Emerging Roles of Estrogen-Regulated Enhancer and Long Non-Coding RNAs

 ,
, 
Abstract
:1. Introduction
2. Estrogen Signaling
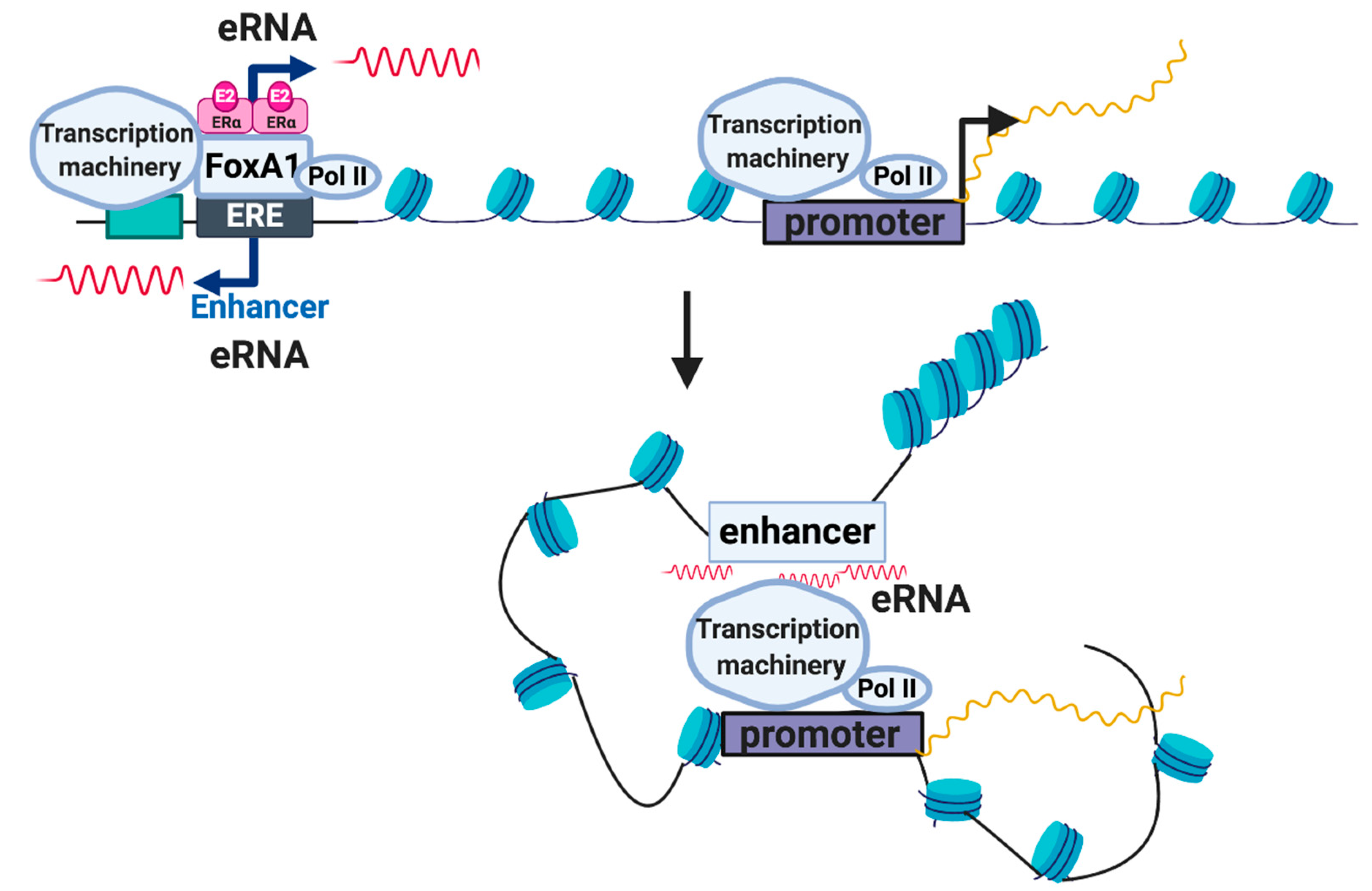
3. Enhancer RNAs (eRNAs)
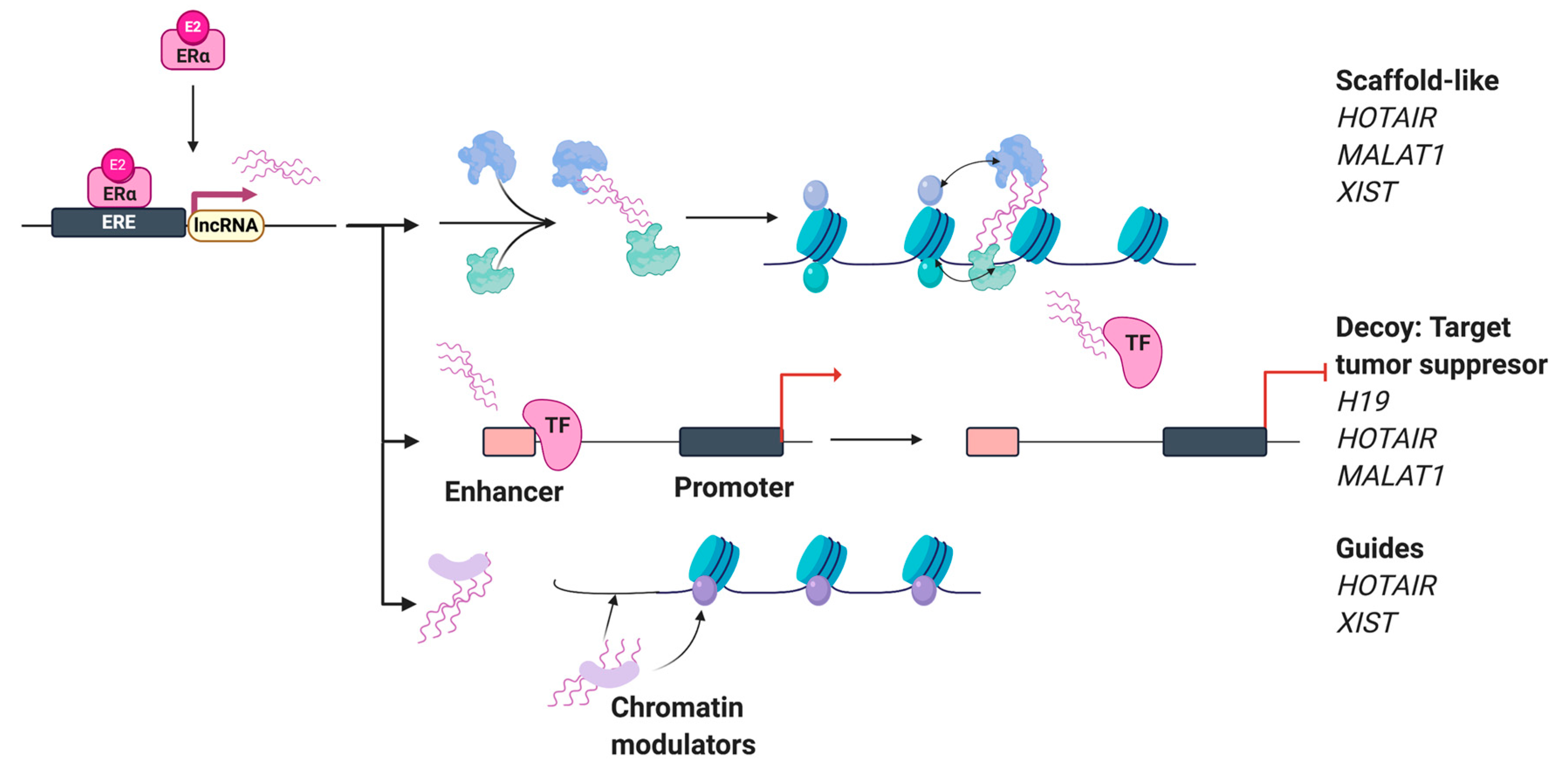
4. Estrogen-Regulated Long Non-Coding RNAs (lncRNAs)
4.1. HOX Transcript Antisense RNA (HOTAIR) and Metastasis-Associated Lung Adenocarcinoma Transcript 1 (MALAT1)
4.2. Myocardial Infarction-Associated Transcript (MIAT)
4.3. DSCAM Antisense RNA 1 (DSCAM-AS1)
4.4. Long Intergenic Non-Protein Coding RNA 472 (LINC00472) and Long Intergenic Non-Protein Coding RNA (LINC01016)
4.5. Eosinophil Granule Ontogeny Transcript (EGOT)
4.6. Long Intergenic Non-Protein Coding RNA, Regulator of Reprogramming (LINC-ROR), and LncRNA In Non-Homologous End Joining (NHEJ) Pathway 1 (LINP1)
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| mRNA | Messenger RNA |
| rRNA | Ribosomal RNA |
| tRNA | Transfer RNA |
| GRO-seq | Global nuclear Run-On Sequencing |
| PRO-seq | Precision nuclear Run-on Sequencing |
| ChIP-seq | Chromatin Immunoprecipitation Sequencing |
| ncRNA | Non-coding RNA |
| LncRNA | Long Non-coding RNA |
| eRNA | Enhancer RNA |
| ERα | Estrogen Receptor α |
| T-UCRs | Transcribed Ultra-Conserved Regions |
| ER+ | ER-positive |
| LncRNA152 | Long Non-coding RNA 152 |
| LncRNA 67 | Long Non-coding RNA 67 |
| H19 | H19 Imprinted Maternally Expressed Transcript |
| ceRNA | Competing Endogenous RNA |
| HOTAIR | HOX Transcript Antisense RNA |
| H3K27me3 | H3 lysine 27 trimethylation |
| PRC2 | Polycomb Repressive Complex 2 |
| PRC1 | Polycomb Repressive Complex 1 |
| MIAT | Myocardial Infarction-Associated Transcript |
| DSCAM-AS1 | DSCAM Antisense RNA 1 |
| NF-kB | Nuclear Factor kappa-light-chain-enhancer of activated B cells |
| LINC00472 | Long Intergenic Non-Protein Coding RNA 472 |
| LINC01016 | Long Intergenic Non-Protein Coding RNA 1016 |
| MALAT1 | Metastasis-Associated Lung Adenocarcinoma Transcript 1 |
| EGOT | Eosinophil Granule Ontogeny Transcript |
| LINC-ROR | Long Intergenic Non-Protein Coding RNA, Regulator of Reprogramming |
| LINP1 | LncRNA In Non-homologous End Joining (NHEJ) Pathway 1 |
References
- Camacho, C.V.; Choudhari, R.; Gadad, S.S. Long noncoding RNAs and cancer, an overview. Steroids 2018, 133, 93–95. [Google Scholar] [CrossRef] [PubMed]
- Choudhari, R.; Sedano, M.J.; Harrison, A.L.; Subramani, R.; Lin, K.Y.; Ramos, E.I.; Lakshmanaswamy, R.; Gadad, S.S. Long noncoding RNAs in cancer: From discovery to therapeutic targets. Adv. Clin. Chem. 2020, 95, 105–147. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Non-coding RNA genes and the modern RNA world. Nat. Rev. Genet. 2001, 2, 919–929. [Google Scholar] [CrossRef] [PubMed]
- He, R.Z.; Luo, D.X.; Mo, Y.Y. Emerging roles of lncRNAs in the post-transcriptional regulation in cancer. Genes Dis. 2019, 6, 6–15. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Hah, N.; Murakami, S.; Nagari, A.; Danko, C.G.; Kraus, W.L. Enhancer transcripts mark active estrogen receptor binding sites. Genome Res. 2013, 23, 1210–1223. [Google Scholar] [CrossRef] [Green Version]
- Shibayama, Y.; Fanucchi, S.; Mhlanga, M.M. Visualization of Enhancer-Derived Noncoding RNA. Methods Mol. Biol. 2017, 1468, 19–32. [Google Scholar] [CrossRef]
- Lee, T.I.; Johnstone, S.E.; Young, R.A. Chromatin immunoprecipitation and microarray-based analysis of protein location. Nat. Protoc. 2006, 1, 729–748. [Google Scholar] [CrossRef]
- Vasquez, Y.M.; Nandu, T.S.; Kelleher, A.M.; Ramos, E.I.; Gadad, S.S.; Kraus, W.L. Genome-wide analysis and functional prediction of the estrogen-regulated transcriptional response in the mouse uterusdagger. Biol. Reprod. 2020, 102, 327–338. [Google Scholar] [CrossRef]
- Yasar, P.; Ayaz, G.; User, S.D.; Gupur, G.; Muyan, M. Molecular mechanism of estrogen-estrogen receptor signaling. Reprod. Med. Biol. 2017, 16, 4–20. [Google Scholar] [CrossRef]
- Ponglikitmongkol, M.; Green, S.; Chambon, P. Genomic organization of the human oestrogen receptor gene. EMBO J. 1988, 7, 3385–3388. [Google Scholar] [CrossRef] [PubMed]
- Sand, P.; Luckhaus, C.; Schlurmann, K.; Gotz, M.; Deckert, J. Untangling the human estrogen receptor gene structure. J. Neural Transm. (Vienna) 2002, 109, 567–583. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Liu, R.; Perez, E.J.; Wen, Y.; Stevens, S.M.; Valencia, T.; Brun-Zinkernagel, A.M.; Prokai, L.; Will, Y.; Dykens, J. Mitochondrial localization of estrogen receptor beta. Proc. Natl. Acad. Sci. USA 2004, 101, 4130–4135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, A.M.; Flamini, M.I.; Baldacci, C.; Goglia, L.; Genazzani, A.R.; Simoncini, T. Estrogen receptor-alpha promotes breast cancer cell motility and invasion via focal adhesion kinase and N-WASP. Mol. Endocrinol. 2010, 24, 2114–2125. [Google Scholar] [CrossRef]
- Pedram, A.; Razandi, M.; Wallace, D.C.; Levin, E.R. Functional estrogen receptors in the mitochondria of breast cancer cells. Mol. Biol. Cell 2006, 17, 2125–2137. [Google Scholar] [CrossRef] [Green Version]
- Mahmoodzadeh, S.; Dworatzek, E. The Role of 17beta-Estradiol and Estrogen Receptors in Regulation of Ca(2+) Channels and Mitochondrial Function in Cardiomyocytes. Front. Endocrinol. (Lausanne) 2019, 10, 310. [Google Scholar] [CrossRef] [Green Version]
- Lazennec, G.; Thomas, J.A.; Katzenellenbogen, B.S. Involvement of cyclic AMP response element binding protein (CREB) and estrogen receptor phosphorylation in the synergistic activation of the estrogen receptor by estradiol and protein kinase activators. J. Steroid Biochem. Mol. Biol. 2001, 77, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Ray, A.; Prefontaine, K.E.; Ray, P. Down-modulation of interleukin-6 gene expression by 17 beta-estradiol in the absence of high affinity DNA binding by the estrogen receptor. J. Biol. Chem. 1994, 269, 12940–12946. [Google Scholar]
- Zhou, Y.; Eppenberger-Castori, S.; Eppenberger, U.; Benz, C.C. The NFkappaB pathway and endocrine-resistant breast cancer. Endocr. Relat. Cancer 2005, 12 (Suppl. 1), S37–S46. [Google Scholar] [CrossRef]
- Giuliano, M.; Trivedi, M.V.; Schiff, R. Bidirectional Crosstalk between the Estrogen Receptor and Human Epidermal Growth Factor Receptor 2 Signaling Pathways in Breast Cancer: Molecular Basis and Clinical Implications. Breast Care (Basel) 2013, 8, 256–262. [Google Scholar] [CrossRef] [Green Version]
- Belandia, B.; Orford, R.L.; Hurst, H.C.; Parker, M.G. Targeting of SWI/SNF chromatin remodelling complexes to estrogen-responsive genes. EMBO J. 2002, 21, 4094–4103. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.; Orom, U.A.; Cesaroni, M.; Beringer, M.; Taatjes, D.J.; Blobel, G.A.; Shiekhattar, R. Activating RNAs associate with Mediator to enhance chromatin architecture and transcription. Nature 2013, 494, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Vasquez, Y.M.; Kraus, W.L. The Estrogen-Regulated Transcriptome: Rapid, Robust, Extensive, and Transient; Humana Press: Cham, Switzerland, 2019; pp. 95–127. [Google Scholar] [CrossRef]
- Ding, M.; Liu, Y.; Liao, X.; Zhan, H.; Liu, Y.; Huang, W. Enhancer RNAs (eRNAs): New Insights into Gene Transcription and Disease Treatment. J. Cancer 2018, 9, 2334–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F. Enhancer-derived RNA: A Primer. Genomics Proteomics Bioinf. 2017, 15, 196–200. [Google Scholar] [CrossRef]
- Kim, T.K.; Hemberg, M.; Gray, J.M. Enhancer RNAs: A class of long noncoding RNAs synthesized at enhancers. Cold Spring Harb Perspect. Biol. 2015, 7, a018622. [Google Scholar] [CrossRef] [Green Version]
- Paralkar, V.R.; Taborda, C.C.; Huang, P.; Yao, Y.; Kossenkov, A.V.; Prasad, R.; Luan, J.; Davies, J.O.; Hughes, J.R.; Hardison, R.C.; et al. Unlinking an lncRNA from Its Associated cis Element. Mol. Cell 2016, 62, 104–110. [Google Scholar] [CrossRef] [Green Version]
- Stees, J.S.; Varn, F.; Huang, S.; Strouboulis, J.; Bungert, J. Recruitment of transcription complexes to enhancers and the role of enhancer transcription. Biology (Basel) 2012, 1, 778–793. [Google Scholar] [CrossRef]
- Li, W.; Notani, D.; Ma, Q.; Tanasa, B.; Nunez, E.; Chen, A.Y.; Merkurjev, D.; Zhang, J.; Ohgi, K.; Song, X. Functional roles of enhancer RNAs for oestrogen-dependent transcriptional activation. Nature 2013, 498, 516–520. [Google Scholar] [CrossRef]
- Rui, X.; Li, Y.; Jin, F.; Li, F. TMPRSS3 is a novel poor prognostic factor for breast cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 5435–5442. [Google Scholar]
- Ko, J.Y.; Oh, S.; Yoo, K.H. Functional Enhancers As Master Regulators of Tissue-Specific Gene Regulation and Cancer Development. Mol. Cells 2017, 40, 169–177. [Google Scholar] [CrossRef]
- Lee, H.; Zhang, Z.; Krause, H.M. Long Noncoding RNAs and Repetitive Elements: Junk or Intimate Evolutionary Partners? Trends Genet. 2019, 35, 892–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Prensner, J.R.; Chinnaiyan, A.M. The emergence of lncRNAs in cancer biology. Cancer Discov. 2011, 1, 391–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Gadad, S.S.; Kim, D.S.; Kraus, W.L. Discovery, Annotation, and Functional Analysis of Long Noncoding RNAs Controlling Cell-Cycle Gene Expression and Proliferation in Breast Cancer Cells. Mol. Cell 2015, 59, 698–711. [Google Scholar] [CrossRef] [Green Version]
- Choudhari, R.; Yang, B.; Rotwein, P.; Gadad, S.S. Structure and expression of the long noncoding RNA gene MIR503 in humans and non-human primates. Mol. Cell. Endocrinol. 2020, 510, 110819. [Google Scholar] [CrossRef]
- Collette, J.; Le Bourhis, X.; Adriaenssens, E. Regulation of Human Breast Cancer by the Long Non-Coding RNA H19. Int. J. Mol. Sci. 2017, 18, 2319. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Sun, J.; Yang, F. The role of long non-coding RNA H19 in breast cancer. Oncol. Lett. 2020, 19, 7–16. [Google Scholar] [CrossRef]
- Basak, P.; Chatterjee, S.; Weger, S.; Bruce, M.C.; Murphy, L.C.; Raouf, A. Estrogen regulates luminal progenitor cell differentiation through H19 gene expression. Endocr. Relat. Cancer 2015, 22, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Basak, P.; Chatterjee, S.; Bhat, V.; Su, A.; Jin, H.; Lee-Wing, V.; Liu, Q.; Hu, P.; Murphy, L.C.; Raouf, A. Long Non-Coding RNA H19 Acts as an Estrogen Receptor Modulator that is Required for Endocrine Therapy Resistance in ER+ Breast Cancer Cells. Cell. Physiol. Biochem. 2018, 51, 1518–1532. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, B.; Wu, X.; Huang, Q.; Chen, W.; Zhu, H.; Qu, X.; Xie, L.; Ma, X.; Huang, G. Long non-coding RNA MIAT is estrogen-responsive and promotes estrogen-induced proliferation in ER-positive breast cancer cells. Biochem. Biophys. Res. Commun. 2018, 503, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Niknafs, Y.S.; Han, S.; Ma, T.; Speers, C.; Zhang, C.; Wilder-Romans, K.; Iyer, M.K.; Pitchiaya, S.; Malik, R.; Hosono, Y. The lncRNA landscape of breast cancer reveals a role for DSCAM-AS1 in breast cancer progression. Nat. Commun. 2016, 7, 12791. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Shi, K.; Cheng, X.; Han, Y.; Li, Y.; Yu, D.; Liu, Z. Long Noncoding RNA LINC00472 Inhibits Proliferation and Promotes Apoptosis of Lung Adenocarcinoma Cells via Regulating miR-24-3p/ DEDD. Technol. Cancer Res. Treat. 2018, 17, 1533033818790490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonsson, P.; Coarfa, C.; Mesmar, F.; Raz, T.; Rajapakshe, K.; Thompson, J.F.; Gunaratne, P.H.; Williams, C. Single-Molecule Sequencing Reveals Estrogen-Regulated Clinically Relevant lncRNAs in Breast Cancer. Mol. Endocrinol. 2015, 29, 1634–1645. [Google Scholar] [CrossRef] [Green Version]
- Aiello, A.; Bacci, L.; Re, A.; Ripoli, C.; Pierconti, F.; Pinto, F.; Masetti, R.; Grassi, C.; Gaetano, C.; Bassi, P.F. MALAT1 and HOTAIR Long Non-Coding RNAs Play Opposite Role in Estrogen-Mediated Transcriptional Regulation in Prostate Cancer Cells. Sci. Rep. 2016, 6, 38414. [Google Scholar] [CrossRef]
- Yang, L.; Lin, C.; Liu, W.; Zhang, J.; Ohgi, K.A.; Grinstein, J.D.; Dorrestein, P.C.; Rosenfeld, M.G. ncRNA- and Pc2 methylation-dependent gene relocation between nuclear structures mediates gene activation programs. Cell 2011, 147, 773–788. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Wang, P.; Zhang, J.; Wu, H.; Sui, S.; Zhang, J.; Wang, Q.; Qiao, K.; Yang, W.; Xu, H. Ai-lncRNA EGOT enhancing autophagy sensitizes paclitaxel cytotoxicity via upregulation of ITPR1 expression by RNA-RNA and RNA-protein interactions in human cancer. Mol. Cancer 2019, 18, 89. [Google Scholar] [CrossRef]
- Peng, W.X.; Huang, J.G.; Yang, L.; Gong, A.H.; Mo, Y.Y. Linc-RoR promotes MAPK/ERK signaling and confers estrogen-independent growth of breast cancer. Mol. Cancer 2017, 16, 161. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Liang, Y.; Li, Y.; Song, X.; Zhang, N.; Li, X.; Chen, B.; Zhao, W.; Wang, L.; Yang, Q. LncRNA LINP1 confers tamoxifen resistance and negatively regulated by ER signaling in breast cancer. Cell Signal. 2020, 68, 109536. [Google Scholar] [CrossRef]
- Bhan, A.; Hussain, I.; Ansari, K.I.; Kasiri, S.; Bashyal, A.; Mandal, S.S. Antisense transcript long noncoding RNA (lncRNA) HOTAIR is transcriptionally induced by estradiol. J. Mol. Biol. 2013, 425, 3707–3722. [Google Scholar] [CrossRef] [Green Version]
- Sparmann, A.; van Lohuizen, M. Polycomb silencers control cell fate, development and cancer. Nat. Rev. Cancer 2006, 6, 846–856. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Qian, J.; Li, J.; Zhu, C. Knockdown of lncRNA-HOTAIR downregulates the drug-resistance of breast cancer cells to doxorubicin via the PI3K/AKT/mTOR signaling pathway. Exp. Ther. Med. 2019, 18, 435–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, J.A.; Davis, C.P.; Sunwoo, H.; Simon, M.D.; Sadreyev, R.I.; Wang, P.I.; Tolstorukov, M.Y.; Kingston, R.E. The long noncoding RNAs NEAT1 and MALAT1 bind active chromatin sites. Mol. Cell 2014, 55, 791–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vausort, M.; Wagner, D.R.; Devaux, Y. Long noncoding RNAs in patients with acute myocardial infarction. Circ. Res. 2014, 115, 668–677. [Google Scholar] [CrossRef] [Green Version]
- Jae, N.; Dimmeler, S. Long noncoding RNAs in diabetic retinopathy. Circ. Res. 2015, 116, 1104–1106. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.Q.; Hu, H.L.; Ye, N.; Shen, Y.; Xu, Q. Genetic variants in long non-coding RNA MIAT contribute to risk of paranoid schizophrenia in a Chinese Han population. Schizophr. Res. 2015, 166, 125–130. [Google Scholar] [CrossRef]
- Zhao, L.; Hu, K.; Cao, J.; Wang, P.; Li, J.; Zeng, K.; He, X.; Tu, P.F.; Tong, T.; Han, L. lncRNA miat functions as a ceRNA to upregulate sirt1 by sponging miR-22-3p in HCC cellular senescence. Aging (Albany NY) 2019, 11, 7098–7122. [Google Scholar] [CrossRef]
- Miano, V.; Ferrero, G.; Reineri, S.; Caizzi, L.; Annaratone, L.; Ricci, L.; Cutrupi, S.; Castellano, I.; Cordero, F.; De Bortoli, M. Luminal long non-coding RNAs regulated by estrogen receptor alpha in a ligand-independent manner show functional roles in breast cancer. Oncotarget 2016, 7, 3201–3216. [Google Scholar] [CrossRef] [Green Version]
- Hung, L.H.; Heiner, M.; Hui, J.; Schreiner, S.; Benes, V.; Bindereif, A. Diverse roles of hnRNP L in mammalian mRNA processing: A combined microarray and RNAi analysis. RNA 2008, 14, 284–296. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Katsaros, D.; Biglia, N.; Shen, Y.; Loo, L.; Yu, X.; Lin, H.; Fu, Y.; Chu, W.M.; Fei, P. ERalpha upregulates the expression of long non-coding RNA LINC00472 which suppresses the phosphorylation of NF-kappaB in breast cancer. Breast Cancer Res. Treat. 2019, 175, 353–368. [Google Scholar] [CrossRef]
- Xu, S.P.; Zhang, J.F.; Sui, S.Y.; Bai, N.X.; Gao, S.; Zhang, G.W.; Shi, Q.Y.; You, Z.L.; Zhan, C.; Pang, D. Downregulation of the long noncoding RNA EGOT correlates with malignant status and poor prognosis in breast cancer. Tumour Biol. 2015, 36, 9807–9812. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Quan, J.; Pan, X.; He, T.; Hu, J.; Li, Y.; Gui, Y.; Yang, S.; Mao, X.; Chen, Y. Identification of lncRNA EGOT as a tumor suppressor in renal cell carcinoma. Mol. Med. Rep. 2017, 16, 7072–7079. [Google Scholar] [CrossRef] [PubMed]
- Pedroza, D.A.; Chandel, V.; Kumar, D.; Doddapattar, P.; Biradar, M.S.; Lakshmanaswamy, R.; Gadad, S.S.; Choudhari, R. Role of Autophagy in Cancer Cell Metabolism. In Cancer Cell Metabolism: A Potential Target for Cancer Therapy; Kumar, D., Ed.; Springer Singapore: Singapore, 2020; pp. 65–87. [Google Scholar]
- Vrtacnik, P.; Ostanek, B.; Mencej-Bedrac, S.; Marc, J. The many faces of estrogen signaling. Biochem. Med. (Zagreb) 2014, 24, 329–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallou-Kabani, C.; Vige, A.; Gross, M.S.; Junien, C. Nutri-epigenomics: Lifelong remodelling of our epigenomes by nutritional and metabolic factors and beyond. Clin. Chem. Lab. Med. 2007, 45, 321–327. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| eRNA | Regulation by E2 | Function |
|---|---|---|
| TFF1e | Induced | Enhancer: promoter looping [29] |
| FOXC1e | Induced | Enhancer: promoter looping [29] |
| CA12e | Induced | Enhancer: promoter looping [29] |
| LncRNA | Regulation by E2 | Function |
|---|---|---|
| LncRNA152 | Induced | Proliferation [35] |
| LncRNA67 | Induced | Proliferation [35] |
| H19 | Induced | DNA methylation [38] |
| HOTAIR | Induced | Transcriptional regulation [41] |
| MIAT | Induced | Cell cycle progression [42] |
| DSCAM-AS1 | Induced | Cell cycle progression [43] |
| LINC00472 | Induced | Tumor suppressor [44] |
| LINC01016 | Induced | N/A [45] |
| MALAT1 | Repressed | Transcriptional regulation [46,47] |
| EGOT | Repressed | Autophagy pathway [48] |
| LINC-ROR | Repressed | Silences ER-signaling [49] |
| LINP1 | Repressed | G1-phase cell cycle arrest, apoptosis [50] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sedano, M.J.; Harrison, A.L.; Zilaie, M.; Das, C.; Choudhari, R.; Ramos, E.; Gadad, S.S. Emerging Roles of Estrogen-Regulated Enhancer and Long Non-Coding RNAs. Int. J. Mol. Sci. 2020, 21, 3711. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103711
Sedano MJ, Harrison AL, Zilaie M, Das C, Choudhari R, Ramos E, Gadad SS. Emerging Roles of Estrogen-Regulated Enhancer and Long Non-Coding RNAs. International Journal of Molecular Sciences. 2020; 21(10):3711. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103711
Chicago/Turabian StyleSedano, Melina J., Alana L. Harrison, Mina Zilaie, Chandrima Das, Ramesh Choudhari, Enrique Ramos, and Shrikanth S. Gadad. 2020. "Emerging Roles of Estrogen-Regulated Enhancer and Long Non-Coding RNAs" International Journal of Molecular Sciences 21, no. 10: 3711. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103711