IRE1-XBP1 Pathway of the Unfolded Protein Response Is Required during Early Differentiation of C2C12 Myoblasts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
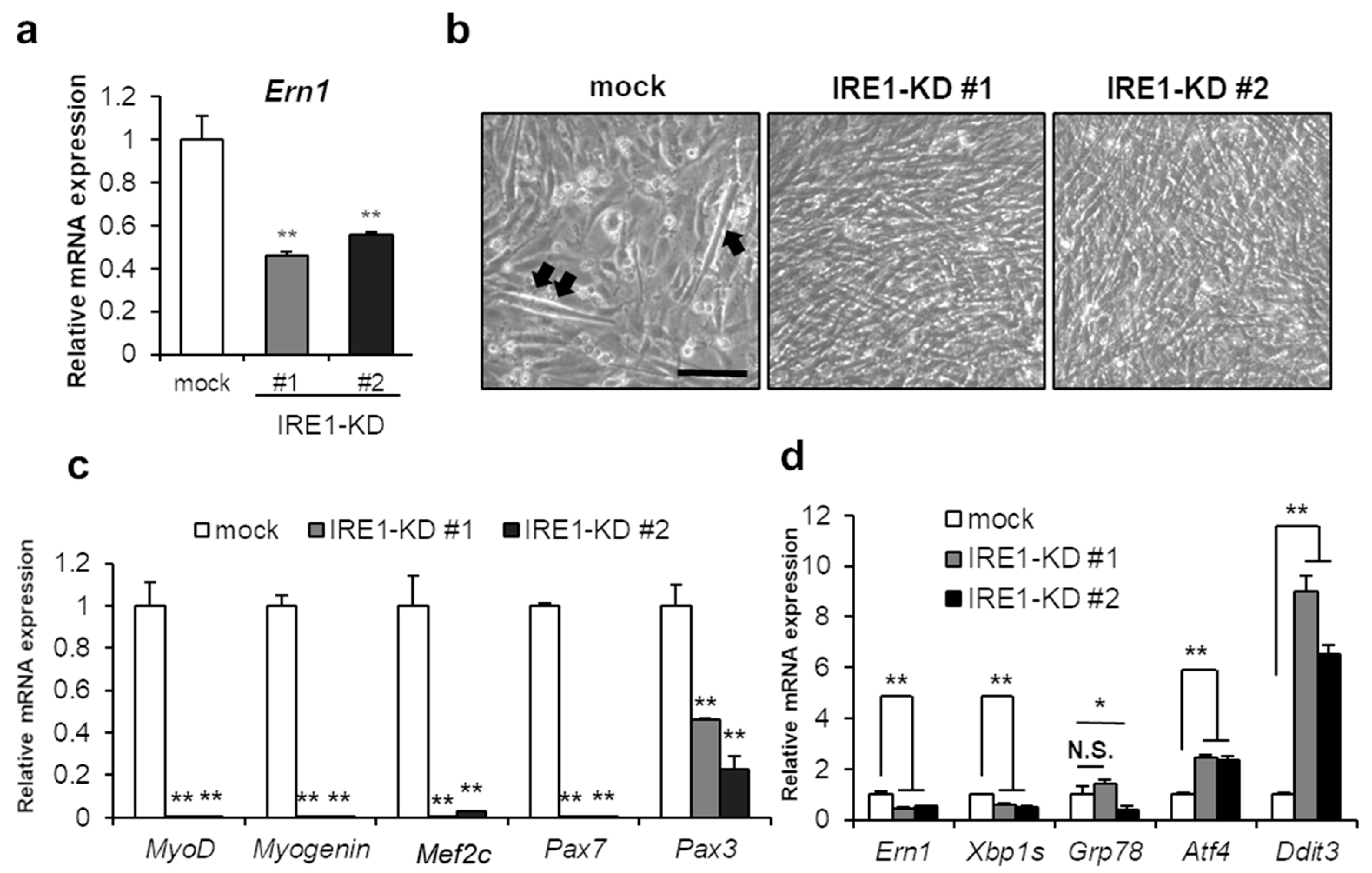
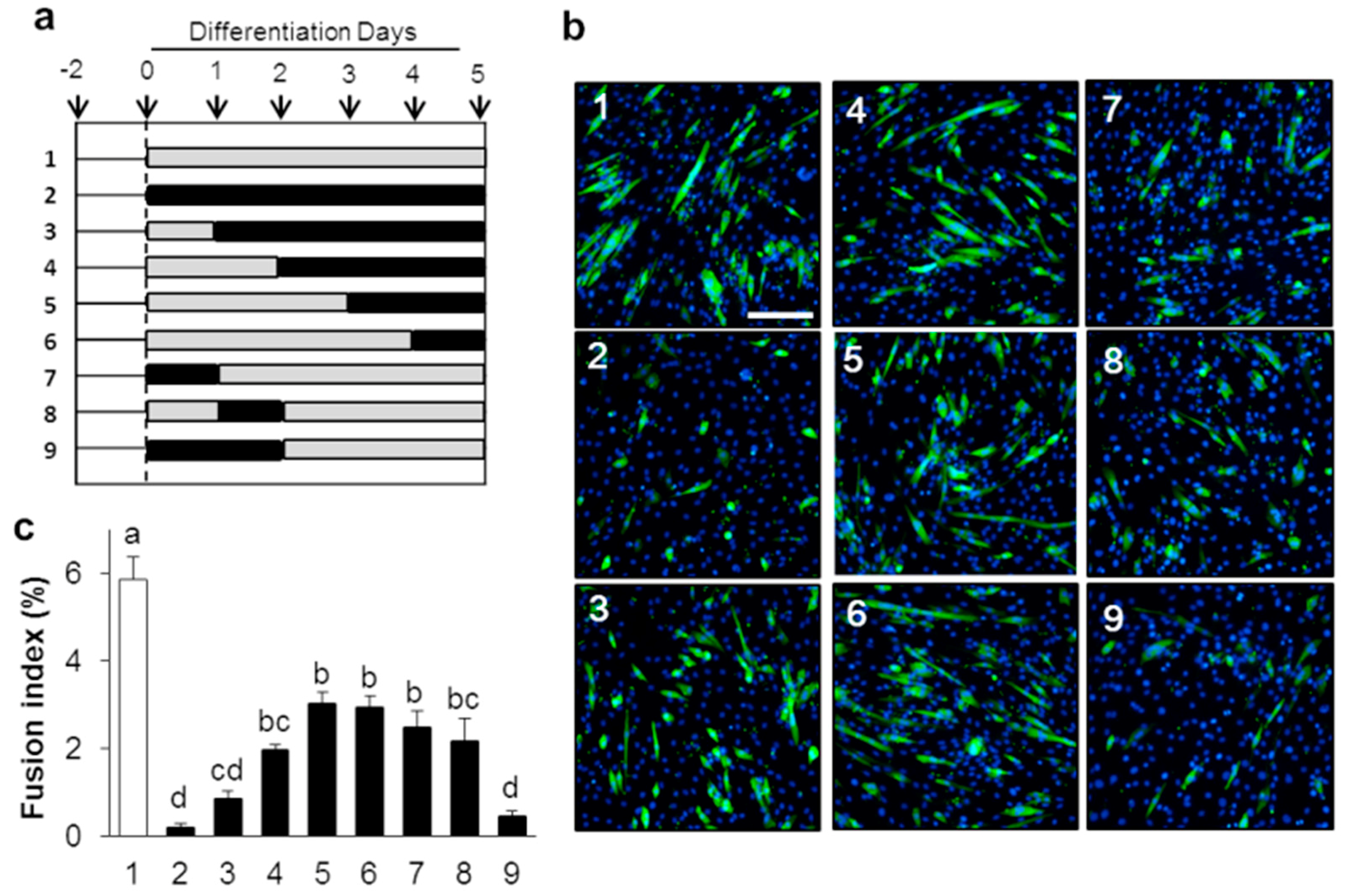
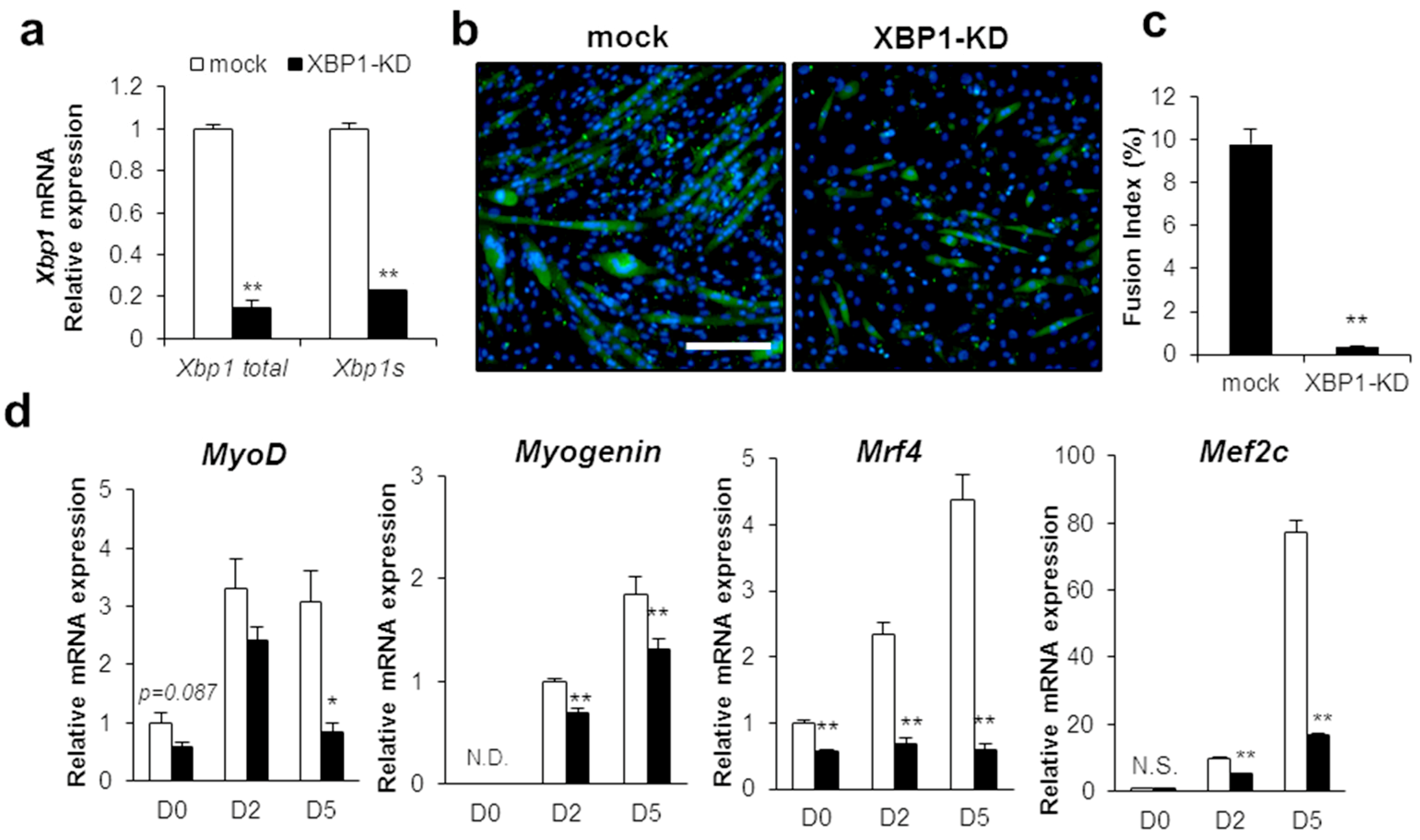
2.1. IRE1–XBP1 Is Essential for the Myotube Formation in C2C12 Differentiation
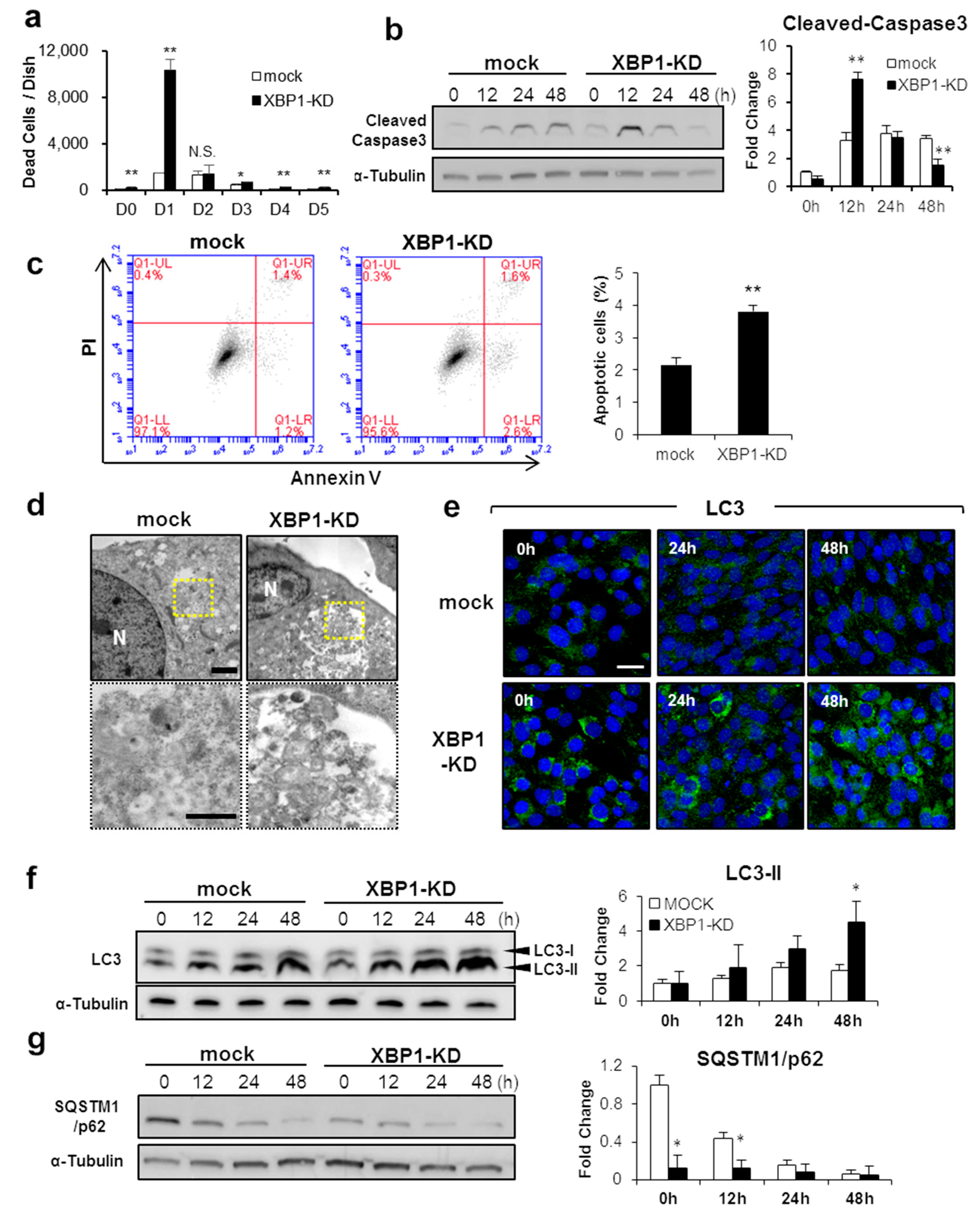
2.2. XBP1 Deficiency Perturbs Apoptosis and Autophagy during Early Myogenic Differentiation
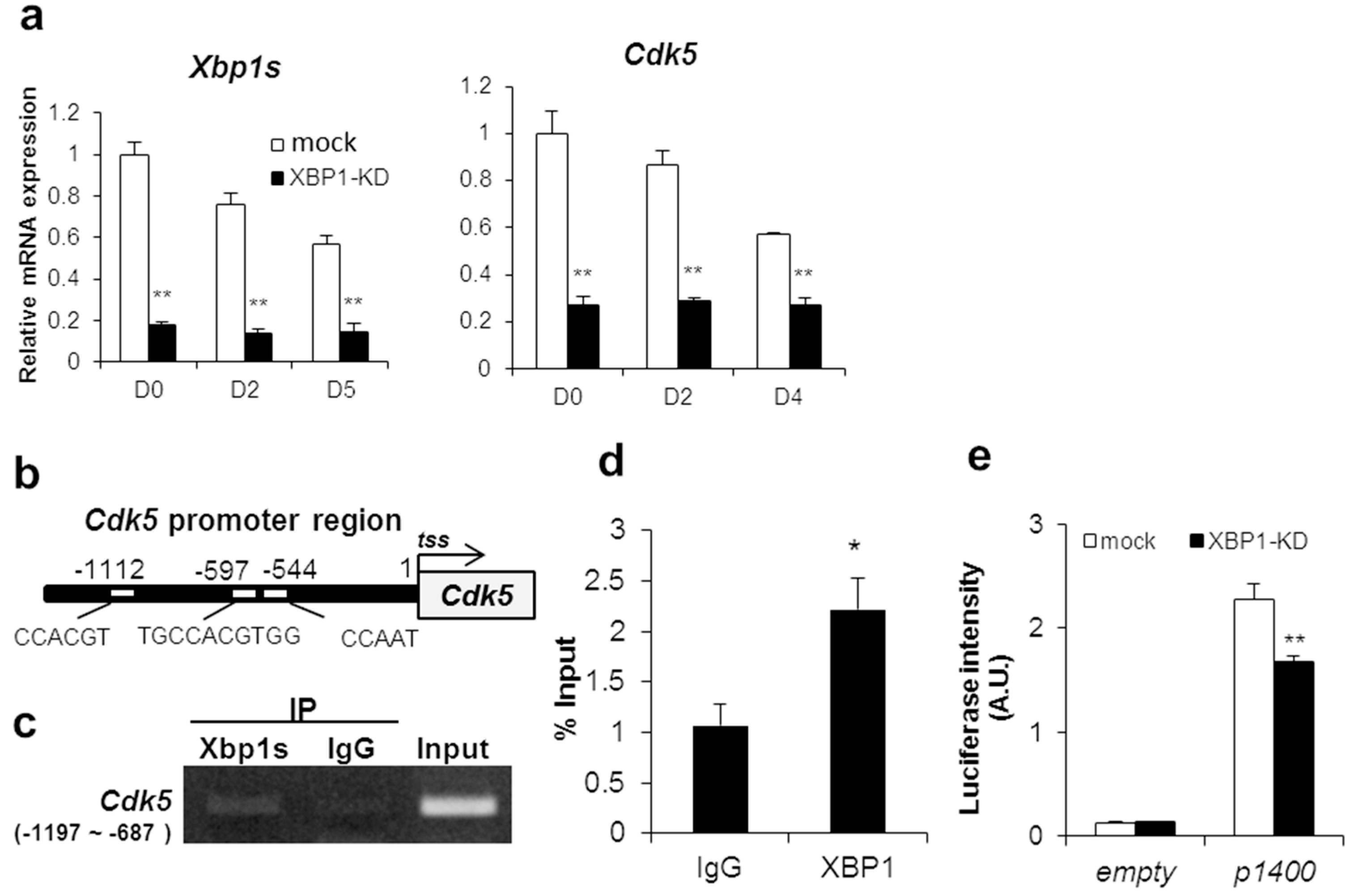
2.3. CDK5 Is a Transcriptional Target Gene of XBP1s
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Culture
4.3. Establishment of shIRE1- and shXBP1- Transduced Cells
4.4. Quantification of Myotube Formation
4.5. Apoptosis Analysis by Flow Cytometry
4.6. Transmission Electron Microscopy (TEM)
4.7. Immunocytochemistry
4.8. Immunoblotting
4.9. ChIP Assay
4.10. Luciferase Assay
4.11. RNA Extraction, Real-Time PCR Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| UPR | Unfolded Protein Response |
| IRE1 | Inositol-requiring Enzyme 1 |
| XBP1 | X-box Binding Protein 1 |
| PERK | Protein Kinase R-like Endoplasmic Reticulum Kinase |
| CHOP | C/EBP Homologous Protein |
| ATF4 | Activating Transcription Factor 4 |
| MyoD | Myogenic Differentiation 1 |
| Myf5 | Myogenic Factor 5 |
| MRF4 | Myogenic Regulatory Factor 4 |
| MEF2C | Myocyte Enhancer Factor 2 |
| CDK5 | Cyclin-dependent Kinase 5 |
References
- Berkes, C.A.; Tapscott, S.J. MyoD and the Transcriptional Control of Myogenesis. Semin. Cell Dev. Biol. 2005, 16, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.N.; Tajbakhsh, S.; Mouly, V.; Cossu, G.; Buckingham, M.; Butler-Browne, G.S. In Vivo Satellite Cell Activation via Myf5 and MyoD in Regenerating Mouse Skeletal Muscle. J. Cell Sci. 1999, 112 Pt 17, 2895–2901. [Google Scholar]
- Kablar, B.; Krastel, K.; Ying, C.Y.; Asakura, A.; Tapscott, S.J.; Rudnicki, M.A. MyoD and Myf-5 Differentially Regulate the Development of Limb Versus Trunk Skeletal Muscle. Development 1997, 124, 4729–4738. [Google Scholar] [PubMed]
- Martin, J.F.; Schwarz, J.J.; Olson, E.N. Myocyte Enhancer Factor (MEF) 2C: A Tissue-Restricted Member of the MEF-2 Family of Transcription Factors. Proc. Natl. Acad. Sci. USA 1993, 90, 5282–5286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venuti, J.M.; Morris, J.H.; Vivian, J.L.; Olson, E.N.; Klein, W.H. Myogenin is Required for Late But Not Early Aspects of Myogenesis during Mouse Development. J. Cell Biol. 1995, 128, 563–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, K.; Kakiguchi, K.; Yonemura, S.; Nakano, A.; Morishima, N. Transient Ca2+ Depletion from the Endoplasmic Reticulum is Critical for Skeletal Myoblast Differentiation. FASEB J. 2015, 29, 2137–2149. [Google Scholar] [CrossRef] [PubMed]
- Sin, J.; Andres, A.M.; Taylor, D.J.R.; Weston, T.; Hiraumi, Y.; Stotland, A.; Kim, B.J.; Huang, C.Q.; Doran, K.S.; Gottlieb, R.A. Mitophagy is Required for Mitochondrial Biogenesis and Myogenic Differentiation of C2C12 Myoblasts. Autophagy 2016, 12, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Rayavarapu, S.; Coley, W.; Nagaraju, K. Endoplasmic Reticulum Stress in Skeletal Muscle Homeostasis and Disease. Curr. Rheumatol. Rep. 2012, 14, 238–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, K.; Dohmae, N.; Morishima, N. Endoplasmic Reticulum Stress Increases Myofiber Formation In Vitro. FASEB. J. 2007, 21, 2994–3003. [Google Scholar] [CrossRef]
- Hetz, C.; Martinon, F.; Rodriguez, D.; Glimcher, L.H. The Unfolded Protein Response: Integrating Stress Signals through the Stress Sensor Ire1 Alpha. Physiol. Rev. 2011, 91, 1219–1243. [Google Scholar] [CrossRef]
- Woehlbier, U.; Hetz, C. Modulating Stress Responses by the UPRosome: A Matter of Life and Death. Trends Biochem. Sci. 2011, 36, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C.; Glimcher, L.H. Fine-Tuning of the Unfolded Protein Response: Assembling the IRE1 alpha Interactome. Mol. Cell 2009, 35, 551–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolotti, A.; Zhang, Y.H.; Hendershot, L.M.; Harding, H.P.; Ron, D. Dynamic Interaction of BiP and ER Stress Transducers in the Unfolded-Protein Response. Nat. Cell Biol. 2000, 2, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Bagratuni, T.; Davenport, E.L.; Nowak, P.R.; Silva-Santisteban, M.C.; Hardcastle, A.; McAndrews, C.; Rowlands, M.G.; Morgan, G.J.; Aherne, W.; et al. Structure of the Ire1 Autophosphorylation Complex and Implications for the Unfolded Protein Response. EMBO J. 2011, 30, 894–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calfon, M.; Zeng, H.Q.; Urano, F.; Till, J.H.; Hubbard, S.R.; Harding, H.P.; Clark, S.G.; Ron, D. IRE1 Couples Endoplasmic Reticulum Load to Secretory Capacity by Processing the XBP-1 mRNA. Nature 2002, 415, 92–96. [Google Scholar] [CrossRef]
- Yoshida, H.; Matsui, T.; Yamamoto, A.; Okada, T.; Mori, K. XBP1 mRNA is Induced by ATF6 and Spliced by IRE1 in Response to ER Stress to Produce a Highly Active Transcription Factor. Cell 2001, 107, 881–891. [Google Scholar] [CrossRef] [Green Version]
- Olivari, S.; Galli, C.; Alanen, H.; Ruddock, L.; Molinari, M. A Novel Stress-Induced EDEM Variant Regulating Endoplasmic Reticulum-Associated Glycoprotein Degradation. J. Biol. Chem. 2005, 280, 2424–2428. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Yoshida, H.; Kokame, K.; Kaufman, R.J.; Mori, K. Differential Contributions of ATF6 and XBP1 to the Activation of Endoplasmic Reticulum Stress-Responsive Cis-Acting Elements ERSE, UPRE and ERSE-II. J. Biochem. 2004, 136, 343–350. [Google Scholar] [CrossRef]
- Harding, H.P.; Novoa, I.; Zhang, Y.H.; Zeng, H.Q.; Wek, R.; Schapira, M.; Ron, D. Regulated Translation Initiation Controls Stress-Induced Gene Expression in Mammalian Cells. Mol. Cell 2000, 6, 1099–1108. [Google Scholar] [CrossRef]
- Harding, H.P.; Zhang, Y.; Zeng, H.; Novoa, I.; Lu, P.D.; Calfon, M.; Sadri, N.; Yun, C.; Popko, B.; Paules, R.; et al. An Integrated Stress Response Regulates Amino Acid Metabolism and Resistance to Oxidative Stress. Mol. Cell 2003, 11, 619–633. [Google Scholar] [CrossRef]
- Haze, K.; Yoshida, H.; Yanagi, H.; Yura, T.; Mori, K. Mammalian Transcription Factor ATF6 is Synthesized as a Transmembrane Protein and Activated by Proteolysis in Response to Endoplasmic Reticulum Stress. Mol. Biol. Cell 1999, 10, 3787–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, J.; Murthy, R.; Wood, B.; Song, B.; Wang, S.; Sun, B.; Malhi, H.; Kaufman, R.J. ER Stress Signalling through eIF2 Alpha and CHOP, but not IRE1 Alpha, Attenuates Adipogenesis in Mice. Diabetologia 2013, 56, 911–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimold, A.M.; Etkin, A.; Clauss, I.; Perkins, A.; Friend, D.S.; Zhang, J.; Horton, H.F.; Scott, A.; Orkin, S.H.; Byrne, M.C.; et al. An Essential Role in Liver Development for Transcription Factor XBP-1. Genes Dev. 2000, 14, 152–157. [Google Scholar] [PubMed]
- Reimold, A.M.; Iwakoshi, N.N.; Manis, J.; Vallabhajosyula, P.; Szomolanyi-Tsuda, E.; Gravallese, E.M.; Friend, D.; Grusby, M.J.; Alt, F.; Glimcher, L.H. Plasma Cell Differentiation Requires the Transcription Factor XBP-1. Nature 2001, 412, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Tohmonda, T.; Miyauchi, Y.; Ghosh, R.; Yoda, M.; Uchikawa, S.; Takito, J.; Morioka, H.; Nakamura, M.; Iwawaki, T.; Chiba, K.; et al. The IRE1alpha-XBP1 Pathway is Essential for Osteoblast Differentiation through Promoting Transcription of Osterix. EMBO Rep. 2011, 12, 451–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, M.; Koizumi, Y.; Hayashi, S.; Hanaoka, M.; Tokutake, Y.; Yonekura, S. The Role of Unfolded Protein Response in Differentiation of Mammary Epithelial Cells. Biochem. Biophys. Res. Commun. 2017, 484, 903–908. [Google Scholar] [CrossRef]
- Nakanishi, K.; Sudo, T.; Morishima, N. Endoplasmic Reticulum Stress Signaling Transmitted by ATF6 Mediates Apoptosis during Muscle Development. J. Cell Biol. 2005, 169, 555–560. [Google Scholar] [CrossRef]
- Xiong, G.Y.; Hindi, S.M.; Mann, A.K.; Gallot, Y.S.; Bohnert, K.R.; Cavener, D.R.; Whittemore, S.R.; Kumar, A. The PERK Arm of the Unfolded Protein Response Regulates Satellite Cell-Mediated Skeletal Muscle Regeneration. Elife 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, A.L.; Mitne-Neto, M.; Silva, H.C.A.; Richieri-Costa, A.; Middleton, S.; Cascio, D.; Kok, F.; Oliveira, J.R.M.; Gillingwater, T.; Webb, J.; et al. A Mutation in the Vesicle-Trafficking Protein VAPB Causes Late-Onset Spinal Muscular Atrophy and Amyotrophic Lateral Sclerosis. Am. J. Hum. Genet. 2004, 75, 822–831. [Google Scholar] [CrossRef] [Green Version]
- Tokutake, Y.; Yamada, K.; Ohata, M.; Obayashi, Y.; Tsuchiya, M.; Yonekura, S. ALS-Linked P56S-VAPB Mutation Impairs the Formation of Multinuclear Myotube in C2C12 Cells. Int. J. Mol. Sci. 2015, 16, 18628–18641. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Alvear, D.; Zhou, Y.; Blais, A.; Tsikitis, M.; Lents, N.H.; Arias, C.; Lennon, C.J.; Kluger, Y.; Dynlacht, B.D. XBP1 Controls Diverse Cell Type-and Condition-Specific Transcriptional Regulatory Networks. Mol. Cell 2007, 27, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Papandreou, I.; Denko, N.C.; Olson, M.; Van Melckebeke, H.; Lust, S.; Tam, A.; Solow-Cordero, D.E.; Bouley, D.M.; Offner, F.; Niwa, M.; et al. Identification of an Ire1alpha Endonuclease Specific Inhibitor with Cytotoxic Activity Against Human Multiple Myeloma. Blood 2011, 117, 1311–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandri, M.; Carraro, U. Apoptosis of Skeletal Muscles during Development and Disease. Int. J. Biochem. Cell Biol. 1999, 31, 1373–1390. [Google Scholar] [CrossRef]
- Fernando, P.; Kelly, J.F.; Balazsi, K.; Slack, R.S.; Megeney, L.A. Caspase 3 Activity is Required for Skeletal Muscle Differentiation. Proc. Natl. Acad. Sci. USA 2002, 99, 11025–11030. [Google Scholar] [CrossRef] [Green Version]
- Hochreiter-Hufford, A.E.; Lee, C.S.; Kinchen, J.M.; Sokolowski, J.D.; Arandjelovic, S.; Call, J.A.; Klibanov, A.L.; Yan, Z.; Mandell, J.W.; Ravichandran, K.S. Phosphatidylserine Receptor BAI1 and Apoptotic Cells as New Promoters of Myoblast fusion. Nature 2013, 497, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMillan, E.M.; Quadrilatero, J. Autophagy is Required and Protects Against Apoptosis during Myoblast Differentiation. Biochem. J. 2014, 462, 267–277. [Google Scholar] [CrossRef]
- Philpott, A.; Porro, E.B.; Kirschner, M.W.; Tsai, L.H. The Role of Cyclin-Dependent Kinase 5 and a Novel Regulatory Subunit in Regulating Muscle Differentiation and Patterning. Genes Dev. 1997, 11, 1409–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras-Vallejos, E.; Utreras, E.; Gonzalez-Billault, C. Going Out of the Brain: Non-Nervous System Physiological and Pathological Functions of Cdk5. Cell. Signal. 2012, 24, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Zhu, H.; Morishima, N.; Li, E.; Xu, J.; Yankner, B.A.; Yuan, J. Caspase-12 Mediates Endoplasmic-Reticulum-Specific Apoptosis and Cytotoxicity by Amyloid-Beta. Nature 2000, 403, 98–103. [Google Scholar] [CrossRef]
- Collins, C.A.; Gnocchi, V.F.; White, R.B.; Boldrin, L.; Perez-Ruiz, A.; Relaix, F.; Morgan, J.E.; Zammit, P.S. Integrated Functions of Pax3 and Pax7 in the Regulation of Proliferation, Cell Size and Myogenic Differentiation. PLoS ONE 2009, 4, e4475. [Google Scholar] [CrossRef] [Green Version]
- Jheng, J.R.; Chen, Y.S.; Ao, U.I.; Chan, D.C.; Huang, J.W.; Hung, K.Y.; Tarng, D.C.; Chiang, C.K. The Double-Edged Sword of Endoplasmic Reticulum Stress in Uremic Sarcopenia through Myogenesis Perturbation. J. Cachexia Sarcopenia Muscle 2018, 9, 570–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassar-Duchossoy, L.; Gayraud-Morel, B.; Gomes, D.; Rocancourt, D.; Buckingham, M.; Shinin, V.; Tajbakhsh, S. Mrf4 Determines Skeletal Muscle Identity in Myf5:Myod Double-Mutant Mice. Nature 2004, 431, 466–471. [Google Scholar] [CrossRef]
- Pavlath, G.K.; Dominov, J.A.; Kegley, K.M.; Miller, J.B. Regeneration of Transgenic Skeletal Muscles with Altered Timing of Expression of the Basic Helix-Loop-Helix Muscle Regulatory Factor MRF4. Am. J. Pathol. 2003, 162, 1685–1691. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Bornemann, A. MRF4 Protein Expression in Regenerating Rat Muscle. J. Muscle Res. Cell Motil. 2001, 22, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.; Guan, K.L.; Kim, J. AMPK and Autophagy in Glucose/Glycogen Metabolism. Mol. Asp. Med. 2015, 46, 46–62. [Google Scholar] [CrossRef]
- Mihaylova, M.M.; Shaw, R.J. The AMPK Signalling Pathway Coordinates Cell Growth, Autophagy and Metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef]
- Lazaro, J.B.; Kitzmann, M.; Poul, M.A.; Vandromme, M.; Lamb, N.J.; Fernandez, A. Cyclin Dependent kinase 5, cdk5, is a Positive Regulator of Myogenesis in Mouse C2 Cells. J. Cell Sci. 1997, 110 Pt 10, 1251–1260. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokutake, Y.; Yamada, K.; Hayashi, S.; Arai, W.; Watanabe, T.; Yonekura, S. IRE1-XBP1 Pathway of the Unfolded Protein Response Is Required during Early Differentiation of C2C12 Myoblasts. Int. J. Mol. Sci. 2020, 21, 182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010182
Tokutake Y, Yamada K, Hayashi S, Arai W, Watanabe T, Yonekura S. IRE1-XBP1 Pathway of the Unfolded Protein Response Is Required during Early Differentiation of C2C12 Myoblasts. International Journal of Molecular Sciences. 2020; 21(1):182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010182
Chicago/Turabian StyleTokutake, Yukako, Keita Yamada, Satoko Hayashi, Wataru Arai, Takafumi Watanabe, and Shinichi Yonekura. 2020. "IRE1-XBP1 Pathway of the Unfolded Protein Response Is Required during Early Differentiation of C2C12 Myoblasts" International Journal of Molecular Sciences 21, no. 1: 182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010182