvement of TGFβ
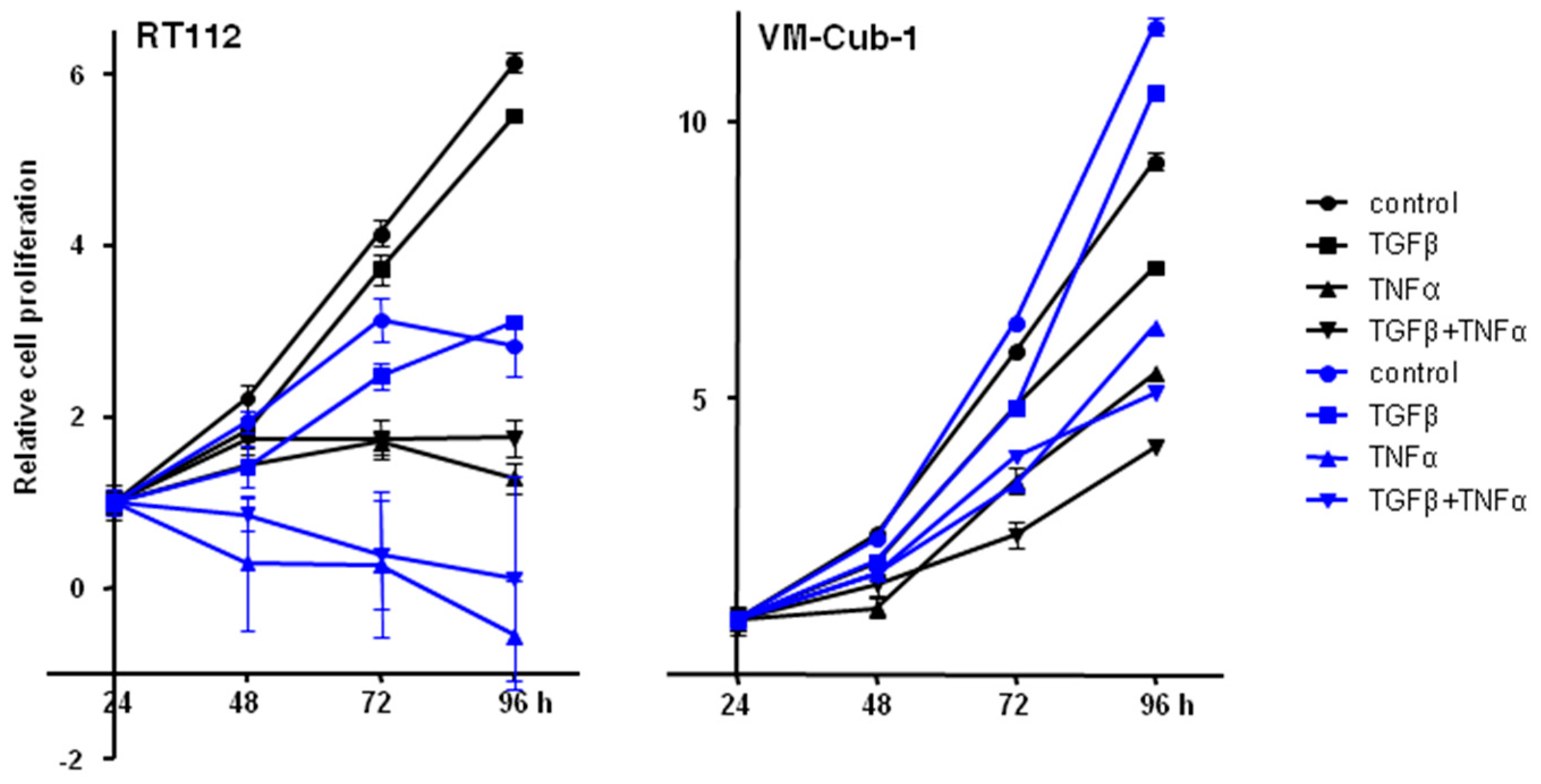
2.6. Effects of Added TGFβ and TNFα on Epithelial UC Cell Lines Are Not Influenced by HDAC5
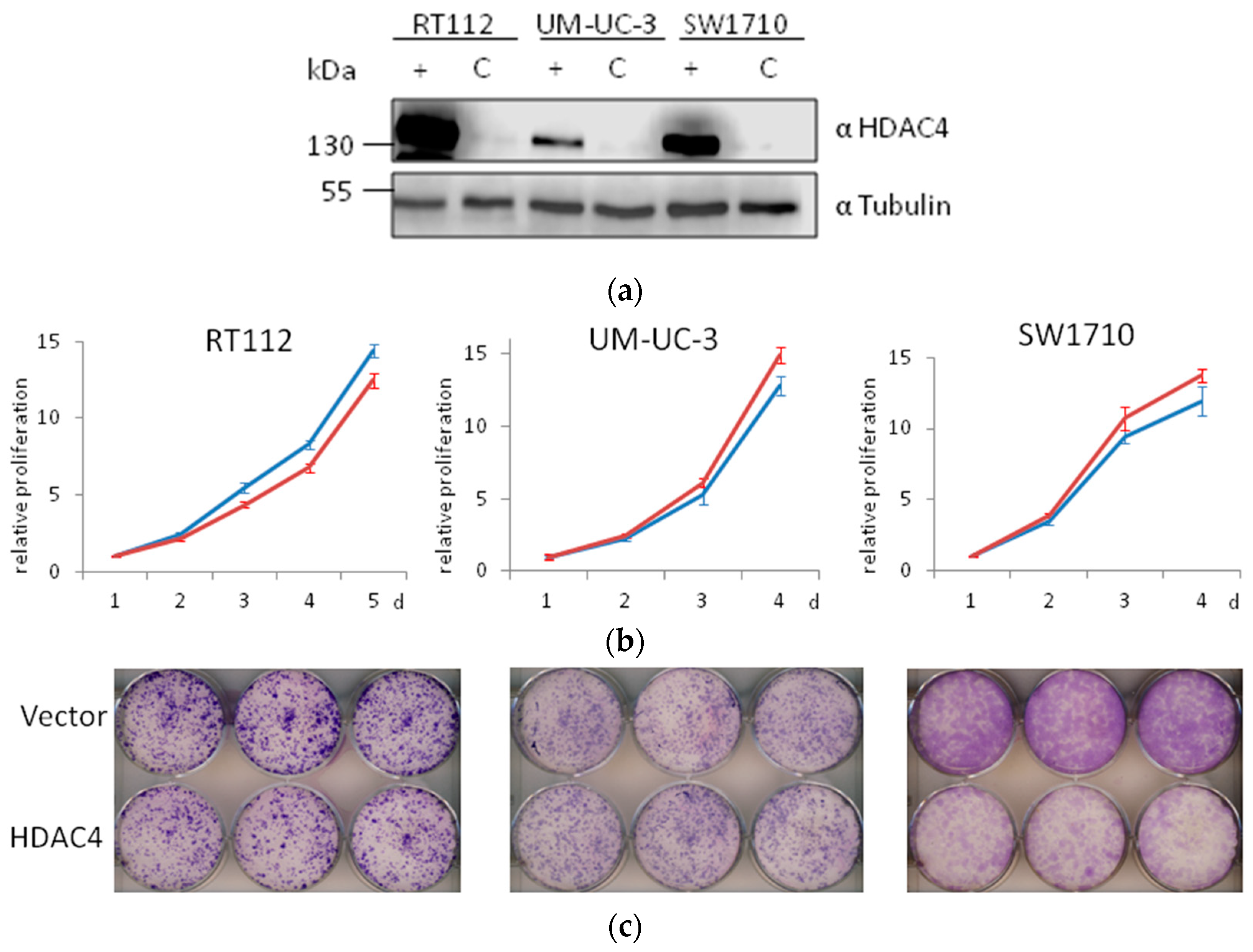
2.7. Weak Effects of HDAC4 Overexpression on UC Cell Lines
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Generation of Stably Eexpressing HDAC5 and Vector Control UC Cell Lines
4.3. Clone Formation Assays
4.4. Cell Proliferation Analysis
4.5. Migration Assay
4.6. Counting of Viable Cells by Flow Cytometry
4.7. Immunoblot Analysis
4.8. RNA Isolation, Sample Preparation, and High Throughput mRNA Sequencing
4.9. Proteome Analysis by Label-Free Quantification Based Mass Spectrometry
4.10. RNA-Seq and Data Analysis
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BCA | Bicinchoninic acid |
| DMEM | Dulbecco’s modified Eagle’s medium |
| EMT | Epithelial-mesenchymal transition |
| FDR | False discovery rate |
| HDAC | Histone deacetylase |
| IPA | Ingenuity Pathway analysis |
| SD | Standard deviation |
| TGF | Transforming growth factor |
| TNF | Tumor necrosis factor |
| UCC(s) | Urothelial carcinoma cell line(s) |
References
- Fischle, W.; Emiliani, S.; Hendzel, M.J.; Nagase, T.; Nomura, N.; Voelter, W.; Verdin, E. A new family of human histone deacetylases related to Saccharomyces cerevisiae HD. A1p. J. Biol. Chem. 1999, 274, 11713–11720. [Google Scholar] [CrossRef]
- Haberland, M.; Montgomery, R.L.; Olson, E.N. The many roles of histone deacetylases in development and physiology: implications for disease and therapy. Nat. Rev. Genet. 2009, 10, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Delcuve, G.P.; Khan, D.H.; Davie, J.R. Roles of histone deacetylases in epigenetic regulation: emerging paradigms from studies with inhibitors. Clin. Epigenetics 2012, 4, E5. [Google Scholar] [CrossRef] [PubMed]
- Di Giorgio, E.; Brancolini, C. Regulation of class IIa HDAC activities: it is not only matter of subcellular localization. Epigenomics. 2016, 8, 251–269. [Google Scholar] [CrossRef] [Green Version]
- Giannopoulou, A.F.; Velentzas, A.D.; Konstantakou, E.G.; Avgeris, M.; Katarachia, S.A.; Papandreou, N.C.; Kalavros, N.I.; Mpakou, V.E.; Iconomidou, V.; Anastasiadou, E.; et al. Revisiting histone deacetylases in human tumorigenesis: The paradigm of urothelial bladder cancer. Int. J. Mol. Sci. 2019, 20, 1291. [Google Scholar] [CrossRef] [PubMed]
- Wawruszak, A.; Kalafut, J.; Okon, E.; Czapinski, J.; Halasa, M.; Przybyszewska, A.; Miziak, P.; Okla, K.; Rivero-Muller, A.; Stepulak, A. Histone Deacetylase inhibitors and phenotypical transformation of cancer cells. Cancers 2019, 11, 148. [Google Scholar] [CrossRef]
- Parra, M. Class IIa HDACs - new insights into their functions in physiology and pathology. FEBS J. 2015, 282, 1736–1744. [Google Scholar] [CrossRef]
- Pinkerneil, M.; Hoffmann, M.J.; Deenen, R.; Köhrer, K.; Arent, T.; Schulz, W.A.; Niegisch, G. Inhibition of class I histone deacetylases 1 and 2 promotes urothelial carcinoma cell death by various mechanisms. Mol. Cancer Ther. 2016, 15, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Pinkerneil, M.; Hoffmann, M.J.; Schulz, W.A.; Niegisch, G. HDACs and HDAC inhibitors in urothelial carcinoma - Perspectives for an antineoplastic treatment. Curr. Med. Chem. 2017, 24, 4151–4165. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, El1. [Google Scholar] [CrossRef]
- Xu, X.S.; Wang, L.; Abrams, J.; Wang, G. Histone deacetylases (HDACs) in XPC gene silencing and bladder cancer. J. Hematol. Oncol. 2011, 4, E17. [Google Scholar] [CrossRef]
- Niegisch, G.; Knievel, J.; Koch, A.; Hader, C.; Fischer, U.; Albers, P.; Schulz, W.A. Changes in histone deacetylase (HDAC) expression patterns and activity of HDAC inhibitors in urothelial cancers. Urol. Oncol. 2013, 31, 1770–1779. [Google Scholar] [CrossRef]
- Kaletsch, A.; Pinkerneil, M.; Hoffmann, M.J.; Jaguva Vasudevan, A.A.; Wang, C.; Hansen, F.K.; Wiek, C.; Hanenberg, H.; Gertzen, C.; Gohlke, H.; et al. Effects of novel HDAC inhibitors on urothelial carcinoma cells. Clin. Epigenetics 2018, 10, E100. [Google Scholar] [CrossRef]
- Lehmann, L.H.; Jebessa, Z.H.; Kreusser, M.M.; Horsch, A.; He, T.; Kronlage, M.; Dewenter, M.; Sramek, V.; Oehl, U.; Krebs-Haupenthal, J.; et al. A proteolytic fragment of histone deacetylase 4 protects the heart from failure by regulating the hexosamine biosynthetic pathway. Nat. Med. 2018, 24, 62–72. [Google Scholar] [CrossRef]
- Mihaylova, M.M.; Vasquez, D.S.; Ravnskjaer, K.; Denechaud, P.D.; Yu, R.T.; Alvarez, J.G.; Downes, M.; Evans, R.M.; Montminy, M.; Shaw, R.J. Class IIa histone deacetylases are hormone-activated regulators of FOXO and mammalian glucose homeostasis. Cell 2011, 145, 607–621. [Google Scholar] [CrossRef]
- Cho, Y.; Cavalli, V. HDAC5 is a novel injury-regulated tubulin deacetylase controlling axon regeneration. EMBO J. 2012, 31, 3063–3078. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, D.A. Transforming growth factors—An overview. Biol. Cell. 1985, 53, 93–98. [Google Scholar] [CrossRef]
- Heubach, J.; Monsior, J.; Deenen, R.; Niegisch, G.; Szarvas, T.; Niedworok, C.; Schulz, W.A.; Hoffmann, M.J. The long noncoding RNA HOTAIR has tissue and cell type-dependent effects on HOX gene expression and phenotype of urothelial cancer cells. Mol. Cancer. 2015, 14, E108. [Google Scholar] [CrossRef]
- Earl, J.; Rico, D.; Carrillo-de-Santa-Pau, E.; Rodríguez-Santiago, B.; Méndez-Pertuz, M.; Auer, H.; Gómez, G.; Grossman, H.B.; Pisano, D.G.; Schulz, W.A. The UBC-40 Urothelial Bladder Cancer cell line index: a genomic resource for functional studies. BMC Genomics 2015, 16, E403. [Google Scholar]
- Gupta, S.; Hau, A.M.; Al-Ahmadie, H.A.; Harwalkar, J.; Shoskes, A.C.; Elson, P.; Beach, J.R.; Husse, G.S.; Schiemann, W.P.; Egelhoff, T.T.; et al. Transforming Growth Factor-β is an upstream regulator of Mammalian Target of Rapamycin Complex 2-dependent bladder cancer cell migration and invasion. Am. J. Pathol. 2016, 186, 1351–1360. [Google Scholar] [CrossRef]
- Liu, C.; Lv, D.; Li, M.; Zhang, X.; Sun, G.; Bai, Y.; Chang, D. Hypermethylation of miRNA-589 promoter leads to upregulation of HDAC5 which promotes malignancy in non-small cell lung cancer. Int. J. Oncol. 2017, 50, 2079–2090. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Piao, Z.H.; Jin, L.; Kim, J.H.; Kim, G.R.; Ryu, Y.; Lin, M.Q.; Kim, H.-S.; Kee, H.J.; Jeong, M.H. Piceatannol attenuates renal fibrosis induced by unilateral ureteral obstruction via downregulation of Histone Deacetylase 4/5 or p38-MAPK signaling. PLoS ONE 2016, 11, e0167340. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-β: the master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef]
- Lv, W.; Booz, G.W.; Wang, Y.; Fan, F.; Roman, R.J. Inflammation and renal fibrosis: Recent developments on key signaling molecules as potential therapeutic targets. Eur. J. Pharmacol. 2018, 820, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.J.; Koutsogiannouli, E.; Skowron, M.A.; Pinkerneil, M.; Niegisch, G.; Brandt, A.; Stepanow, S.; Rieder, H.; Schulz, W.A. The new immortalized uroepithelial cell line HBLAK contains defined genetic aberrations typical of early stage urothelial tumors. Bladder Cancer 2016, 2, 449–463. [Google Scholar] [CrossRef]
- Wiek, C.; Schmidt, E.M.; Roellecke, K.; Freund, M.; Nakano, M.; Kelly, E.J.; Kaisers, W.; Yarov-Yarovoy, V.; Kramm, C.M.; Rettie, A.E.; et al. Identification of amino acid determinants in CYP4B1 for optimal catalytic processing of 4-ipomeanol. Biochem J. 2015, 465, 103–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freshney, R.I. Culture of Animal Cells; Wiley-Liss: New York, NY, USA, 2000; pp. 200–202. [Google Scholar]
- Jaguva Vasudevan, A.A.; Kreimer, U.; Schulz, W.A.; Krikoni, A.; Schumann, G.G.; Häussinger, D.; Munk, C.; Goering, W. APOBEC3B activity is prevalent in urothelial carcinoma cells and only slightly affected by LINE-1 expression. Front. Microbiol. 2018, 9, E2088. [Google Scholar] [CrossRef]
- Grube, L.; Dellen, R.; Kruse, F.; Schwender, H.; Stühler, K.; Poschmann, G. Mining the secretome of C2C12 muscle cells: Data dependent experimental approach to analyze protein secretion using label-free quantification and peptide based analysis. J. Proteome Res. 2018, 17, 879–890. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | No. Genes FDR 1: p ≤ 0.05 | No. Genes Fold Change ≥2 | No. Genes Bonferroni: p ≤ 0.05 | No. Genes Fold Change ≥2 |
|---|---|---|---|---|
| RT112 Vector vs. HDAC5 | 3278 | 510 173 up 337 down | 1245 | 261 69 up 192 down |
| VM-Cub-1 Vector vs. HDAC5 | 16,883 | 9387 3331 up 6056 down | 12,074 | 6948 2645 up 4303 down |
| SW1710 Vector vs. HDAC5 | 10,941 | 2381 927 up 1454 down | 5949 | 1265 516 up 749 down |
| UM-UC-3 Vector vs. HDAC5 | 8318 | 1379 542 up 837 down | 4062 | 733 233 up 500 down |
| Rank | RT112 | VM-Cub-1 | SW1710 | UM-UC-3 |
|---|---|---|---|---|
| 1 | LPS/IL-1 Mediated Inhibition of RXR Function (9.5% 21/221) 1 | Axonal Guidance Signaling (46.9% 212/452) | Axonal Guidance Signaling (18.1% 82/452) | Hepatic Fibrosis/Hepatic Stellate Cell Activation (16.9% 31/183) |
| 2 | Granulocyte Adhesion and Diapedesis (10.1% 18/179) | Hepatic Fibrosis/Hepatic Stellate Cell Activation (55.2% 101/183) | LPS/IL-1 Mediated Inhibition of RXR Function (18.1% 40/221) | Axonal Guidance Signaling (10.8% 49/452) |
| 3 | Agranulocyte Adhesion and Diapedesis (9.4% 18/191) | Role of Macrophages, Fibroblasts and Endothelial Cells in Rheumatoid Arthritis (48.2% 150/311) | Coagulation System (34.3% 12/35) | Cellular Effects of Sildenafil (Viagra) (15.3% 20/131) |
| 4 | Xenobiotic Metabolism Signaling (7.3% 21/287) | Colorectal Cancer Metastasis Signaling (49.6% 123/248) | LXR/RXR Activation (20.7% 25/121) | Osteoarthritis Pathway (12.9% 27/210) |
| 5 | Inhibition of Matrix Metalloproteases (17.9% 7/39) | IL-8 Signaling (51.8% 102/197) | Hepatic Fibrosis/Hepatic Stellate Cell Activation (18.0% 33/183) | Coagulation System (25.7% 9/35) |
| Rank | RT112 | VM-Cub-1 | SW1710 | UM-UC-3 |
|---|---|---|---|---|
| 1 | TNF (Inhibited) | TNF (Inhibited) | TNF (Activated) | TNF (Inhibited) |
| 2 | SMARCA4 (not given) | estrogen receptor (Inhibited) | dexamethasone (not given) | Lipopolysaccharide (Inhibited) |
| 3 | Tretinoin (Inhibited) | TGFB1 (not given) | TGFB1 (Activated) | TGFB1 (not given) |
| 4 | Lipopolysaccharide (Inhibited) | SMARCA4 (Inhibited) | SMARCA4 (not given) | IFNG (Inhibited) |
| 5 | beta-estradiol (not given) | beta-estradiol (not given) | IFNG (not given) | IL1B (Inhibited) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaguva Vasudevan, A.A.; Hoffmann, M.J.; Beck, M.L.C.; Poschmann, G.; Petzsch, P.; Wiek, C.; Stühler, K.; Köhrer, K.; Schulz, W.A.; Niegisch, G. HDAC5 Expression in Urothelial Carcinoma Cell Lines Inhibits Long-Term Proliferation but Can Promote Epithelial-to-Mesenchymal Transition. Int. J. Mol. Sci. 2019, 20, 2135. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092135
Jaguva Vasudevan AA, Hoffmann MJ, Beck MLC, Poschmann G, Petzsch P, Wiek C, Stühler K, Köhrer K, Schulz WA, Niegisch G. HDAC5 Expression in Urothelial Carcinoma Cell Lines Inhibits Long-Term Proliferation but Can Promote Epithelial-to-Mesenchymal Transition. International Journal of Molecular Sciences. 2019; 20(9):2135. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092135
Chicago/Turabian StyleJaguva Vasudevan, Ananda Ayyappan, Michèle J. Hoffmann, Michael L. C. Beck, Gereon Poschmann, Patrick Petzsch, Constanze Wiek, Kai Stühler, Karl Köhrer, Wolfgang A. Schulz, and Günter Niegisch. 2019. "HDAC5 Expression in Urothelial Carcinoma Cell Lines Inhibits Long-Term Proliferation but Can Promote Epithelial-to-Mesenchymal Transition" International Journal of Molecular Sciences 20, no. 9: 2135. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092135