CX3CR1-Targeted PLGA Nanoparticles Reduce Microglia Activation and Pain Behavior in Rats with Spinal Nerve Ligation

 , , and
, , and 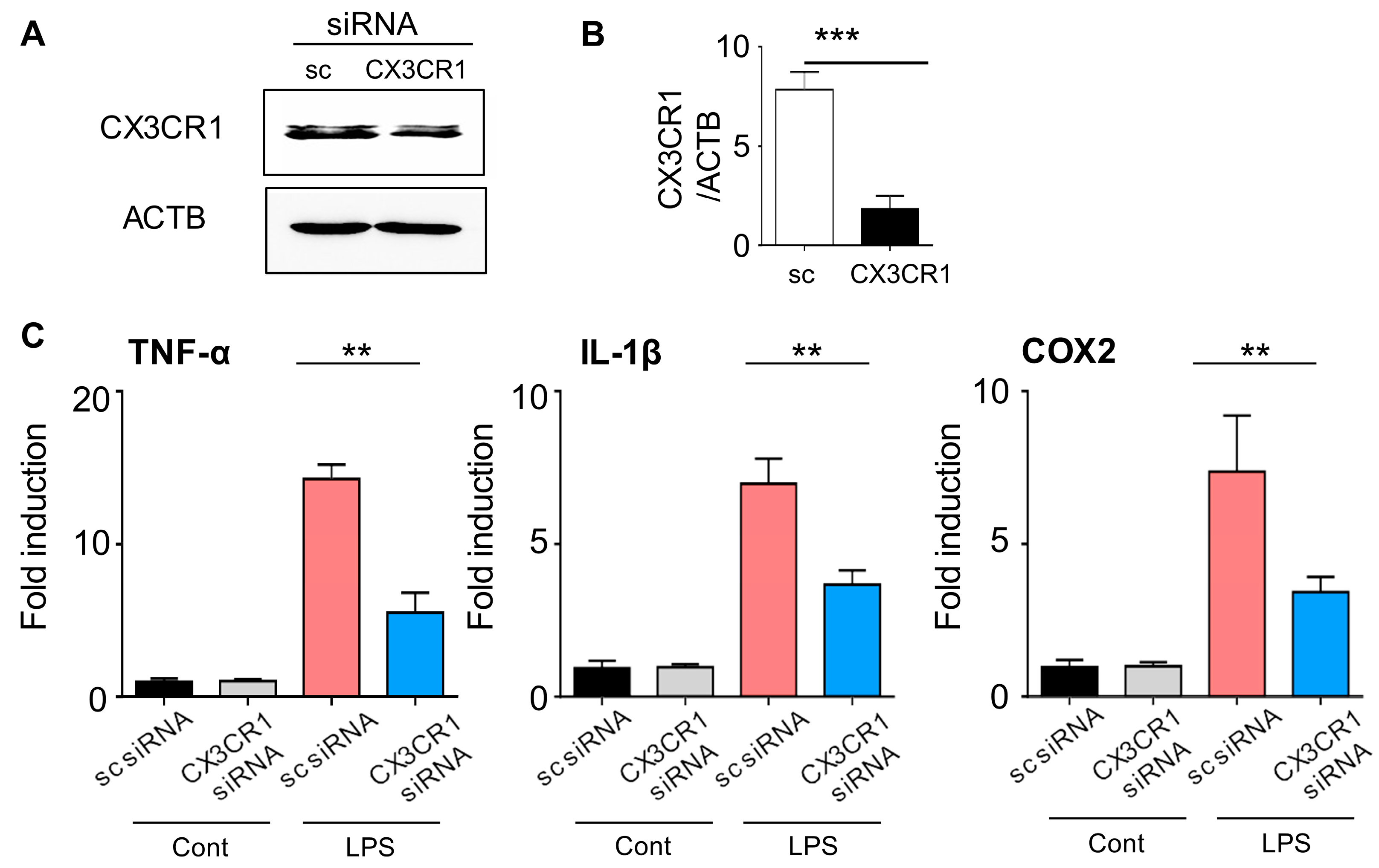{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. CX3CR1 siRNA Blocks CX3CR1 Expression in BV2 Cells and Reduces Expression of Pro-Inflammatory Mediators in Lipopolysaccharide-Stimulated BV2 Cells
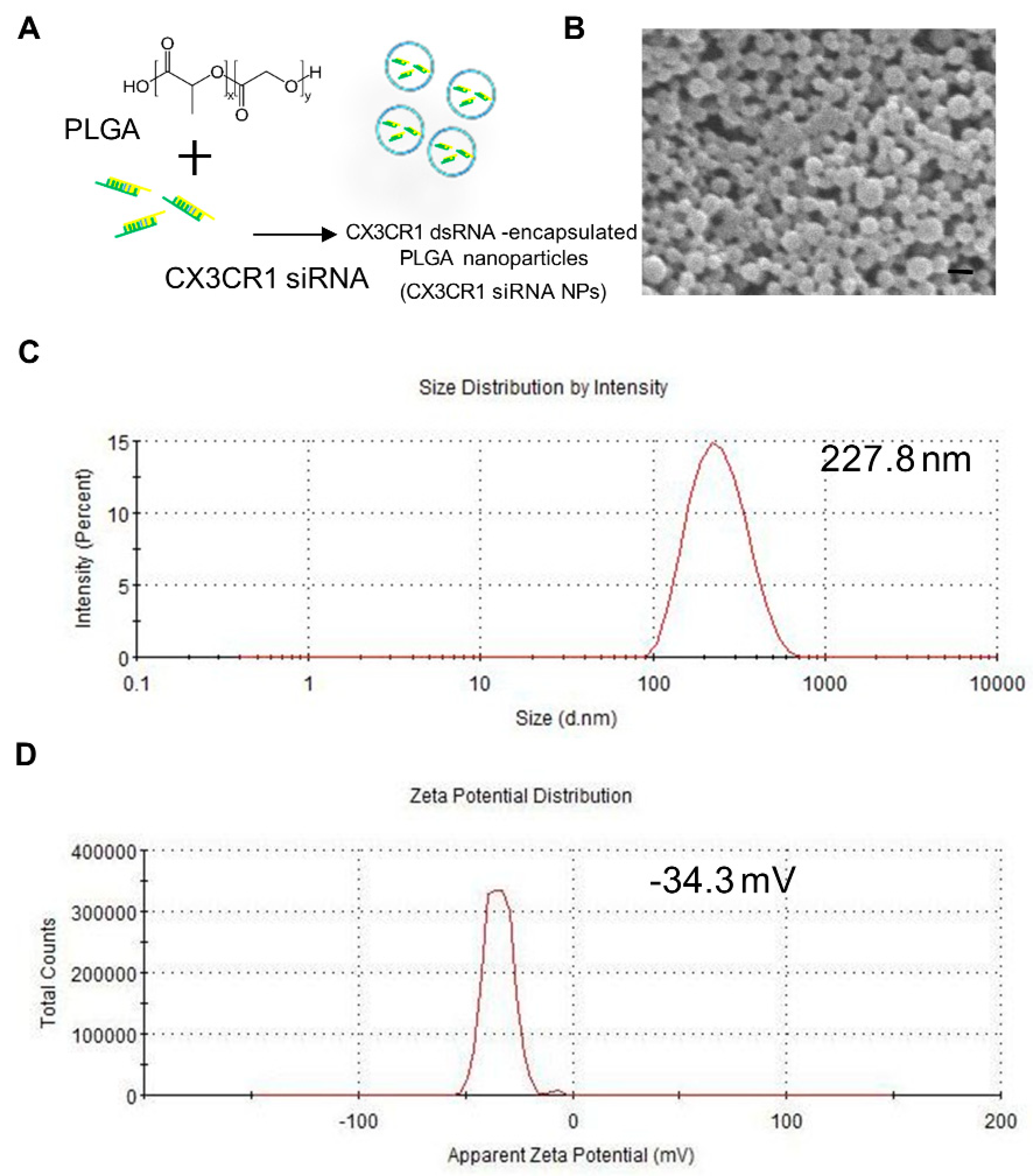
2.2. Preparation and Characterization of PLGA-Encapsulated CX3CR1 siRNA Nanoparticles
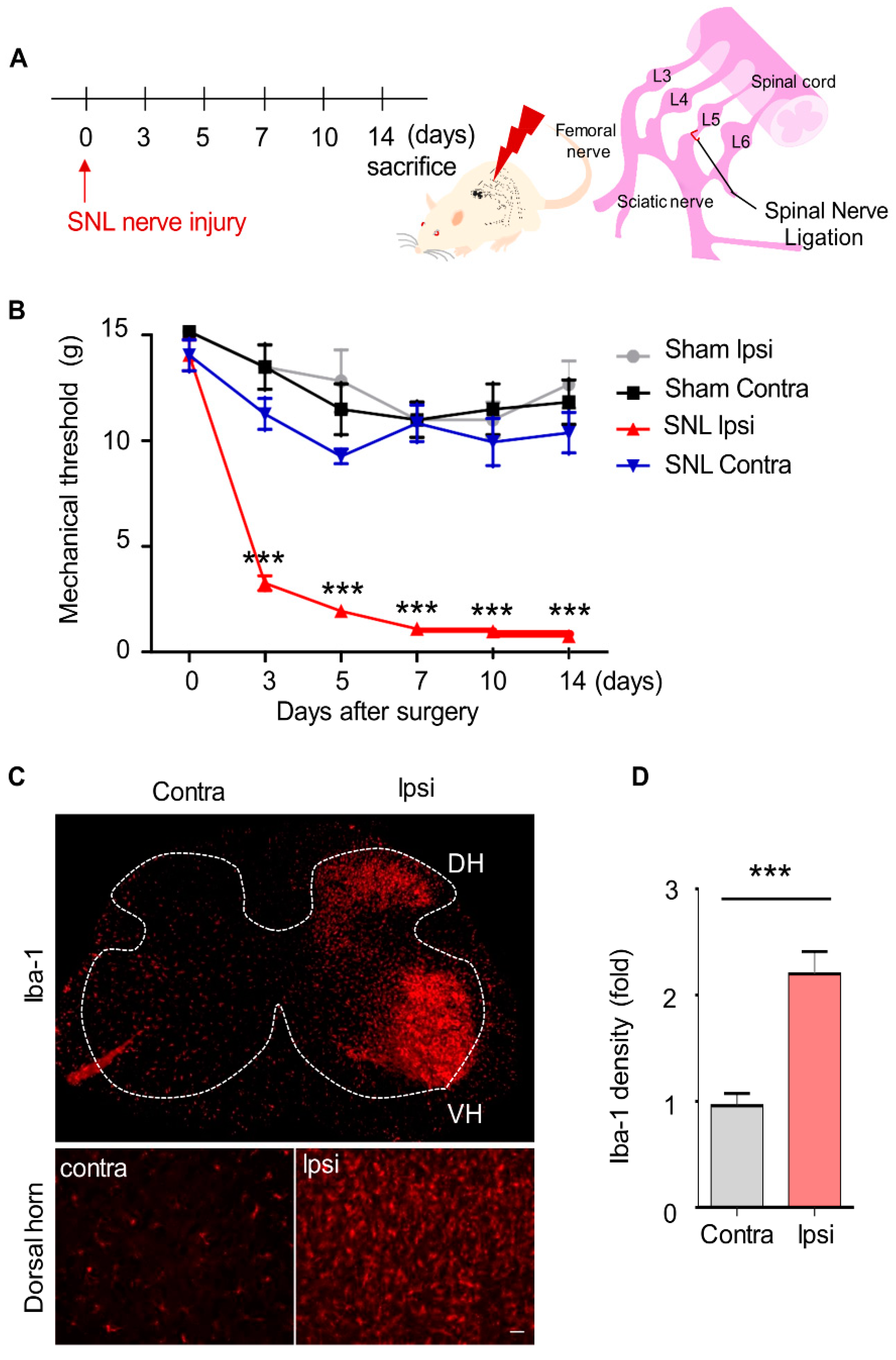
2.3. Mechanical Allodynia and Upregulated Microglia Activation in SNL-Induced Rats
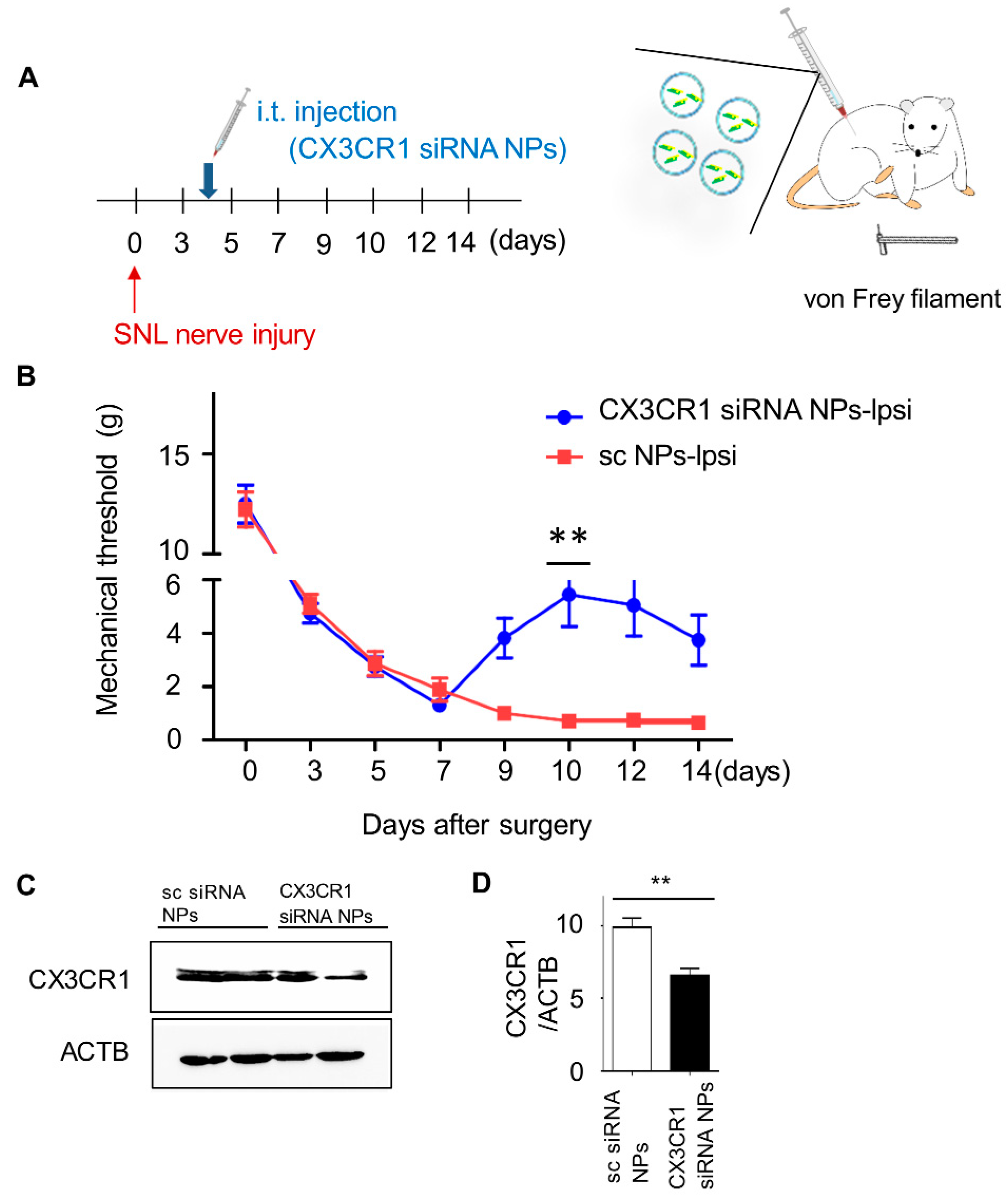
2.4. Intrathecal Injection of PLGA-Encapsulated CX3CR1 siRNA Nanoparticles Reduces Mechanical Allodynia in SNL-Induced Rats
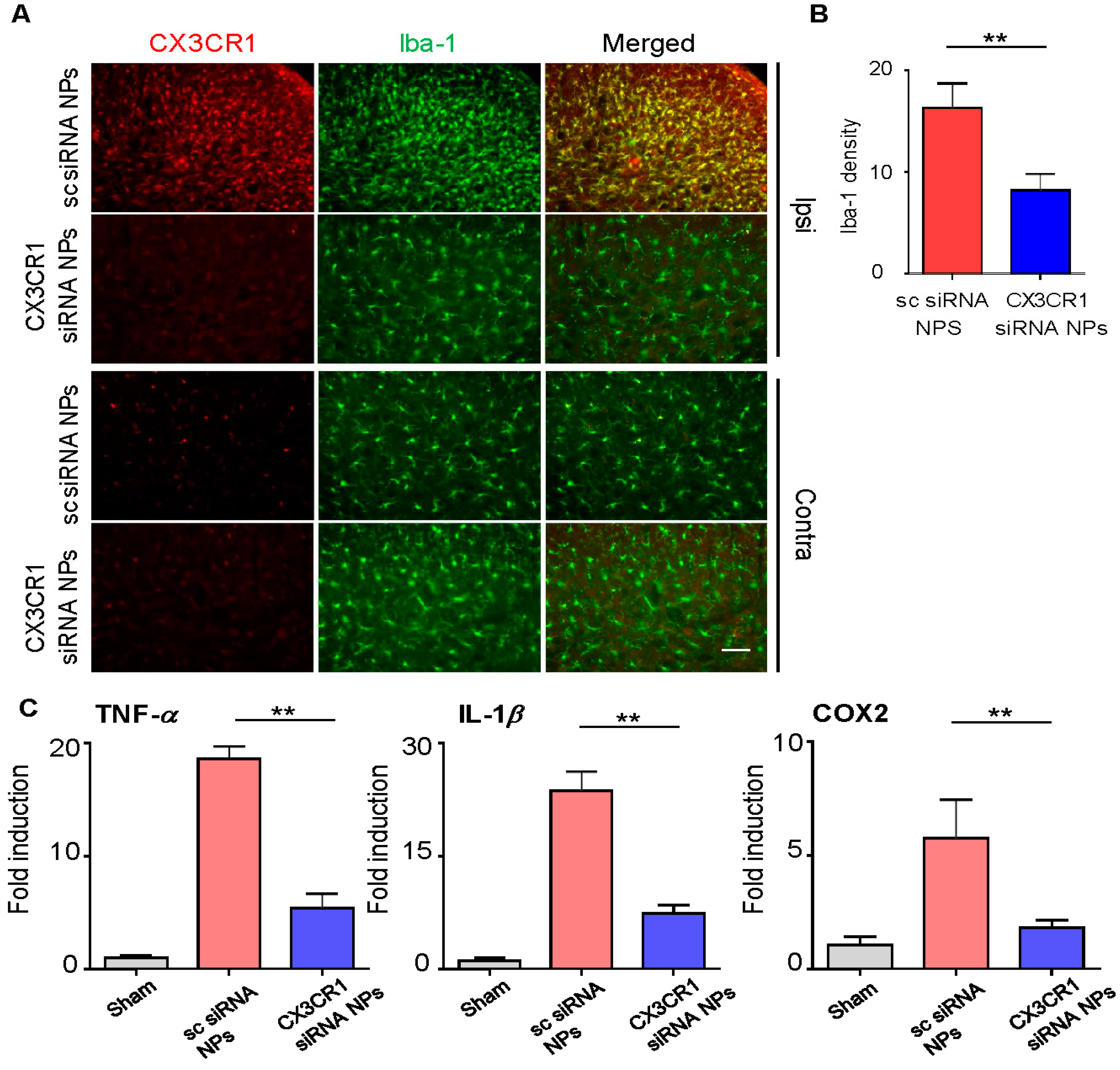
2.5. PLGA-Encapsulated CX3CR1 siRNA Nanoparticles Reduce Microglial Activation and Downregulate the Expression of Neuropathic Pain-Related Genes in the Spinal Dorsal Horn of the SNL-Induced Rat Spinal Cord
3. Discussion
4. Materials and Methods
4.1. Animals and Materials
4.2. siRNA Transfection into BV2 Cells
4.3. Western Blot and Immunohistochemistry
4.4. Quantitative RT-PCR
4.5. PLGA Nanoparticle Preparation
4.6. Induction of Neuropathic Pain by L5 SNL
4.7. Pain Behavior Test with Von Frey Filaments
4.8. Intrathecal Injection
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| SNL | Spinal nerve ligation |
| PLGA | Poly(D,L-lactic-co-glycolic acid) |
| siRNA | Small interfering RNA |
| LPS | Lipopolysaccharide |
| NPs | Nanoparticles |
References
- Jensen, T.S.; Baron, R.; Haanpää, M.; Kalso, E.; Loeser, J.D.; Rice, A.P.; Treede, R.-D. A new definition of neuropathic pain. Pain 2011, 152, 2204–2205. [Google Scholar] [CrossRef]
- Zhuang, Z.-Y.; Kawasaki, Y.; Tan, P.-H.; Wen, Y.-R.; Huang, J.; Ji, R.-R. Role of the CX3CR1/p38 MAPK pathway in spinal microglia for the development of neuropathic pain following nerve injury-induced cleavage of fractalkine. Brain Behav. Immun. 2007, 21, 642–651. [Google Scholar] [CrossRef] [Green Version]
- Chaban, V.V. Peripheral sensitization of sensory neurons. Ethn. Dis. 2010, 20, S1–S3. [Google Scholar]
- Ma, Z.; Han, Q.; Wang, X.; Ai, Z.-S.; Zheng, Y. Galectin-3 Inhibition Is Associated with Neuropathic Pain Attenuation after Peripheral Nerve Injury. PLoS ONE 2016, 11, e0148792. [Google Scholar] [CrossRef] [Green Version]
- Psaroulaki, A.; Antoniou, M.; Toumazos, P.; Mazeris, A.; Ioannou, I.; Chochlakis, D.; Christophi, N.; Loukaides, P.; Patsias, A.; Moschandrea, I.; et al. Rats as indicators of the presence and dispersal of six zoonotic microbial agents in Cyprus, an island ecosystem: A seroepidemiological study. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 733–739. [Google Scholar] [CrossRef]
- Austin, P.J.; Moalem-Taylor, G. The neuro-immune balance in neuropathic pain: Involvement of inflammatory immune cells, immune-like glial cells and cytokines. J. Neuroimmunol. 2010, 229, 26–50. [Google Scholar] [CrossRef]
- Morioka, N.; Fujii, S.; Kondo, S.; Zhang, F.F.; Miyauchi, K.; Nakamura, Y.; Hisaoka-Nakashima, K.; Nakata, Y. Downregulation of spinal astrocytic connexin43 leads to upregulation of interleukin-6 and cyclooxygenase-2 and mechanical hypersensitivity in mice. Glia 2017, 66, 428–444. [Google Scholar] [CrossRef]
- Yin, Y.; Phạm, T.L.; Shin, J.; Shin, N.; Kang, D.-W.; Lee, S.Y.; Lee, W.; Kim, C.-S.; Kim, S.R.; Hong, J.; et al. Arginase 2 Deficiency Promotes Neuroinflammation and Pain Behaviors Following Nerve Injury in Mice. J. Clin. Med. 2020, 9, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Hong, J.; Pham, T.L.; Shin, J.; Gwon, D.H.; Kwon, H.H.; Shin, N.; Shin, H.J.; Lee, S.Y.; Lee, W.H.; et al. Evans Blue Reduces Neuropathic Pain Behavior by Inhibiting Spinal ATP Release. Int. J. Mol. Sci. 2019, 20, 4443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, M.-H.; Shin, J.; Shin, N.; Yin, Y.; Lee, S.Y.; Kim, C.-S.; Kim, S.R.; Zhang, E.; Kim, D.W. PINK1 mediates spinal cord mitophagy in neuropathic pain. J. Pain Res. 2019, 12, 1685–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Park, H.; Lee, S.Y.; Lee, W.-H.; Song, H.-J.; Kim, J.; Kim, D.W.; Hong, J. Analgesic Effect of Toll-like Receptor 4 Antagonistic Peptide 2 on Mechanical Allodynia Induced with Spinal Nerve Ligation in Rats. Exp. Neurobiol. 2019, 28, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Bazan, J.F.; Bacon, K.B.; Hardiman, G.; Wang, W.; Soo, K.; Rossi, D.; Greaves, D.R.; Zlotnik, A.; Schall, T.J. A new class of membrane-bound chemokine with a CX3C motif. Nature 1997, 385, 640–644. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.K.; Jiang, Y.; Chen, S.; Xia, Y.; Maciejewski, D.; McNamara, R.K.; Streit, W.J.; Salafranca, M.N.; Adhikari, S.; Thompson, D.; et al. Role for neuronally derived fractalkine in mediating interactions between neurons and CX3CR1-expressing microglia. Proc. Natl. Acad. Sci. USA 1998, 95, 10896–10901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbadie, C.; Lindia, J.A.; Cumiskey, A.M.; Peterson, L.B.; Mudgett, J.S.; Bayne, E.K.; DeMartino, J.A.; MacIntyre, D.E.; Forrest, M.J. Impaired neuropathic pain responses in mice lacking the chemokine receptor CCR2. Proc. Natl. Acad. Sci. USA 2003, 100, 7947–7952. [Google Scholar] [CrossRef] [Green Version]
- Johnston, I.N.; Milligan, E.D.; Wieseler-Frank, J.; Frank, M.G.; Zapata, V.; Campisi, J.; Langer, S.; Martín, D.; Green, P.; Fleshner, M.; et al. A Role for Proinflammatory Cytokines and Fractalkine in Analgesia, Tolerance, and Subsequent Pain Facilitation Induced by Chronic Intrathecal Morphine. J. Neurosci. 2004, 24, 7353–7365. [Google Scholar] [CrossRef] [Green Version]
- Lindia, J.; McGowan, E.; Jochnowitz, N.; Abbadie, C. Induction of CX3CL1 Expression in Astrocytes and CX3CR1 in Microglia in the Spinal Cord of a Rat Model of Neuropathic Pain. J. Pain 2005, 6, 434–438. [Google Scholar] [CrossRef]
- Chen, G.; Wang, Y.; Xie, R.; Gong, S. Tumor-targeted pH/redox dual-sensitive unimolecular nanoparticles for efficient siRNA delivery. J. Control. Release 2017, 259, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Pantazis, P.; Dimas, K.; Wyche, J.H.; Anant, S.; Houchen, C.W.; Panyam, J.; Ramanujam, R.P. Preparation of siRNA-Encapsulated PLGA Nanoparticles for Sustained Release of siRNA and Evaluation of Encapsulation Efficiency. In Nanoparticles in Biology and Medicine; Springer: Berlin/Heidelberg, Germany, 2012; pp. 311–319. [Google Scholar]
- Menon, J.U.; Kuriakose, A.; Iyer, R.; Hernandez, E.; Gandee, L.; Zhang, S.; Takahashi, M.; Zhang, Z.; Saha, D.; Nguyen, K.T. Dual-Drug Containing Core-Shell Nanoparticles for Lung Cancer Therapy. Sci. Rep. 2017, 7, 13249. [Google Scholar] [CrossRef] [Green Version]
- Berrocoso, E.; Rey-Brea, R.; Arevalo, M.F.; Mico, J.A.; Martín-Banderas, L. Single oral dose of cannabinoid derivate loaded PLGA nanocarriers relieves neuropathic pain for eleven days. Nanomed.: Nanotechnol. Boil. Med. 2017, 13, 2623–2632. [Google Scholar] [CrossRef]
- Campolongo, M.J.; Luo, D. Drug delivery: Old polymer learns new tracts. Nat. Mater. 2009, 8, 447–448. [Google Scholar] [CrossRef]
- Hippenstiel, S.; Soeth, S.; Kellas, B.; Fuhrmann, O.; Seybold, J.; Krüll, M.; Eichel-Streiber, C.v.; Goebeler, M.; Ludwig, S.; Suttorp, N. Rho proteins and the p38-MAPK pathway are important mediators for LPS-induced interleukin-8 expression in human endothelial cells. Blood J. Am. Soc. Hematol. 2000, 95, 3044–3051. [Google Scholar]
- Shin, H.J.; Park, H.; Shin, N.; Kwon, H.H.; Yin, Y.; Hwang, J.-A.; I Kim, S.; Kim, S.R.; Kim, S.; Joo, Y.; et al. p47phox siRNA-Loaded PLGA Nanoparticles Suppress ROS/Oxidative Stress-Induced Chondrocyte Damage in Osteoarthritis. Polymers 2020, 12, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.; Yin, Y.; Kim, K.; Lee, S.Y.; Lee, W.; Kang, J.W.; Kim, D.W.; Hong, J. Foxp3 plasmid-encapsulated PLGA nanoparticles attenuate pain behavior in rats with spinal nerve ligation. Nanomed.: Nanotechnol. Boil. Med. 2019, 18, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Yin, Y.; Park, H.; Park, S.; Triantafillu, U.L.; Kim, Y.; Kim, S.R.; Lee, S.Y.; Kim, D.K.; Hong, J.; et al. p38 siRNA-encapsulated PLGA nanoparticles alleviate neuropathic pain behavior in rats by inhibiting microglia activation. Nanomedicine 2018, 13, 1607–1621. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Chung, J.M. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain 1992, 50, 355–363. [Google Scholar] [CrossRef]
- Mu, A.; Weinberg, E.; Moulin, D.E.; Clarke, H. Pharmacologic management of chronic neuropathic pain: Review of the Canadian Pain Society consensus statement. Can. Fam. Phys. 2017, 63, 844–852. [Google Scholar]
- Pinkernelle, J.; Fansa, H.; Ebmeyer, U.; Keilhoff, G. Prolonged Minocycline Treatment Impairs Motor Neuronal Survival and Glial Function in Organotypic Rat Spinal Cord Cultures. PLoS ONE 2013, 8, e73422. [Google Scholar] [CrossRef] [Green Version]
- Zarei, M.; Sabetkasaei, M.; Moini-Zanjani, T. Paradoxical effect of minocycline on established neuropathic pain in rat. EXCLI J. 2017, 16, 229–235. [Google Scholar]
- Verge, G.M.; Milligan, E.; Maier, S.F.; Watkins, L.R.; Naeve, G.S.; Foster, A.C. Fractalkine (CX3CL1) and fractalkine receptor (CX3CR1) distribution in spinal cord and dorsal root ganglia under basal and neuropathic pain conditions. Eur. J. Neurosci. 2004, 20, 1150–1160. [Google Scholar] [CrossRef]
- Zhang, E.; Yi, M.-H.; Ko, Y.; Kim, H.-W.; Seo, J.H.; Lee, Y.; Lee, W.; Kim, D.W. Expression of LC3 and Beclin 1 in the spinal dorsal horn following spinal nerve ligation-induced neuropathic pain. Brain Res. 2013, 1519, 31–39. [Google Scholar] [CrossRef]
- Chung, J.M.; Kim, H.K.; Chung, K. Segmental spinal nerve ligation model of neuropathic pain. In Pain Research; Springer: Berlin/Heidelberg, Germany, 2004; pp. 35–45. [Google Scholar]
- Pitcher, G.M.; Ritchie, J.; Henry, J.L. Paw withdrawal threshold in the von Frey hair test is influenced by the surface on which the rat stands. J. Neurosci. Methods 1999, 87, 185–193. [Google Scholar] [CrossRef]
- Hylden, J.L.; Wilcox, G.L. Intrathecal morphine in mice: A new technique. Eur. J. Pharmacol. 1980, 67, 313–316. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noh, C.; Shin, H.J.; Lee, S.; Kim, S.I.; Kim, Y.-H.; Lee, W.H.; Kim, D.W.; Lee, S.Y.; Ko, Y.K. CX3CR1-Targeted PLGA Nanoparticles Reduce Microglia Activation and Pain Behavior in Rats with Spinal Nerve Ligation. Int. J. Mol. Sci. 2020, 21, 3469. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103469
Noh C, Shin HJ, Lee S, Kim SI, Kim Y-H, Lee WH, Kim DW, Lee SY, Ko YK. CX3CR1-Targeted PLGA Nanoparticles Reduce Microglia Activation and Pain Behavior in Rats with Spinal Nerve Ligation. International Journal of Molecular Sciences. 2020; 21(10):3469. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103469
Chicago/Turabian StyleNoh, Chan, Hyo Jung Shin, Seounghun Lee, Song I Kim, Yoon-Hee Kim, Won Hyung Lee, Dong Woon Kim, Sun Yeul Lee, and Young Kwon Ko. 2020. "CX3CR1-Targeted PLGA Nanoparticles Reduce Microglia Activation and Pain Behavior in Rats with Spinal Nerve Ligation" International Journal of Molecular Sciences 21, no. 10: 3469. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103469