In Vivo Transmigrated Human Neutrophils Are Highly Primed for Intracellular Radical Production Induced by Monosodium Urate Crystals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. MSU Crystals Trigger Intracellular ROS Production in Neutrophils
2.2. The Oxidative Response to MSU Is Dependent on the NADPH-Oxidase
2.3. The Oxidative Response to MSU Can Be Inhibited by Colchicine and Cytochalasin B
2.4. Increased MSU Responses after In Vivo Transmigration or In Vitro Priming
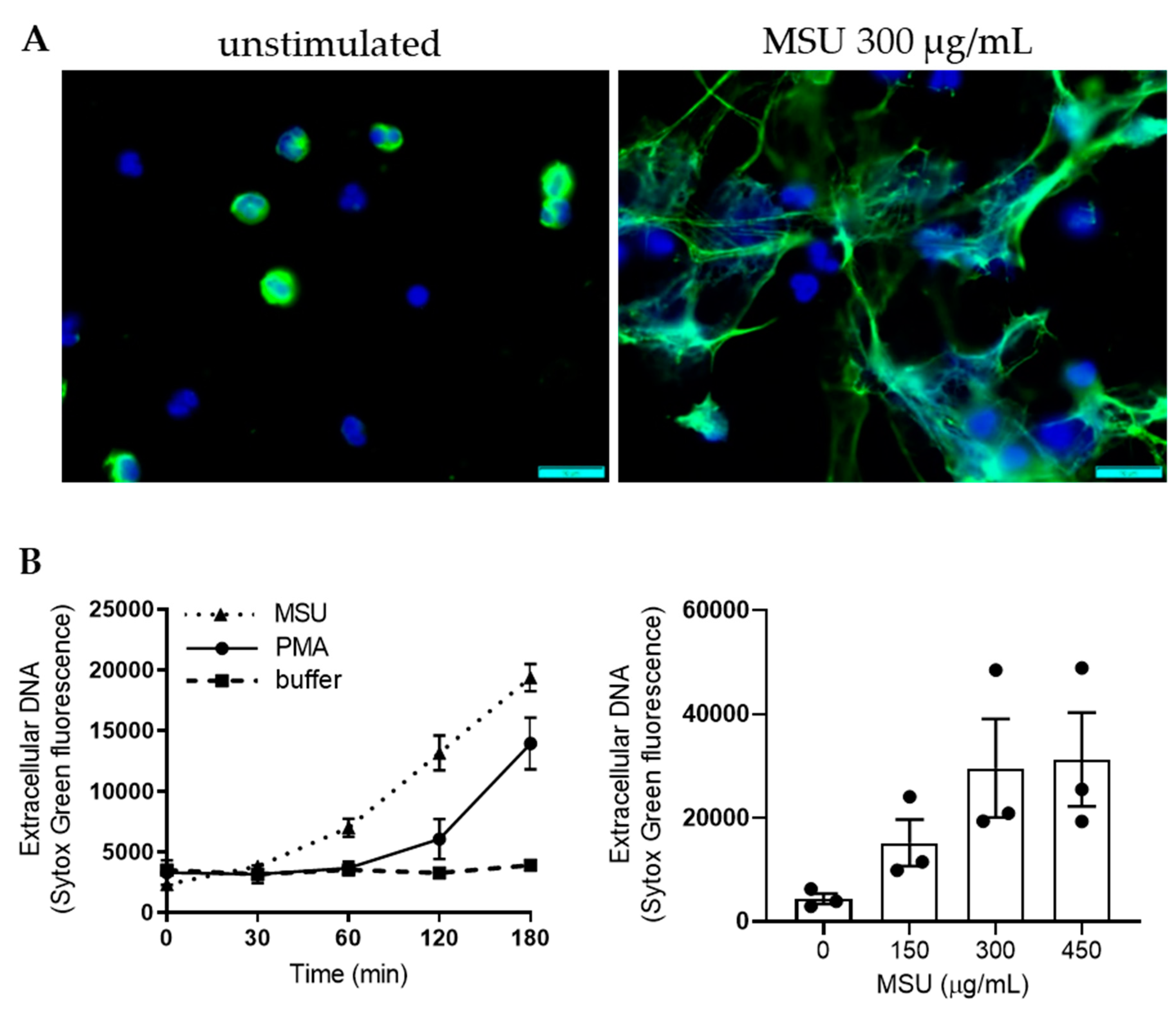
2.5. MSU Crystal Induced NET Formation is ROS Independent
2.6. In Vivo Transmigrated Neutrophils Are not Primed for NET Formation
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Study Subjects
4.3. Cell Preparation
4.4. ROS Measurements
4.4.1. Chemiluminescense (CL)
4.4.2. Extracellular H2O2 Production as Measured by Amplex Red
4.5. Quantification of NET Formation—Sytox Green Assay
4.6. Visualization of NET Formation
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kuo, C.F.; Grainge, M.J.; Zhang, W.; Doherty, M. Global epidemiology of gout: Prevalence, incidence and risk factors. Nat. Rev. Rheumatol. 2015, 11, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Petrilli, V.; Mayor, A.; Tardivel, A.; Tschopp, J. Gout-Associated uric acid crystals activate the NALP3 inflammasome. Nature 2006, 440, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitroulis, I.; Kambas, K.; Ritis, K. Neutrophils, IL-1beta, and gout: Is there a link? Semin Immunopathol. 2013, 35, 501–512. [Google Scholar] [CrossRef]
- Sil, P.; Wicklum, H.; Surell, C.; Rada, B. Macrophage-Derived IL-1beta enhances monosodium urate crystal-Triggered NET formation. Inflamm. Res. 2017, 66, 227–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarember, K.A.; Kuhns, D.B. Editorial: Will the real neutrophil please stand up? J. Leukoc Biol. 2011, 90, 1039–1041. [Google Scholar] [CrossRef] [PubMed]
- Nauseef, W.M. How human neutrophils kill and degrade microbes: An integrated view. Immunol. Rev. 2007, 219, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Bylund, J.; Brown, K.L.; Movitz, C.; Dahlgren, C.; Karlsson, A. Intracellular generation of superoxide by the phagocyte NADPH oxidase: How, where, and what for? Free Radic Biol. Med. 2010, 49, 1834–1845. [Google Scholar] [CrossRef]
- Dahlgren, C.; Karlsson, A.; Bylund, J. Intracellular Neutrophil Oxidants: From Laboratory Curiosity to Clinical Reality. J. Immunol. 2019, 202, 3127–3134. [Google Scholar] [CrossRef]
- Bylund, J.; Goldblatt, D.; Speert, D.P. Chronic granulomatous disease: From genetic defect to clinical presentation. Adv. Exp. Med. Biol. 2005, 568, 67–87. [Google Scholar] [CrossRef]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef]
- Mayadas, T.N.; Cullere, X.; Lowell, C.A. The multifaceted functions of neutrophils. Annu. Rev. Pathol. 2014, 9, 181–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeltz, S.; Amini, P.; Anders, H.J.; Andrade, F.; Bilyy, R.; Chatfield, S.; Cichon, I.; Clancy, D.M.; Desai, J.; Dumych, T.; et al. To NET or not to NET:current opinions and state of the science regarding the formation of neutrophil extracellular traps. Cell Death Differ. 2019, 26, 395–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rada, B. Neutrophil Extracellular Traps and Microcrystals. J. Immunol. Res. 2017, 2017, 2896380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Branzk, N.; Lubojemska, A.; Hardison, S.E.; Wang, Q.; Gutierrez, M.G.; Brown, G.D.; Papayannopoulos, V. Neutrophils sense microbe size and selectively release neutrophil extracellular traps in response to large pathogens. Nat. Immunol. 2014, 15, 1017–1025. [Google Scholar] [CrossRef] [Green Version]
- Behnen, M.; Leschczyk, C.; Moller, S.; Batel, T.; Klinger, M.; Solbach, W.; Laskay, T. Immobilized immune complexes induce neutrophil extracellular trap release by human neutrophil granulocytes via FcgammaRIIIB and Mac-1. J. Immunol. 2014, 193, 1954–1965. [Google Scholar] [CrossRef]
- Keshari, R.S.; Jyoti, A.; Dubey, M.; Kothari, N.; Kohli, M.; Bogra, J.; Barthwal, M.K.; Dikshit, M. Cytokines induced neutrophil extracellular traps formation: Implication for the inflammatory disease condition. PLoS ONE 2012, 7, e48111. [Google Scholar] [CrossRef] [Green Version]
- Sollberger, G.; Tilley, D.O.; Zychlinsky, A. Neutrophil Extracellular Traps: The Biology of Chromatin Externalization. Dev. Cell 2018, 44, 542–553. [Google Scholar] [CrossRef] [Green Version]
- Bjornsdottir, H.; Welin, A.; Dahlgren, C.; Karlsson, A.; Bylund, J. Quantification of heterotypic granule fusion in human neutrophils by imaging flow cytometry. Data Brief 2016, 6, 386–393. [Google Scholar] [CrossRef]
- Bjornsdottir, H.; Welin, A.; Michaelsson, E.; Osla, V.; Berg, S.; Christenson, K.; Sundqvist, M.; Dahlgren, C.; Karlsson, A.; Bylund, J. Neutrophil NET formation is regulated from the inside by myeloperoxidase-processed reactive oxygen species. Free Radic Biol. Med. 2015, 89, 1024–1035. [Google Scholar] [CrossRef]
- Abramson, S.; Hoffstein, S.T.; Weissmann, G. Superoxide anion generation by human neutrophils exposed to monosodium urate. Arthritis Rheum. 1982, 25, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Chatfield, S.M.; Grebe, K.; Whitehead, L.W.; Rogers, K.L.; Nebl, T.; Murphy, J.M.; Wicks, I.P. Monosodium Urate Crystals Generate Nuclease-Resistant Neutrophil Extracellular Traps via a Distinct Molecular Pathway. J. Immunol. 2018, 200, 1802–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudry, M.; Roberge, C.J.; de Medicis, R.; Lussier, A.; Poubelle, P.E.; Naccache, P.H. Crystal-Induced neutrophil activation. III. Inflammatory microcrystals induce a distinct pattern of tyrosine phosphorylation in human neutrophils. J. Clin. Invest. 1993, 91, 1649–1655. [Google Scholar] [CrossRef] [PubMed]
- Naccache, P.H.; Bourgoin, S.; Plante, E.; Roberge, C.J.; de Medicis, R.; Lussier, A.; Poubelle, P.E. Crystal-Induced neutrophil activation. II. Evidence for the activation of a phosphatidylcholine-specific phospholipase D. Arthritis Rheum. 1993, 36, 117–125. [Google Scholar] [CrossRef]
- Schauer, C.; Janko, C.; Munoz, L.E.; Zhao, Y.; Kienhofer, D.; Frey, B.; Lell, M.; Manger, B.; Rech, J.; Naschberger, E.; et al. Aggregated neutrophil extracellular traps limit inflammation by degrading cytokines and chemokines. Nat. Med. 2014, 20, 511–517. [Google Scholar] [CrossRef]
- Schorn, C.; Janko, C.; Krenn, V.; Zhao, Y.; Munoz, L.E.; Schett, G.; Herrmann, M. Bonding the foe-NETting neutrophils immobilize the pro-inflammatory monosodium urate crystals. Front. Immunol. 2012, 3, 376. [Google Scholar] [CrossRef] [Green Version]
- Mitroulis, I.; Kambas, K.; Chrysanthopoulou, A.; Skendros, P.; Apostolidou, E.; Kourtzelis, I.; Drosos, G.I.; Boumpas, D.T.; Ritis, K. Neutrophil extracellular trap formation is associated with IL-1beta and autophagy-Related signaling in gout. PLoS ONE 2011, 6, e29318. [Google Scholar] [CrossRef]
- Schorn, C.; Janko, C.; Latzko, M.; Chaurio, R.; Schett, G.; Herrmann, M. Monosodium urate crystals induce extracellular DNA traps in neutrophils, eosinophils, and basophils but not in mononuclear cells. Front. Immunol. 2012, 3, 277. [Google Scholar] [CrossRef] [Green Version]
- Bylund, J.; Bjornsdottir, H.; Sundqvist, M.; Karlsson, A.; Dahlgren, C. Measurement of respiratory burst products, released or retained, during activation of professional phagocytes. Methods Mol. Biol. 2014, 1124, 321–338. [Google Scholar] [CrossRef]
- Hirano, K.; Chen, W.S.; Chueng, A.L.; Dunne, A.A.; Seredenina, T.; Filippova, A.; Ramachandran, S.; Bridges, A.; Chaudry, L.; Pettman, G.; et al. Discovery of GSK2795039, a Novel Small Molecule NADPH Oxidase 2 Inhibitor. Antioxid Redox Signal 2015, 23, 358–374. [Google Scholar] [CrossRef] [Green Version]
- Buck, A.; Sanchez Klose, F.P.; Venkatakrishnan, V.; Khamzeh, A.; Dahlgren, C.; Christenson, K.; Bylund, J. DPI Selectively Inhibits Intracellular NADPH Oxidase Activity in Human Neutrophils. Immunohorizons 2019, 3, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Sengelov, H.; Follin, P.; Kjeldsen, L.; Lollike, K.; Dahlgren, C.; Borregaard, N. Mobilization of granules and secretory vesicles during in vivo exudation of human neutrophils. J. Immunol. 1995, 154, 4157–4165. [Google Scholar] [PubMed]
- Bjorkman, L.; Christenson, K.; Davidsson, L.; Martensson, J.; Amirbeagi, F.; Welin, A.; Forsman, H.; Karlsson, A.; Dahlgren, C.; Bylund, J. Neutrophil recruitment to inflamed joints can occur without cellular priming. J. Leukoc Biol. 2019, 105, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Bjornsdottir, H.; Dahlstrand Rudin, A.; Klose, F.P.; Elmwall, J.; Welin, A.; Stylianou, M.; Christenson, K.; Urban, C.F.; Forsman, H.; Dahlgren, C.; et al. Phenol-Soluble Modulin alpha Peptide Toxins from Aggressive Staphylococcus aureus Induce Rapid Formation of Neutrophil Extracellular Traps through a Reactive Oxygen Species-Independent Pathway. Front. Immunol. 2017, 8, 257. [Google Scholar] [CrossRef] [Green Version]
- Pascual, E. Persistence of monosodium urate crystals and low-grade inflammation in the synovial fluid of patients with untreated gout. Arthritis Rheum. 1991, 34, 141–145. [Google Scholar] [CrossRef]
- Bjorkman, L.; Dahlgren, C.; Karlsson, A.; Brown, K.L.; Bylund, J. Phagocyte-Derived reactive oxygen species as suppressors of inflammatory disease. Arthritis Rheum. 2008, 58, 2931–2935. [Google Scholar] [CrossRef]
- Sareila, O.; Kelkka, T.; Pizzolla, A.; Hultqvist, M.; Holmdahl, R. NOX2 complex-Derived ROS as immune regulators. Antioxid Redox Signal 2011, 15, 2197–2208. [Google Scholar] [CrossRef]
- Boyum, A.; Lovhaug, D.; Tresland, L.; Nordlie, E.M. Separation of leucocytes: Improved cell purity by fine adjustments of gradient medium density and osmolality. Scand J. Immunol. 1991, 34, 697–712. [Google Scholar] [CrossRef]
- Courtney, P.; Doherty, M. Joint aspiration and injection. Best Pract Res. Clin. Rheumatol. 2005, 19, 345–369. [Google Scholar] [CrossRef]
- Christenson, K.; Bjorkman, L.; Davidsson, L.; Karlsson, A.; Follin, P.; Dahlgren, C.; Bylund, J. Collection of in vivo transmigrated neutrophils from human skin. Methods Mol. Biol. 2014, 1124, 39–52. [Google Scholar] [CrossRef]
- Christenson, K.; Bjorkman, L.; Karlsson, J.; Sundqvist, M.; Movitz, C.; Speert, D.P.; Dahlgren, C.; Bylund, J. In vivo-Transmigrated human neutrophils are resistant to antiapoptotic stimulation. J. Leukoc Biol. 2011, 90, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, C.; Bjornsdottir, H.; Sundqvist, M.; Christenson, K.; Bylund, J. Measurement of Respiratory Burst Products, Released or Retained, During Activation of Professional Phagocytes. Methods Mol. Biol. 2020, 2087, 301–324. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davidsson, L.; Dahlstrand Rudin, A.; Sanchez Klose, F.P.; Buck, A.; Björkman, L.; Christenson, K.; Bylund, J. In Vivo Transmigrated Human Neutrophils Are Highly Primed for Intracellular Radical Production Induced by Monosodium Urate Crystals. Int. J. Mol. Sci. 2020, 21, 3750. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113750
Davidsson L, Dahlstrand Rudin A, Sanchez Klose FP, Buck A, Björkman L, Christenson K, Bylund J. In Vivo Transmigrated Human Neutrophils Are Highly Primed for Intracellular Radical Production Induced by Monosodium Urate Crystals. International Journal of Molecular Sciences. 2020; 21(11):3750. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113750
Chicago/Turabian StyleDavidsson, Lisa, Agnes Dahlstrand Rudin, Felix Peter Sanchez Klose, Alicia Buck, Lena Björkman, Karin Christenson, and Johan Bylund. 2020. "In Vivo Transmigrated Human Neutrophils Are Highly Primed for Intracellular Radical Production Induced by Monosodium Urate Crystals" International Journal of Molecular Sciences 21, no. 11: 3750. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113750