Increased Level of Vascular Endothelial Growth Factors by 4-hexylresorcinol is Mediated by Transforming Growth Factor-β1 and Accelerates Capillary Regeneration in the Burns in Diabetic Animals

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
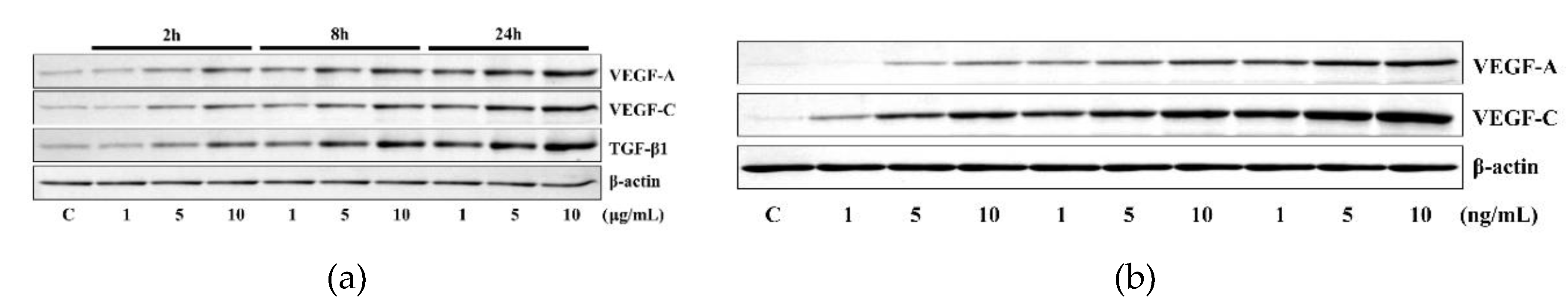
2. Results
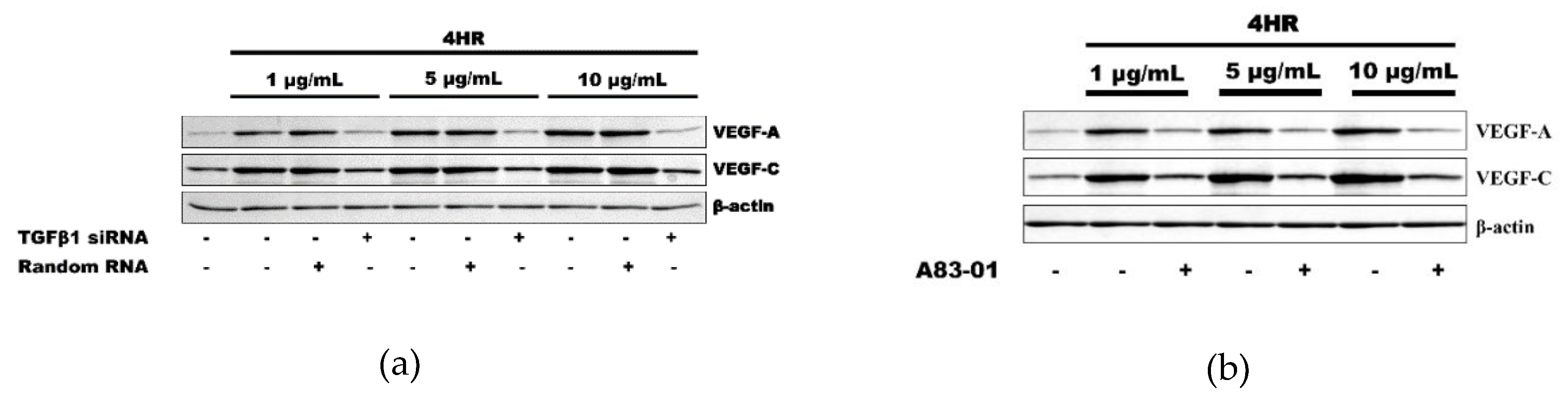
2.1. HR-Induced VEGF Expression in HUVECs Was Reduced by TGF-β1 siRNA
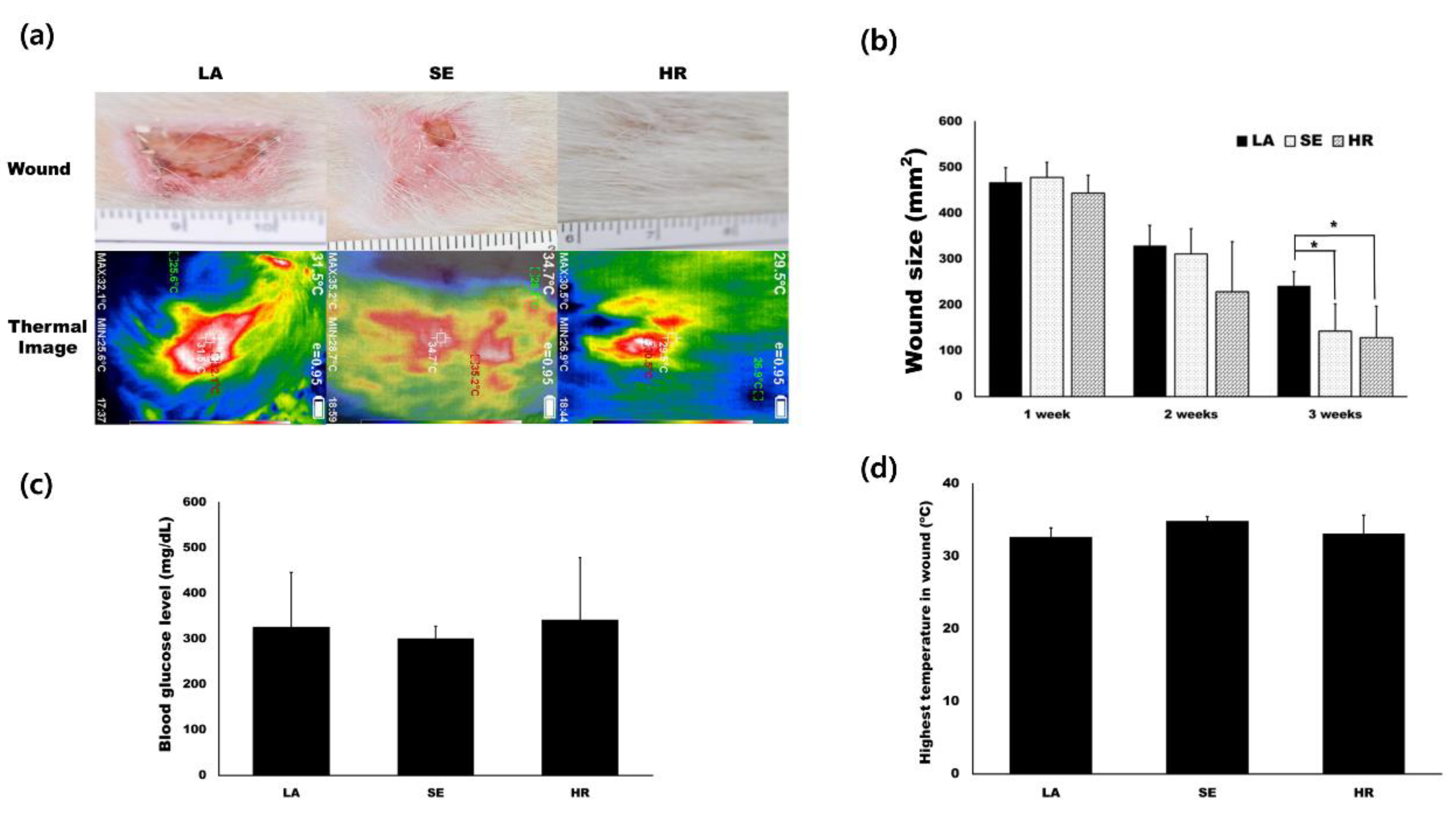
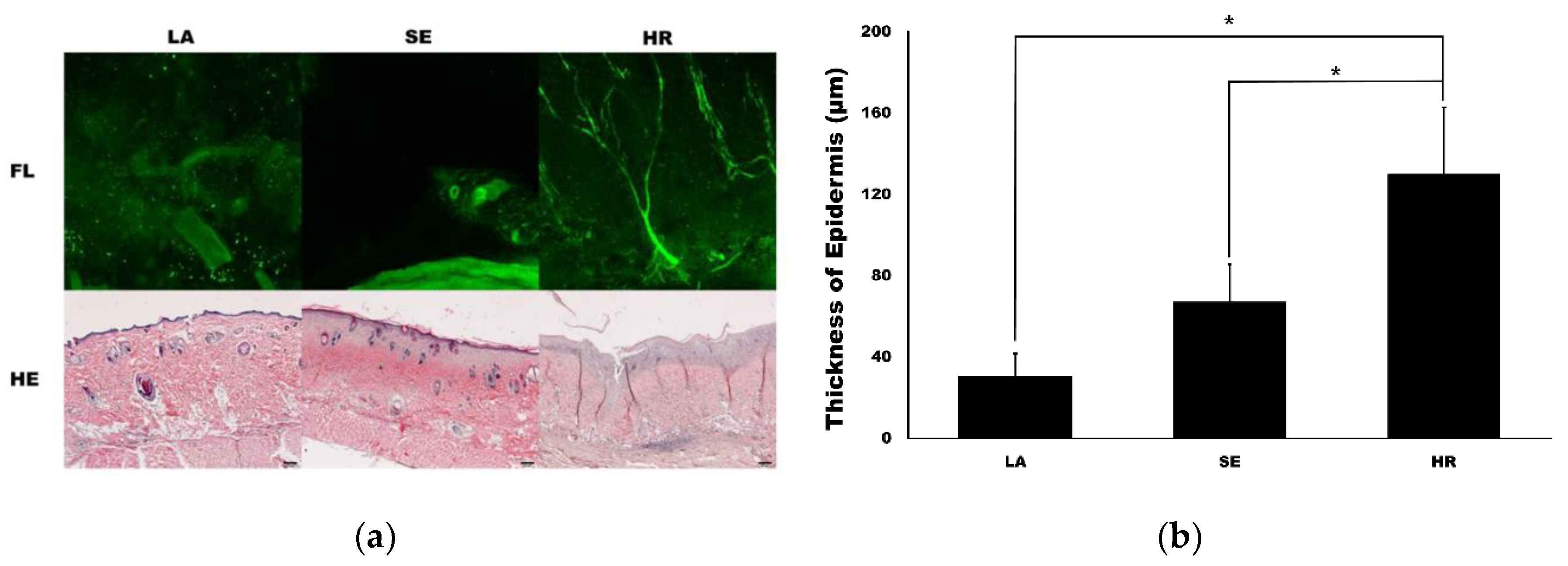
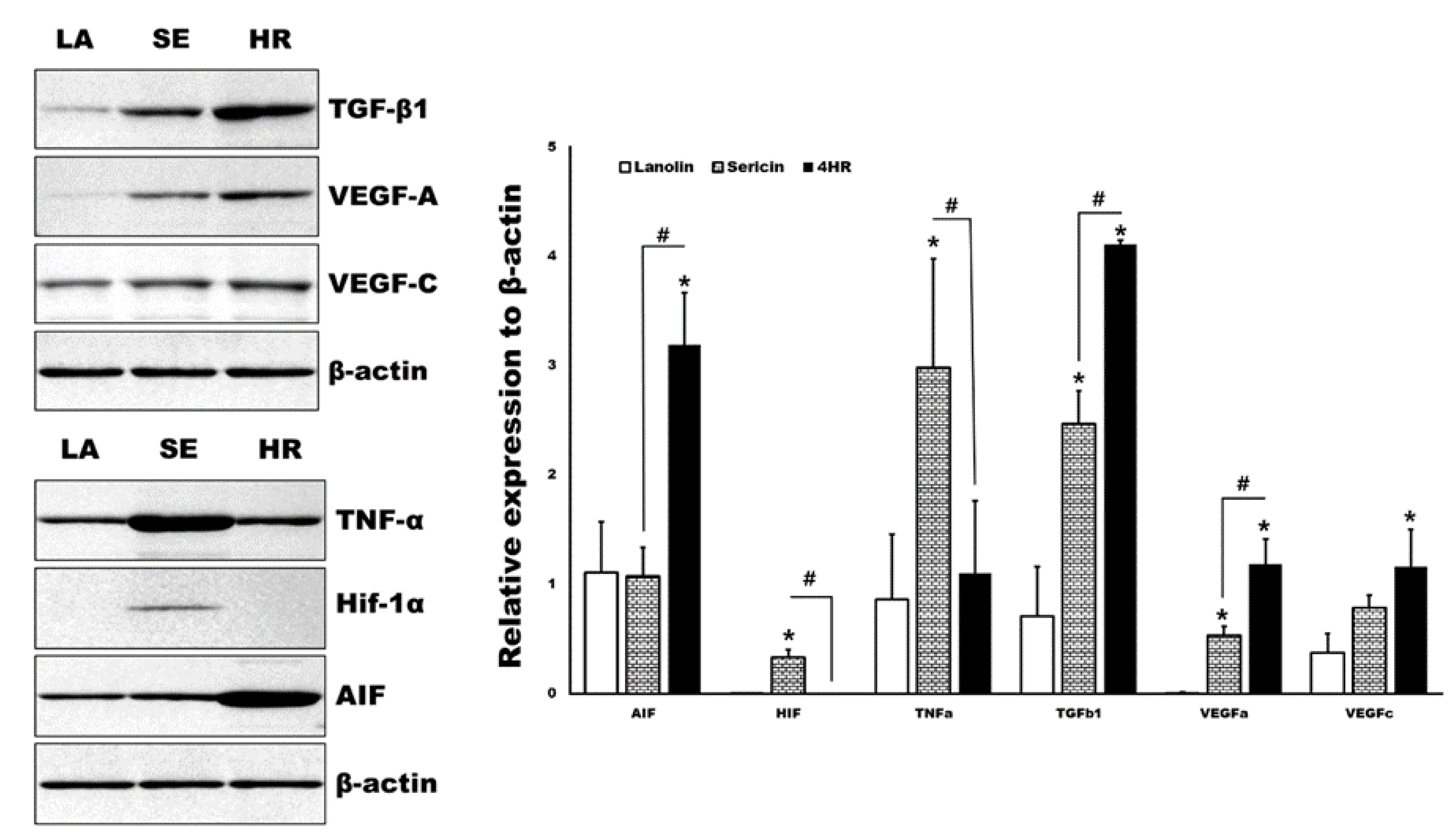
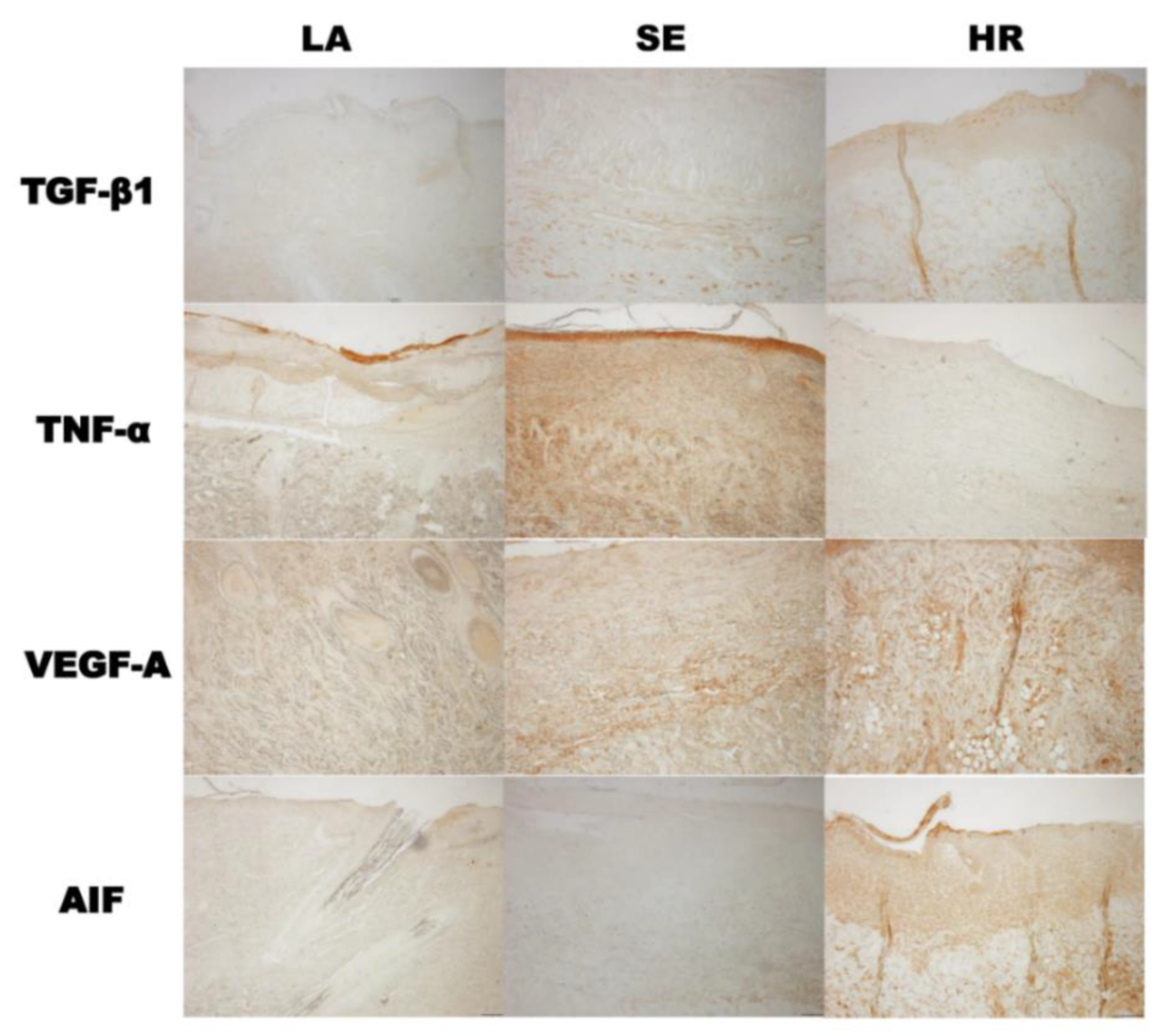
2.2. HR-Induced Capillary Regeneration in a Diabetic Rat Model
3. Discussion
4. Materials and Methods
4.1. HUVEC Culture
4.2. Western Blot and Inhibitory Assay
4.3. Animal Experiments
4.4. Tissue Transparent Procedure and Confocal Microscopic Examination
4.5. Histological Analysis and Western Blot for Tissue Samples
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Son, H.J.; Kim, J.W.; Kim, S.J. Pharmacoepidemiology and clinical characteristics of medication-related osteonecrosis of the jaw. Maxillofac. Plast. Reconstr. Surg. 2019, 41, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.M.; Lee, J.E.; Lee, S.J.; Lee, J.T.; Kwon, T.Y.; Kwon, T.G. Development of an experimental model for radiation-induced inhibition of cranial bone regeneration. Maxillofac. Plast. Reconstr. Surg. 2018, 40, 34. [Google Scholar] [CrossRef] [PubMed]
- Okonkwo, U.A.; DiPietro, L.A. Diabetes and wound angiogenesis. Int. J. Mol. Sci. 2017, 18, 1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alavi, A.; Sibbald, R.G.; Mayer, D.; Goodman, L.; Botros, M.; Armstrong, D.G.; Woo, K.; Boeni, T.; Ayello, E.A.; Kirsner, R.S. Diabetic foot ulcers: Part, I. Pathophysiology and prevention. J. Am. Acad. Dermatol. 2014, 70. [Google Scholar] [CrossRef] [PubMed]
- Jeffcoate, W.J.; Harding, K.G. Diabetic foot ulcers. Lancet 2003, 361, 1545–1551. [Google Scholar] [CrossRef]
- Goutos, I.; Nicholas, R.S.; Pandya, A.A.; Ghosh, S.J. Diabetes mellitus and burns. Part II-outcomes from burn injuries and future directions. Int. J. Burn Trauma 2015, 5, 13–21. [Google Scholar]
- Duke, J.M.; Randall, S.M.; Fear, M.W.; Boyd, J.H.; O’Halloran, E.; Rea, S.; Wood, F.M. Increased admissions for diabetes mellitus after burn. Burns 2016, 42, 1734–1739. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G. Immunomodulation for maxillofacial reconstructive surgery. Maxillofac. Plast. Reconstr. Surg. 2020, 42, 5. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Klein, T.; Murray, R.Z.; Crawford, R.; Chang, J.; Wu, C.; Xiao, Y. Osteoimmunomodulation for the development of advanced bone biomaterials. Mater. Today 2016, 19, 304–321. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.W.; Lee, S.J.; Kim, J.C. TNF-α upregulates HIF-1α expression in pterygium fibroblasts and enhances their susceptibility to VEGF independent of hypoxia. Exp. Eye Res. 2017, 164, 74–81. [Google Scholar] [CrossRef]
- Jo, Y.Y.; Kim, D.W.; Choi, J.Y.; Kim, S.G. 4-Hexylresorcinol and silk sericin increase the expression of vascular endothelial growth factor via different pathways. Sci. Rep. 2019, 9, 3448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiller, K.L.; Nassiri, S.; Witherel, C.E.; Anfang, R.R.; Ng, J.; Nakazawa, K.R.; Yu, T.; Vunjak-Novakovic, G. Sequential delivery of immunomodulatory cytokines to facilitate the M1-to-M2 transition of macrophages and enhance vascularization of bone scaffolds. Biomaterials 2015, 37, 194–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kariya, T.; Nishimura, H.; Mizuno, M.; Suzuki, Y.; Matsukawa, Y.; Sakata, F.; Maruyama, S.; Takei, Y.; Ito, Y. TGF-β1-VEGF-A pathway induces neoangiogenesis with peritoneal fibrosis in patients undergoing peritoneal dialysis. Am. J. Physiol. Renal Physiol. 2018, 314, F167–F180. [Google Scholar] [CrossRef] [PubMed]
- Zou, M.H.; Cohen, R.; Ullrich, V. Peroxynitrite and vascular endothelial dysfunction in diabetes mellitus. Endothelium 2004, 11, 89–97. [Google Scholar] [CrossRef]
- Ritchie, S.A.; Ewart, M.A.; Perry, C.G.; Connell, J.M.; Salt, I.P. The role of insulin and the adipocytokines in regulation of vascular endothelial function. Clin. Sci. 2004, 107, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Maresh, J.G.; Shohet, R.V. In vivo endothelial gene regulation in diabetes. Cardiovasc. Diabetol. 2008, 7, 8. [Google Scholar] [CrossRef] [Green Version]
- Joob, B.; Wiwanitkit, V. Silk sericin and burn wound. Arch. Dermatol. Res. 2013, 305, 555. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.K.; Yoon, C.S.; Kim, S.G.; Park, Y.W.; Lee, S.K. Effects of 4-hexylresorcinol on protein expressions in RAW 264.7 cells as determined by immunoprecipitation high performance liquid chromatography. Sci. Rep. 2019, 9, 3379. [Google Scholar] [CrossRef]
- Choi, K.H.; Kim, D.W.; Lee, S.K.; Kim, S.G.; Kim, T.W. The administration of 4-hexylresorcinol accelerates orthodontic tooth movement and increases the expression level of bone turnover markers in ovariectomized rats. Int. J. Mol. Sci. 2020, 21, 1526. [Google Scholar] [CrossRef] [Green Version]
- Tojo, M.; Hamashima, Y.; Hanyu, A.; Kajimoto, T.; Saitoh, M.; Miyazono, K.; Node, M.; Imamura, T. The ALK-5 inhibitor A-83-01 inhibits Smad signaling and epithelial-to-mesenchymal transition by transforming growth factor-beta. Cancer Sci. 2005, 96, 791–800. [Google Scholar] [CrossRef]
- Feng, S.; Song, X.H.; Zeng, C.M. Inhibition of amyloid fibrillation of lysozyme by phenolic compounds involves quinoprotein formation. FEBS Lett. 2012, 586, 3951–3955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.K.; Kim, S.G.; Lee, S.K. 4-Hexylresorcinol-induced angiogenesis potential in human endothelial cells. Maxillofac. Plast. Reconstr. Surg. 2020. accepted for publication. [Google Scholar]
- Ahn, J.; Kim, S.G.; Kim, M.K.; Kim, D.W.; Lee, J.H.; Seok, H.; Choi, J.Y. Topical delivery of 4-hexylresorcinol promotes wound healing via tumor necrosis factor-α suppression. Burns 2016, 42, 1534–1541. [Google Scholar] [CrossRef] [PubMed]
- Ranosz-Janicka, I.; Lis-Święty, A.; Skrzypek-Salamon, A.; Brzezińska-Wcisło, L. Detecting and quantifying activity/inflammation in localized scleroderma with thermal imaging. Skin Res. Technol. 2019, 25, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Eyuboglu, A.A.; Uysal, C.A.; Ozgun, G.; Coskun, E.; Markal Ertas, N.; Haberal, M. The effect of adipose derived stromal vascular fraction on stasis zone in an experimental burn model. Burns 2018, 44, 386–396. [Google Scholar] [CrossRef]
- Kim, S.G.; Kim, A.S.; Jeong, J.H.; Choi, J.Y.; Kweon, H. 4-hexylresorcinol stimulates the differentiation of SCC-9 cells through the suppression of E2F2, E2F3 and Sp3 expression and the promotion of Sp1 expression. Oncol. Rep. 2012, 28, 677–681. [Google Scholar] [CrossRef] [Green Version]
- Rochon, M.H.; Fradette, J.; Fortin, V.; Tomasetig, F.; Roberge, C.J.; Baker, K.; Berthod, F.; Auger, F.A.; Germain, L. Normal human epithelial cells regulate the size and morphology of tissue-engineered capillaries. Tissue Eng. Part A 2010, 16, 1457–1468. [Google Scholar] [CrossRef]
- Akçay, M.N.; Ozcan, O.; Gündoğdu, C.; Akçay, G.; Balik, A.; Köse, K.; Oren, D. Effect of nitric oxide synthase inhibitor on experimentally induced burn wounds. J. Trauma 2000, 49, 327–330. [Google Scholar] [CrossRef]
- Wang, X.; Niu, X.; Cheng, D. TGF-beta 1 gene expression in the healing process of skin wound in rat. Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi 1997, 11, 257–260. [Google Scholar]
- Amadasi, A.; Mozzarelli, A.; Meda, C.; Maggi, A.; Cozzini, P. Identification of xenoestrogens in food additives by an integrated in silico and in vitro approach. Chem. Res. Toxicol. 2009, 22, 52–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trenti, A.; Tedesco, S.; Boscaro, C.; Trevisi, L.; Bolego, C.; Cignarella, A. Estrogen, angiogenesis, immunity and cell metabolism: Solving the puzzle. Int. J. Mol. Sci. 2018, 19, 859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.J.; Oh, J.H.; Seok, H.; Jo, Y.Y.; Kim, D.W.; Garagiola, U.; Choi, J.Y.; Kim, S.G. 4-Hexylresorcinol exhibits different characteristics to estrogen. Appl. Sci. 2020, 10, 1737. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-W.; Jo, Y.-Y.; Garagiola, U.; Choi, J.-Y.; Kang, Y.-J.; Oh, J.-H.; Kim, S.-G. Increased Level of Vascular Endothelial Growth Factors by 4-hexylresorcinol is Mediated by Transforming Growth Factor-β1 and Accelerates Capillary Regeneration in the Burns in Diabetic Animals. Int. J. Mol. Sci. 2020, 21, 3473. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103473
Kim D-W, Jo Y-Y, Garagiola U, Choi J-Y, Kang Y-J, Oh J-H, Kim S-G. Increased Level of Vascular Endothelial Growth Factors by 4-hexylresorcinol is Mediated by Transforming Growth Factor-β1 and Accelerates Capillary Regeneration in the Burns in Diabetic Animals. International Journal of Molecular Sciences. 2020; 21(10):3473. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103473
Chicago/Turabian StyleKim, Dae-Won, You-Young Jo, Umberto Garagiola, Je-Yong Choi, Yei-Jin Kang, Ji-Hyeon Oh, and Seong-Gon Kim. 2020. "Increased Level of Vascular Endothelial Growth Factors by 4-hexylresorcinol is Mediated by Transforming Growth Factor-β1 and Accelerates Capillary Regeneration in the Burns in Diabetic Animals" International Journal of Molecular Sciences 21, no. 10: 3473. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103473