Characterizing a Halo-Tolerant GH10 Xylanase from Roseithermus sacchariphilus Strain RA and Its CBM-Truncated Variant


 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
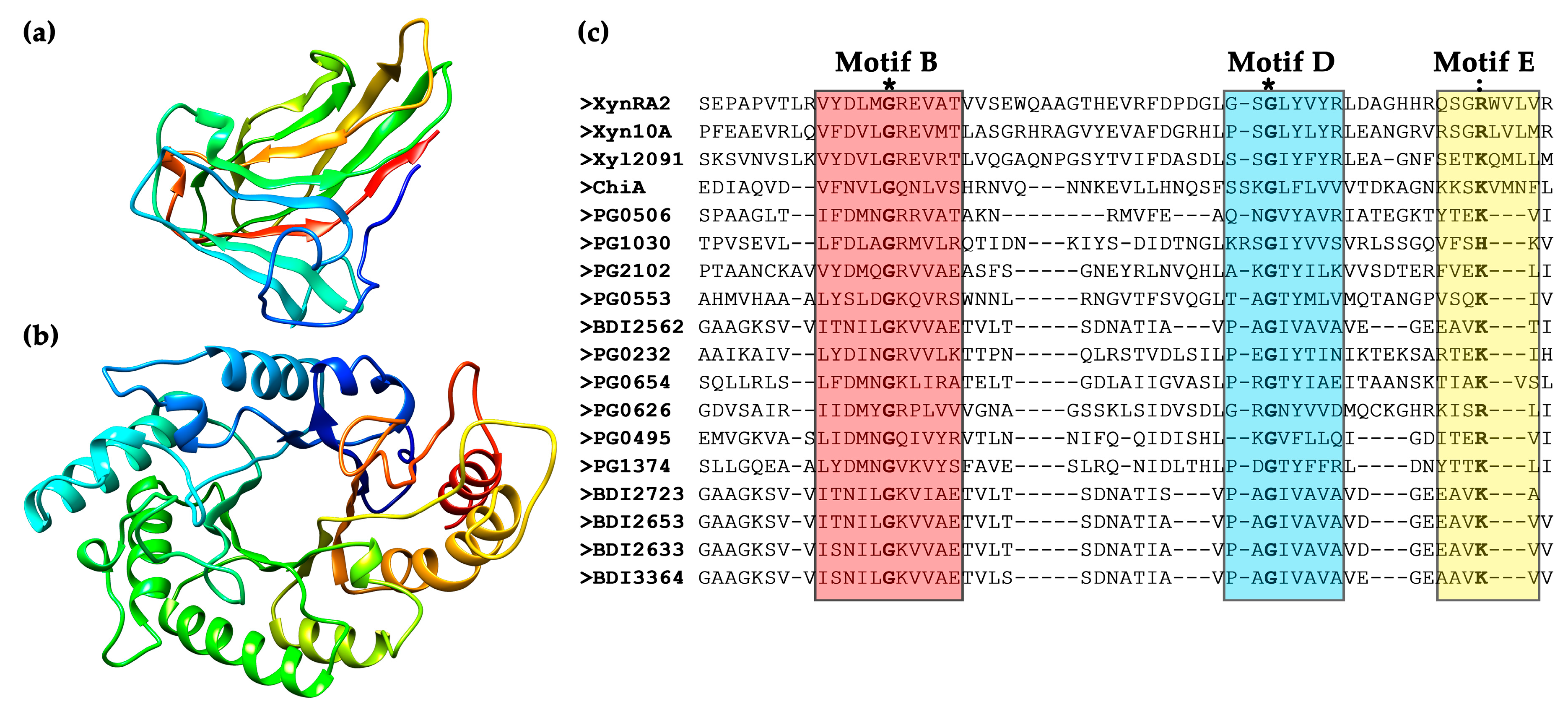
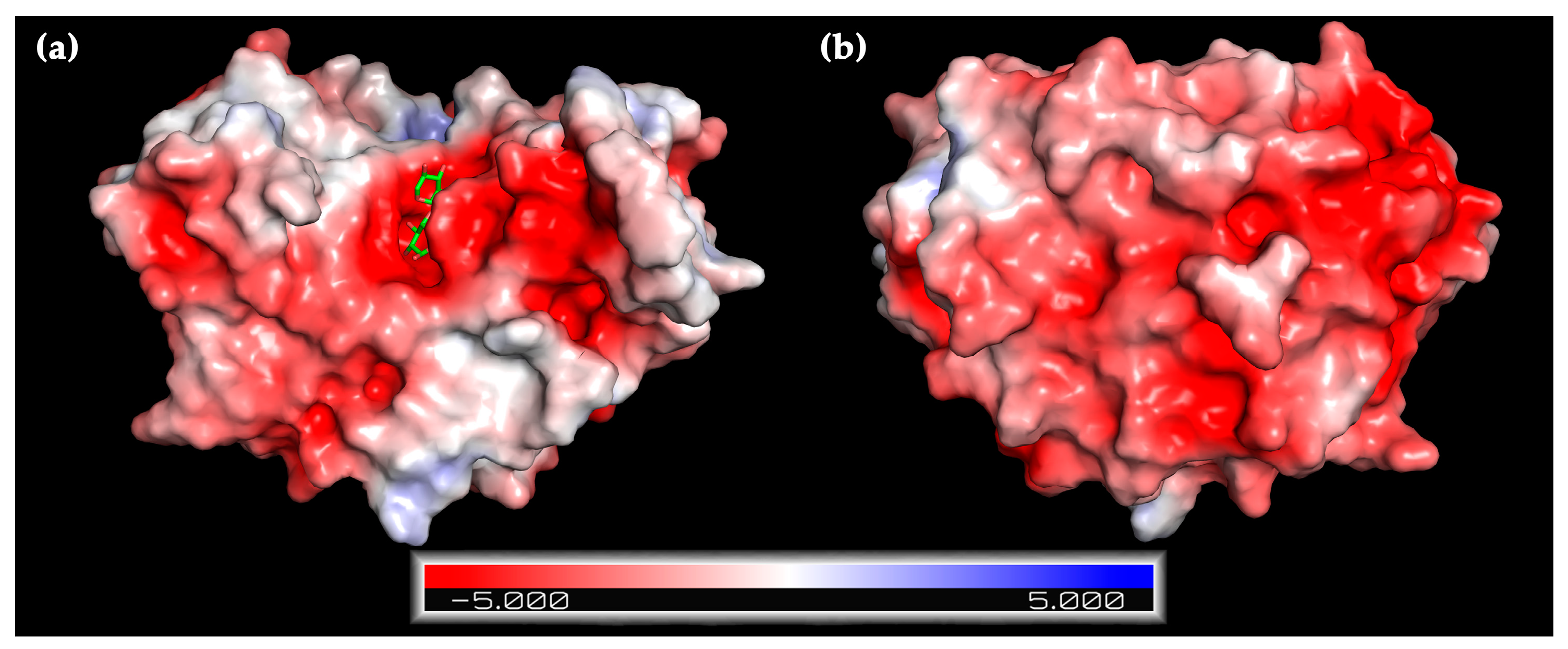
2.1. Bioinformatic Analysis
2.2. Expression of Recombinant XynRA2 and XynRA2ΔCBM
2.3. Biochemical Characterization of XynRA2 and XynRA2ΔCBM
2.3.1. Effect of pH and Temperature
2.3.2. NaCl Tolerance
2.3.3. Enzyme Kinetics
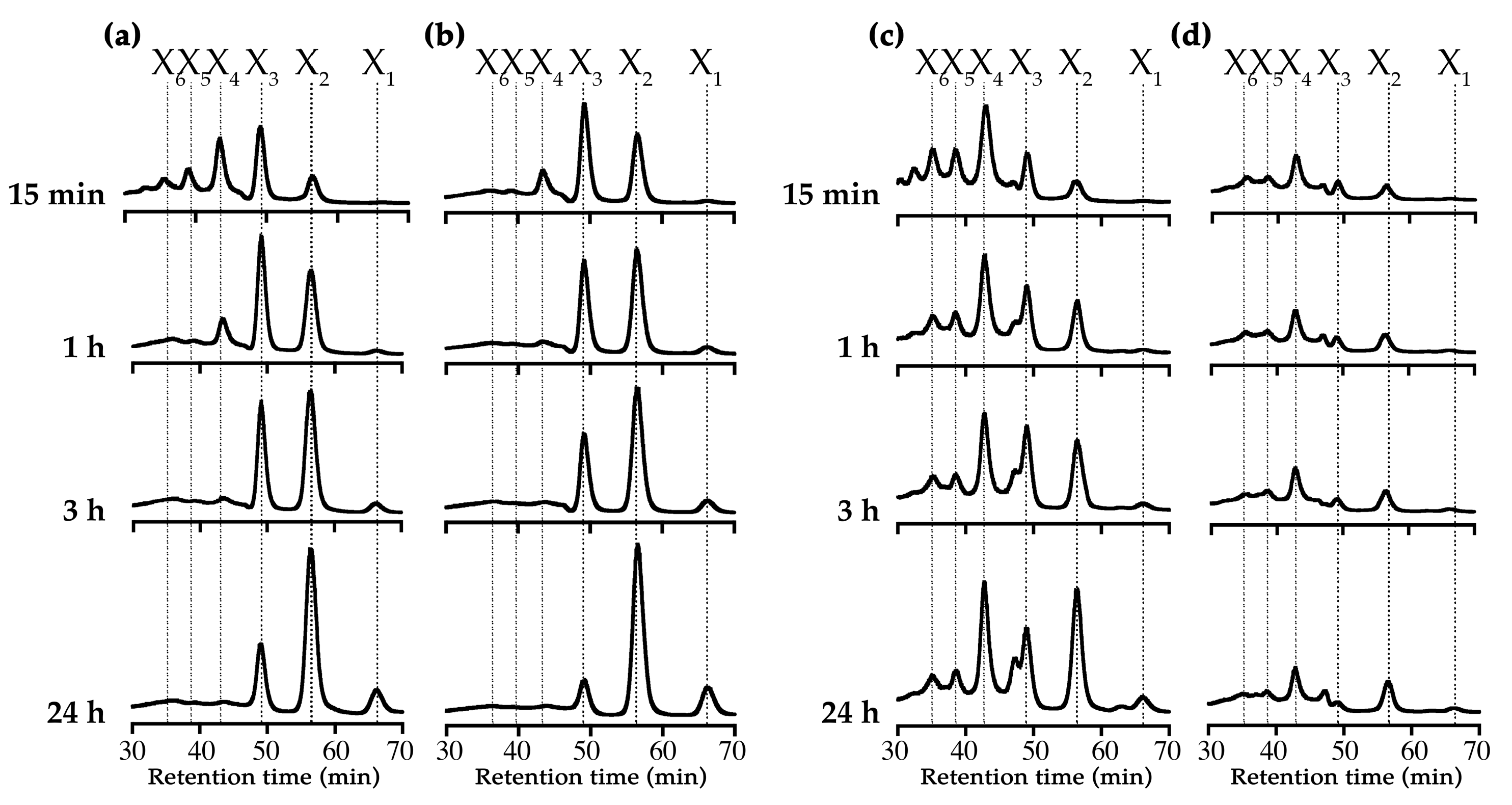
2.3.4. Substrate and Product Specificities
3. Materials and Methods
3.1. Sequence Analysis
3.2. Cloning of Xylanases
3.3. Expression and Purification of Xylanases
3.4. Xylanase Assay
3.5. Biochemical Characterization of XynRA2 and XynRA2ΔCBM
3.6. Analysis of Substrate Specificity and Hydrolysis Products
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANI | average nucleotide identity |
| BCA | bicinchoninic acid |
| BSA | bovine serum albumin |
| BLAST | Basic Local Alignment Search Tool |
| CAZy | carbohydrate active enzymes database |
| CBM | carbohydrate-binding module |
| CMC | carboxymethylcellulose |
| CTD | C-terminal domain |
| DNS | dinitrosalicylic acid |
| ELSD | evaporative light scattering detector |
| GH | glycoside hydrolase |
| HPLC | high-performance liquid chromatography |
| IPTG | isopropyl-β-d-1-thiogalactopyranoside |
| MEGA 7 | Molecular Evolutionary Genetics Analysis version 7 |
| MWCO | molecular weight cut-off |
| NCBI | National Center for Biotechnology Information |
| Ni-NTA | nickel-nitrilotriacetic acid |
| PorSS | Por secretion system |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| TIM | triosephosphate isomerase |
| T9SS | type IX secretion system |
| XOs | xylo-oligosaccharides |
| X1 | xylose |
| X2 | xylobiose |
| X3 | xylotriose |
| X4 | xylotetraose |
| X5 | xylopentaose |
| X6 | xylohexaose |
References
- Collins, T.; Gerday, C.; Feller, G. Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol. Rev. 2005, 29, 3–23. [Google Scholar] [CrossRef] [Green Version]
- Moreira, L.R.S. Insights into the mechanism of enzymatic hydrolysis of xylan. Appl. Microbiol. Biotechnol. 2016, 100, 5205–5214. [Google Scholar] [CrossRef] [PubMed]
- Terrapon, N.; Lombard, V.; Drula, E.; Coutinho, P.M.; Henrissat, B. The CAZy Database/the Carbohydrate-Active Enzyme (CAZy) Database: Principles and Usage Guidelines. In A Practical Guide to Using Glycomics Databases; Aoki-Kinoshita, K.F., Ed.; Springer: Tokyo, Japan, 2017; pp. 117–131. [Google Scholar]
- Nguyen, S.T.; Freund, H.L.; Kasanjian, J.; Berlemont, R. Function, distribution, and annotation of characterized cellulases, xylanases, and chitinases from CAZy. Appl. Microbiol. Biotechnol. 2018, 102, 1629–1637. [Google Scholar] [CrossRef]
- Chakdar, H.; Kumar, M.; Pandiyan, K.; Singh, A.; Nanjappan, K.; Kashyap, P.L.; Srivastava, A.K. Bacterial xylanases: Biology to biotechnology. 3 Biotech 2016, 6, 150. [Google Scholar] [CrossRef] [PubMed]
- Juturu, V.; Wu, J.C. Microbial xylanases: Engineering, production and industrial applications. Biotechnol. Adv. 2012, 30, 1219–1227. [Google Scholar] [CrossRef]
- Kumar, V.; Marin-Navarro, J.; Shukla, P. Thermostable microbial xylanases for pulp and paper industries: Trends, applications and further perspectives. World J. Microb. Biotechnol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Luo, H.; Wang, Y.; Li, J.; Yang, J.; Yang, Y.; Huang, H.; Fan, Y.; Yao, B. Cloning, expression and characterization of a novel acidic xylanase, XYL11B, from the acidophilic fungus Bispora sp. MEY-1. Enzyme Microb. Technol. 2009, 45, 126–133. [Google Scholar] [CrossRef]
- Kumar, B.K.; Balakrishnan, H.; Rele, M. Compatibility of alkaline xylanases from an alkaliphilic Bacillus NCL (87-6-10) with commercial detergents and proteases. J. Ind. Microbiol. Biot. 2004, 31, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Dornez, E.; Verjans, P.; Arnaut, F.; Delcour, J.A.; Courtin, C.M. Use of psychrophilic xylanases provides insight into the xylanase functionality in bread making. J. Agr. Food Chem. 2011, 59, 9553–9562. [Google Scholar] [CrossRef]
- Nagar, S.; Mittal, A.; Gupta, V.K. Enzymatic clarification of fruit juices (apple, pineapple, and tomato) using purified Bacillus pumilus SV-85S xylanase. Biotechnol. Bioproc. Eng. 2012, 17, 1165–1175. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, X.; Bai, F. Production of xylanase by an alkaline-tolerant marine-derived Streptomyces viridochromogenes strain and improvement by ribosome engineering. Appl. Microbiol. Biotechnol. 2013, 97, 4361–4368. [Google Scholar] [CrossRef]
- Talamantes, D.; Biabini, N.; Dang, H.; Abdoun, K.; Berlemont, R. Natural diversity of cellulases, xylanases, and chitinases in bacteria. Biotechnol. Biofuels 2016, 9, 133. [Google Scholar] [CrossRef]
- Chawachart, N.; Anbarasan, S.; Turunen, S.; Li, H.; Khanongnuch, C.; Hummel, M.; Sixta, H.; Granström, T.; Lumyong, S.; Turunen, O. Thermal behaviour and tolerance to ionic liquid [emim]OAc in GH10 xylanase from Thermoascus aurantiacus SL16W. Extremophiles 2014, 18, 1023–1034. [Google Scholar] [CrossRef]
- Evangelista, D.E.; Kadowaki, M.A.S.; Mello, B.L.; Polikarpov, I. Biochemical and biophysical characterization of novel GH10 xylanase prospected from a sugar cane bagasse compost-derived microbial consortia. Int. J. Biol. Macromol. 2018, 109, 560–568. [Google Scholar] [CrossRef]
- Niderhaus, C.; Garrido, M.; Insani, M.; Campos, E.; Wirth, S. Heterologous production and characterization of a thermostable GH10 family endo-xylanase from Pycnoporus sanguineus BAFC 2126. Process Biochem. 2018, 67, 92–98. [Google Scholar] [CrossRef]
- Sharma, K.; Antunes, I.L.; Rajulapati, V.; Goyal, A. Molecular characterization of a first endo-acting β-1,4-xylanase of family 10 glycoside hydrolase (PsGH10A) from Pseudopedobacter saltans comb. nov. Process Biochem. 2018, 70, 79–89. [Google Scholar] [CrossRef]
- Guillén, D.; Sánchez, S.; Rodríguez-Sanoja, R. Carbohydrate-binding domains: Multiplicity of biological roles. Appl. Microbiol. Biotechnol. 2010, 85, 1241–1249. [Google Scholar] [CrossRef]
- Varnai, A.; Mäkelä, M.R.; Djajadi, D.T.; Rahikainen, J.; Hatakka, A.; Viikari, L. Carbohydrate-binding modules of fungal cellulases: Occurrence in nature, function, and relevance in industrial biomass conversion. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 88, pp. 103–165. [Google Scholar]
- Goh, K.M.; Chan, K.-G.; Lim, S.W.; Liew, K.J.; Chan, C.S.; Shamsir, M.S.; Ee, R.; Adrian, T.-G.-S. Genome analysis of a new Rhodothermaceae strain isolated from a hot spring. Front. Microbiol. 2016, 7, 1109. [Google Scholar] [CrossRef]
- Liew, K.J.; Teo, S.C.; Shamsir, M.S.; Sani, R.K.; Chong, C.S.; Chan, K.-G.; Goh, K.M. Complete genome sequence of Rhodothermaceae bacterium RA with cellulolytic and xylanolytic activities. 3 Biotech 2018, 8, 376. [Google Scholar] [CrossRef]
- Park, M.-J.; Oh, J.H.; Yang, S.-H.; Kwon, K.K. Roseithermus sacchariphilus gen. nov., sp. nov. and proposal of Salisaetaceae fam. nov., representing new family in the order Rhodothermales. Int. J. Syst. Evol. Microbiol. 2019, 69, 1213–1219. [Google Scholar] [CrossRef]
- Cai, Z.-W.; Ge, H.-H.; Yi, Z.-W.; Zeng, R.-Y.; Zhang, G.-Y. Characterization of a novel psychrophilic and halophilic β-1,3-xylanase from deep-sea bacterium, Flammeovirga pacifica strain WPAGA1. Int. J. Biol. Macromol. 2018, 118, 2176–2184. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wu, J.; Yan, R.; Lin, J.; Ye, X. A novel multi-domain high molecular, salt-stable alkaline xylanase from Alkalibacterium sp. SL3. Front. Microbiol. 2017, 7, 2120. [Google Scholar] [CrossRef]
- Yadav, P.; Maharjan, J.; Korpole, S.; Prasad, G.S.; Sahni, G.; Bhattarai, T.; Sreerama, L. Production, purification, and characterization of thermostable alkaline xylanase from Anoxybacillus kamchatkensis NASTPD13. Front. Bioeng. Biotechnol. 2018, 6, 65. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.S.; Chan, K.-G.; Ee, R.; Hong, K.-W.; Urbieta, M.S.; Donati, E.R.; Shamsir, M.S.; Goh, K.M. Effects of physiochemical factors on prokaryotic biodiversity in Malaysian circumneutral hot springs. Front. Microbiol. 2017, 8, 1252. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, E.N.; Bartonek-Roxå, E.; Holst, O. Cloning and sequence of a thermostable multidomain xylanase from the bacterium Rhodothermus marinus. BBA - Gene Struct. Expr. 1997, 1353, 118–124. [Google Scholar] [CrossRef]
- Rakitin, A.L.; Ermakova, A.Y.; Ravin, N.V. Novel endoxylanases of the moderately thermophilic polysaccharide-degrading bacterium Melioribacter roseus. J. Microbiol. Biotechnol. 2015, 25, 1476–1484. [Google Scholar] [CrossRef]
- Beliën, T.; Van Campenhout, S.; Van Acker, M.; Volckaert, G. Cloning and characterization of two endoxylanases from the cereal phytopathogen Fusarium graminearum and their inhibition profile against endoxylanase inhibitors from wheat. Biochem. Biophys. Res. Commun. 2005, 327, 407–414. [Google Scholar] [CrossRef]
- La Grange, D.C.; Pretorius, I.S.; Van Zyl, W.H. Expression of a Trichoderma reesei beta-xylanase gene (XYN2) in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 1996, 62, 1036–1044. [Google Scholar]
- Khasin, A.; Alchanati, I.; Shoham, Y. Purification and characterization of a thermostable xylanase from Bacillus stearothermophilus T-6. Appl. Environ. Microbiol. 1993, 59, 1725–1730. [Google Scholar]
- Lee, Y.-E.; Lowe, S.; Henrissat, B.; Zeikus, J.G. Characterization of the active site and thermostability regions of endoxylanase from Thermoanaerobacterium saccharolyticum B6A-RI. J. Bacteriol. 1993, 175, 5890–5898. [Google Scholar] [CrossRef]
- Lasica, A.M.; Ksiazek, M.; Madej, M.; Potempa, J. The type IX secretion system (T9SS): Highlights and recent insights into its structure and function. Front. Cell. Infect. Microbiol. 2017, 7, 215. [Google Scholar] [CrossRef]
- Sato, K.; Yukitake, H.; Narita, Y.; Shoji, M.; Naito, M.; Nakayama, K. Identification of Porphyromonas gingivalis proteins secreted by the Por secretion system. FEMS Microbiol. Lett. 2013, 338, 68–76. [Google Scholar] [CrossRef]
- Karlsson, E.N.; Hachem, M.A.; Ramchuran, S.; Costa, H.; Holst, O.; Svenningsen, Å.F.; Hreggvidsson, G.O. The modular xylanase Xyn10A from Rhodothermus marinus is cell-attached, and its C-terminal domain has several putative homologues among cell-attached proteins within the phylum Bacteroidetes. FEMS Microbiol. Lett. 2004, 241, 233–242. [Google Scholar] [CrossRef]
- Kharade, S.S.; McBride, M.J. Flavobacterium johnsoniae chitinase ChiA is required for chitin utilization and is secreted by the type IX secretion system. J. Bacteriol. 2014, 196, 961–970. [Google Scholar] [CrossRef]
- Veith, P.D.; Nor Muhammad, N.A.; Dashper, S.G.; Likić, V.A.; Gorasia, D.G.; Chen, D.; Byrne, S.J.; Catmull, D.V.; Reynolds, E.C. Protein substrates of a novel secretion system are numerous in the Bacteroidetes phylum and have in common a cleavable C-terminal secretion signal, extensive post-translational modification, and cell-surface attachment. J. Proteome Res. 2013, 12, 4449–4461. [Google Scholar] [CrossRef]
- Hachem, M.A.; Karlsson, E.N.; Bartonek-Roxå, E.; Raghothama, S.; Simpson, P.J.; Gilbert, H.J.; Williamson, M.P.; Holst, O. Carbohydrate-binding modules from a thermostable Rhodothermus marinus xylanase: Cloning, expression and binding studies. Biochem. J. 2000, 345, 53–60. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, L.; Li, W.; Guo, W.; Zheng, H.; Wang, J.; Lu, F. Studies on properties of the xylan-binding domain and linker sequence of xylanase XynG1-1 from Paenibacillus campinasensis G1-1. J. Ind. Microbiol. Biot. 2015, 42, 1591–1599. [Google Scholar] [CrossRef]
- Li, N.; Shi, P.; Yang, P.; Wang, Y.; Luo, H.; Bai, Y.; Zhou, Z.; Yao, B. A xylanase with high pH stability from Streptomyces sp. S27 and its carbohydrate-binding module with/without linker-region-truncated versions. Appl. Microbiol. Biotechnol. 2009, 83, 99–107. [Google Scholar] [CrossRef]
- Zhang, M.; Chekan, J.R.; Dodd, D.; Hong, P.-Y.; Radlinski, L.; Revindran, V.; Nair, S.K.; Mackie, R.I.; Cann, I. Xylan utilization in human gut commensal bacteria is orchestrated by unique modular organization of polysaccharide-degrading enzymes. Proc. Natl. Acad. Sci. USA 2014, 111, E3708–E3717. [Google Scholar] [CrossRef]
- Karlsson, E.N.; Bartonek-Roxå, E.; Holst, O. Evidence for substrate binding of a recombinant thermostable xylanase originating from Rhodothermus marinus. FEMS Microbiol. Lett. 1998, 168, 1–7. [Google Scholar] [CrossRef]
- Li, Q.; Sun, B.; Li, X.; Xiong, K.; Xu, Y.; Yang, R.; Hou, J.; Teng, C. Improvement of the catalytic characteristics of a salt-tolerant GH10 xylanase from Streptomyce rochei L10904. Int. J. Biol. Macromol. 2018, 107, 1447–1455. [Google Scholar] [CrossRef]
- Lim, H.K.; Lee, K.I.; Hwang, I.T. Identification of a novel cellulose-binding domain within the endo-β-1,4-xylanase KRICT PX-3 from Paenibacillus terrae HPL-003. Enzyme Microb. Technol. 2016, 93, 166–173. [Google Scholar]
- Karlsson, E.N.; Dahlberg, L.; Torto, N.; Gorton, L.; Holst, O. Enzymatic specificity and hydrolysis pattern of the catalytic domain of the xylanase Xyn1 from Rhodothermus marinus. J. Biotechnol. 1998, 60, 23–35. [Google Scholar] [CrossRef]
- Hachem, M.A.; Olsson, F.; Nordberg Karlsson, E. Probing the stability of the modular family 10 xylanase from Rhodothermus marinus. Extremophiles 2003, 7, 483–491. [Google Scholar] [CrossRef]
- Graziano, G.; Merlino, A. Molecular bases of protein halotolerance. BBA - Proteins Proteom. 2014, 1844, 850–858. [Google Scholar] [CrossRef]
- Fukuchi, S.; Yoshimune, K.; Wakayama, M.; Moriguchi, M.; Nishikawa, K. Unique amino acid composition of proteins in halophilic bacteria. J. Mol. Biol. 2003, 327, 347–357. [Google Scholar] [CrossRef]
- Liu, X.; Huang, Z.; Zhang, X.; Shao, Z.; Liu, Z. Cloning, expression and characterization of a novel cold-active and halophilic xylanase from Zunongwangia profunda. Extremophiles 2014, 18, 441–450. [Google Scholar] [CrossRef]
- Guo, B.; Chen, X.-L.; Sun, C.-Y.; Zhou, B.-C.; Zhang, Y.-Z. Gene cloning, expression and characterization of a new cold-active and salt-tolerant endo-β-1,4-xylanase from marine Glaciecola mesophila KMM 241. Appl. Microbiol. Biotechnol. 2009, 84, 1107–1115. [Google Scholar] [CrossRef]
- Huang, X.; Lin, J.; Ye, X.; Wang, G. Molecular characterization of a thermophilic and salt-and alkaline-tolerant xylanase from Planococcus sp. SL4, a strain isolated from the sediment of a soda lake. J. Microbiol. Biotechnol. 2015, 25, 662–671. [Google Scholar] [CrossRef]
- Xu, B.; Dai, L.; Li, J.; Deng, M.; Miao, H.; Zhou, J.; Mu, Y.; Wu, Q.; Tang, X.; Yang, Y. Molecular and biochemical characterization of a novel xylanase from Massilia sp. RBM26 isolated from the feces of Rhinopithecus bieti. J. Microbiol. Biotechnol. 2015, 26, 9–19. [Google Scholar] [CrossRef]
- Zhou, J.; Gao, Y.; Dong, Y.; Tang, X.; Li, J.; Xu, B.; Mu, Y.; Wu, Q.; Huang, Z. A novel xylanase with tolerance to ethanol, salt, protease, SDS, heat, and alkali from actinomycete Lechevalieria sp. HJ3. J. Ind. Microbiol. Biot. 2012, 39, 965–975. [Google Scholar] [CrossRef]
- Poosarla, V.G.; Chandra, T. Purification and characterization of novel halo-acid-alkali-thermo-stable xylanase from Gracilibacillus sp. TSCPVG. Appl. Biochem. Biotech. 2014, 173, 1375–1390. [Google Scholar] [CrossRef]
- Verjans, P.; Dornez, E.; Segers, M.; Van Campenhout, S.; Bernaerts, K.; Beliën, T.; Delcour, J.A.; Courtin, C.M. Truncated derivatives of a multidomain thermophilic glycosyl hydrolase family 10 xylanase from Thermotoga maritima reveal structure related activity profiles and substrate hydrolysis patterns. J. Biotechnol. 2010, 145, 160–167. [Google Scholar] [CrossRef]
- Sunna, A.; Gibbs, M.D.; Bergquist, P.L. A novel thermostable multidomain 1,4-β-xylanase from ‘Caldibacillus cellulovorans’ and effect of its xylan-binding domain on enzyme activity. Microbiol. 2000, 146, 2947–2955. [Google Scholar] [CrossRef]
- Ali, M.K.; Hayashi, H.; Karita, S.; Goto, M.; Kimura, T.; Sakka, K.; Ohmiya, K. Importance of the carbohydrate-binding module of Clostridium stercorarium Xyn10B to xylan hydrolysis. Biosci. Biotechnol. Biochem. 2001, 65, 41–47. [Google Scholar] [CrossRef]
- Bai, W.; Xue, Y.; Zhou, C.; Ma, Y. Cloning, expression, and characterization of a novel alkali-tolerant xylanase from alkaliphilic Bacillus sp. SN5. Biotechnol. Appl. Biochem. 2015, 62, 208–217. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Gullfot, F.; Tan, T.-C.; von Schantz, L.; Karlsson, E.N.; Ohlin, M.; Brumer, H.; Divne, C. The crystal structure of XG-34, an evolved xyloglucan-specific carbohydrate-binding module. Proteins 2010, 78, 785–789. [Google Scholar] [CrossRef]
- Santos, C.R.; Hoffmam, Z.B.; de Matos Martins, V.P.; Zanphorlin, L.M.; de Paula Assis, L.H.; Honorato, R.V.; de Oliveira, P.S.L.; Ruller, R.; Murakami, M.T. Molecular mechanisms associated with xylan degradation by Xanthomonas plant pathogens. J. Biol. Chem. 2014, 289, 32186–32200. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Xylanases | Activity in NaCl (%) | Strains | Reference | ||
|---|---|---|---|---|---|
| 3.0 M | 4.0 M | 5.0 M | |||
| XynRA2 | 95 | 94 | 71 | R. sacchariphilus RA strain RA | This study |
| M11 | 60 | 50 | 47 | Streptomyces viridochromogenes | [12] |
| Xyn512 | 60 | 47 | 32 | Flammeovirga pacifica WPAGA1 | [23] |
| XynSL3 | 60 | 40 | ND | Alkalibacterium sp. SL3 | [24] |
| XynA | 78 | 53 | ND | Glaciecola mesophila KMM 241 | [50] |
| XynA | 180 | 140 | 100 | Zunongwangia profunda | [49] |
| XynSL4 | 59 | 41 | ND | Planococcus sp. SL4 | [51] |
| XynRBM26 | 96 | 93 | 87 | Massilia sp. RBM26 | [52] |
| XynAHJ3 | 40 | 26 | ND | Lechevalieria sp. HJ3 | [53] |
| Xylanase | 90 | 87 | 84 | Gracillibacillus sp. TSCPVG | [54] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teo, S.C.; Liew, K.J.; Shamsir, M.S.; Chong, C.S.; Bruce, N.C.; Chan, K.-G.; Goh, K.M. Characterizing a Halo-Tolerant GH10 Xylanase from Roseithermus sacchariphilus Strain RA and Its CBM-Truncated Variant. Int. J. Mol. Sci. 2019, 20, 2284. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092284
Teo SC, Liew KJ, Shamsir MS, Chong CS, Bruce NC, Chan K-G, Goh KM. Characterizing a Halo-Tolerant GH10 Xylanase from Roseithermus sacchariphilus Strain RA and Its CBM-Truncated Variant. International Journal of Molecular Sciences. 2019; 20(9):2284. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092284
Chicago/Turabian StyleTeo, Seng Chong, Kok Jun Liew, Mohd Shahir Shamsir, Chun Shiong Chong, Neil C. Bruce, Kok-Gan Chan, and Kian Mau Goh. 2019. "Characterizing a Halo-Tolerant GH10 Xylanase from Roseithermus sacchariphilus Strain RA and Its CBM-Truncated Variant" International Journal of Molecular Sciences 20, no. 9: 2284. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20092284