Immune Checkpoints Contribute Corneal Immune Privilege: Implications for Dry Eye Associated with Checkpoint Inhibitors

Abstract
:1. Introduction
2. Immune Suppressive Microenvironment in the Anterior Segment of the Eye
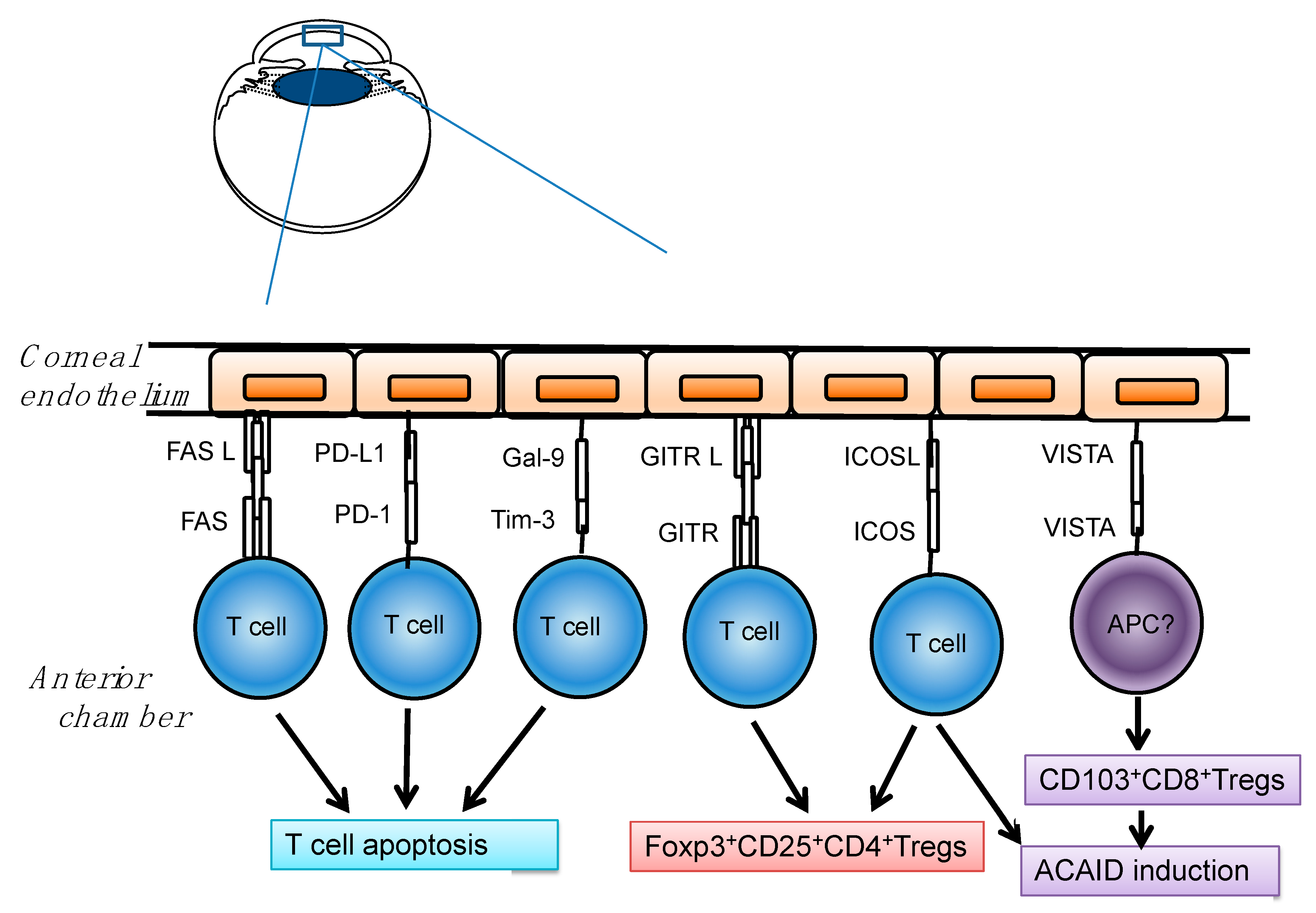
3. Role of Immune Checkpoints Molecules in the Cornea
3.1. Immune Checkpoints-Mediated T Cell Apoptosis in the Cornea
3.1.1. Fas Ligand and Fas
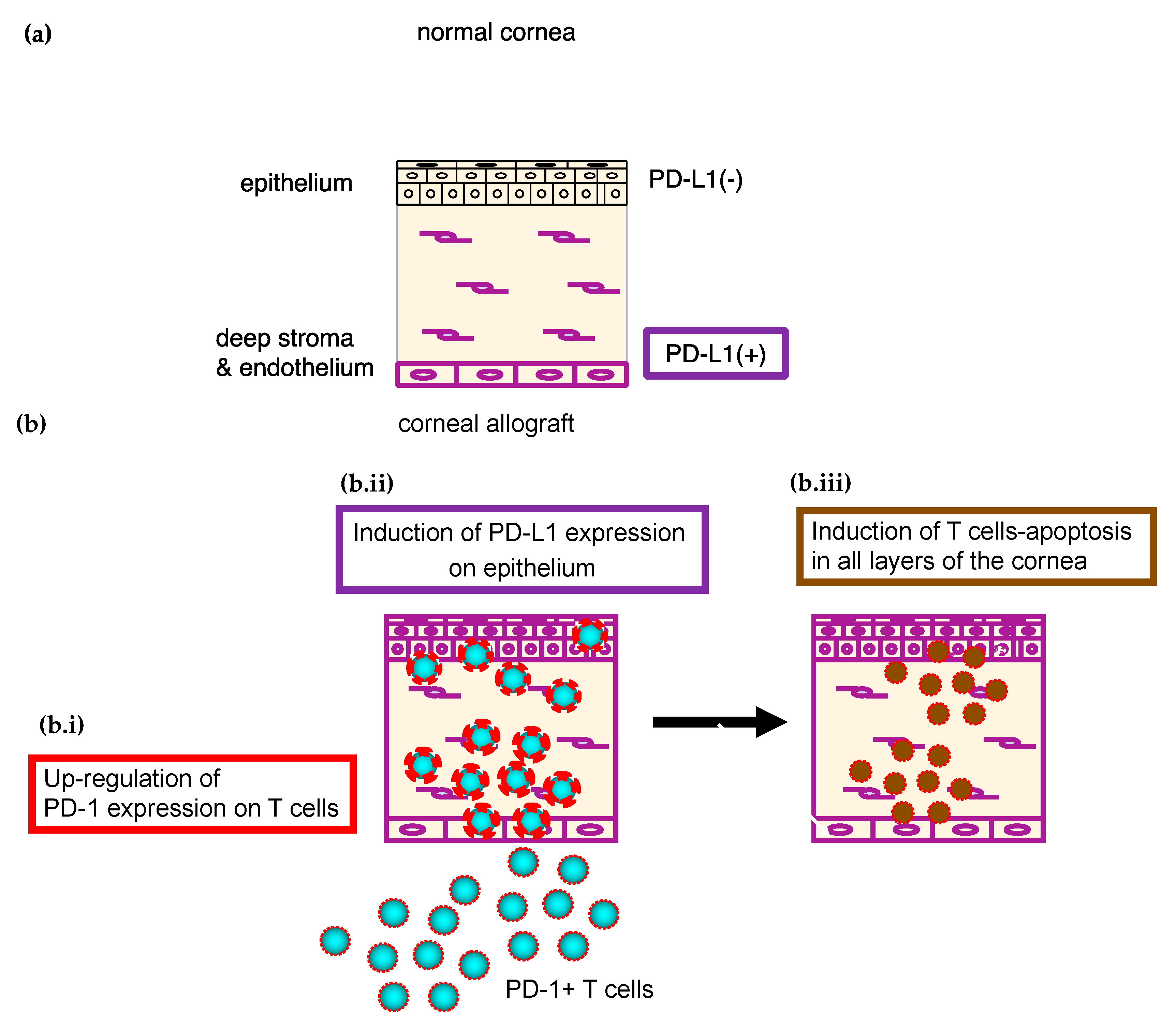
3.1.2. PD-Ligand 1 and PD-1
3.1.3. Galectin-9 and Tim-3
3.2. Immune Checkpoints-Mediated Treg and Peripheral Tolerance in the Cornea
3.2.1. ICOS Ligand and ICOS
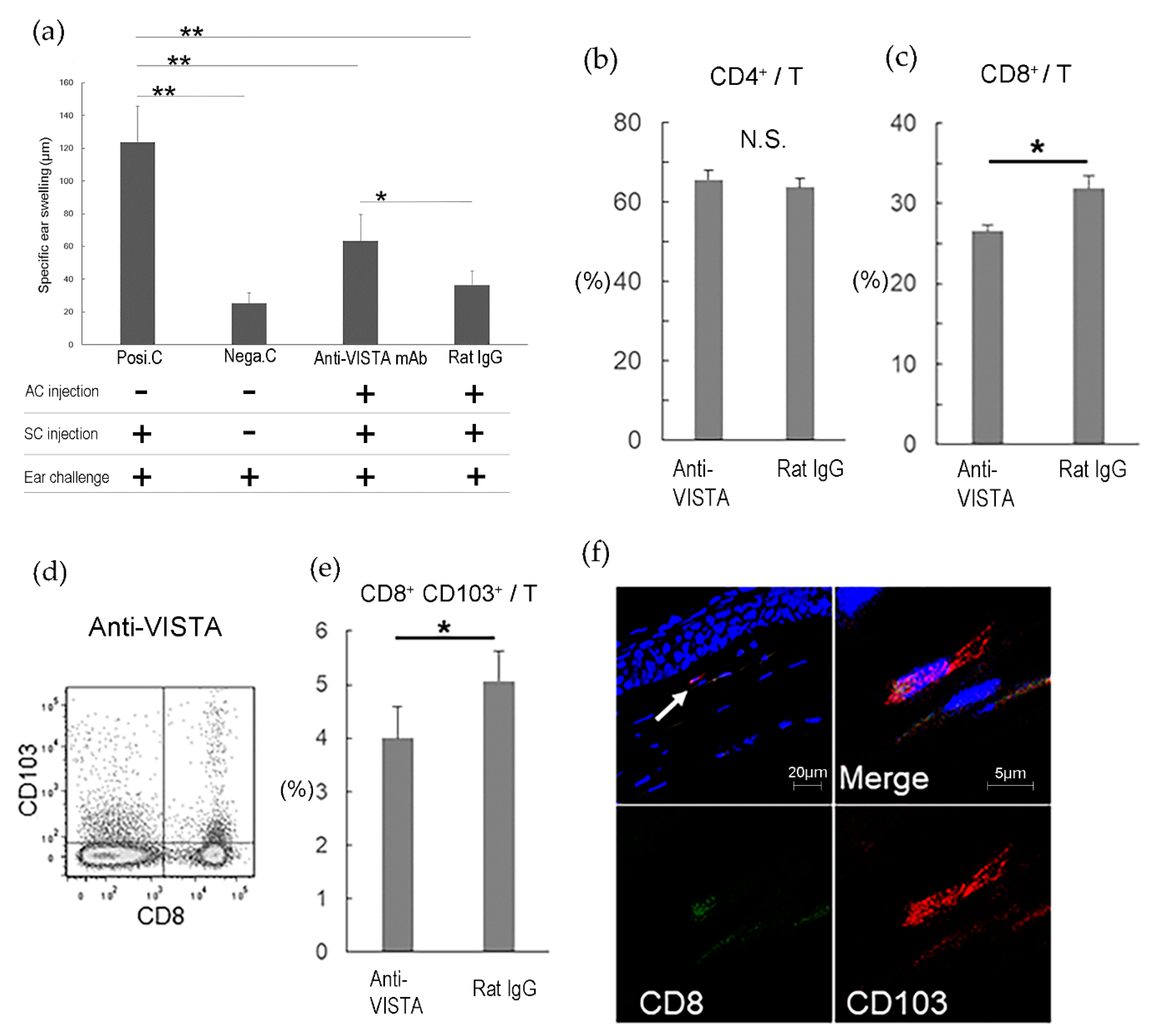
3.2.2. VISTA(PD-1H)
3.2.3. GITR Ligand and GITR
3.3. Other Molecules Contributing to Treg in the Cornea
4. Dry Eye as irAE Induced by Immune Checkpoint Inhibitors
4.1. Immune-Related Adverse Events (irAEs)
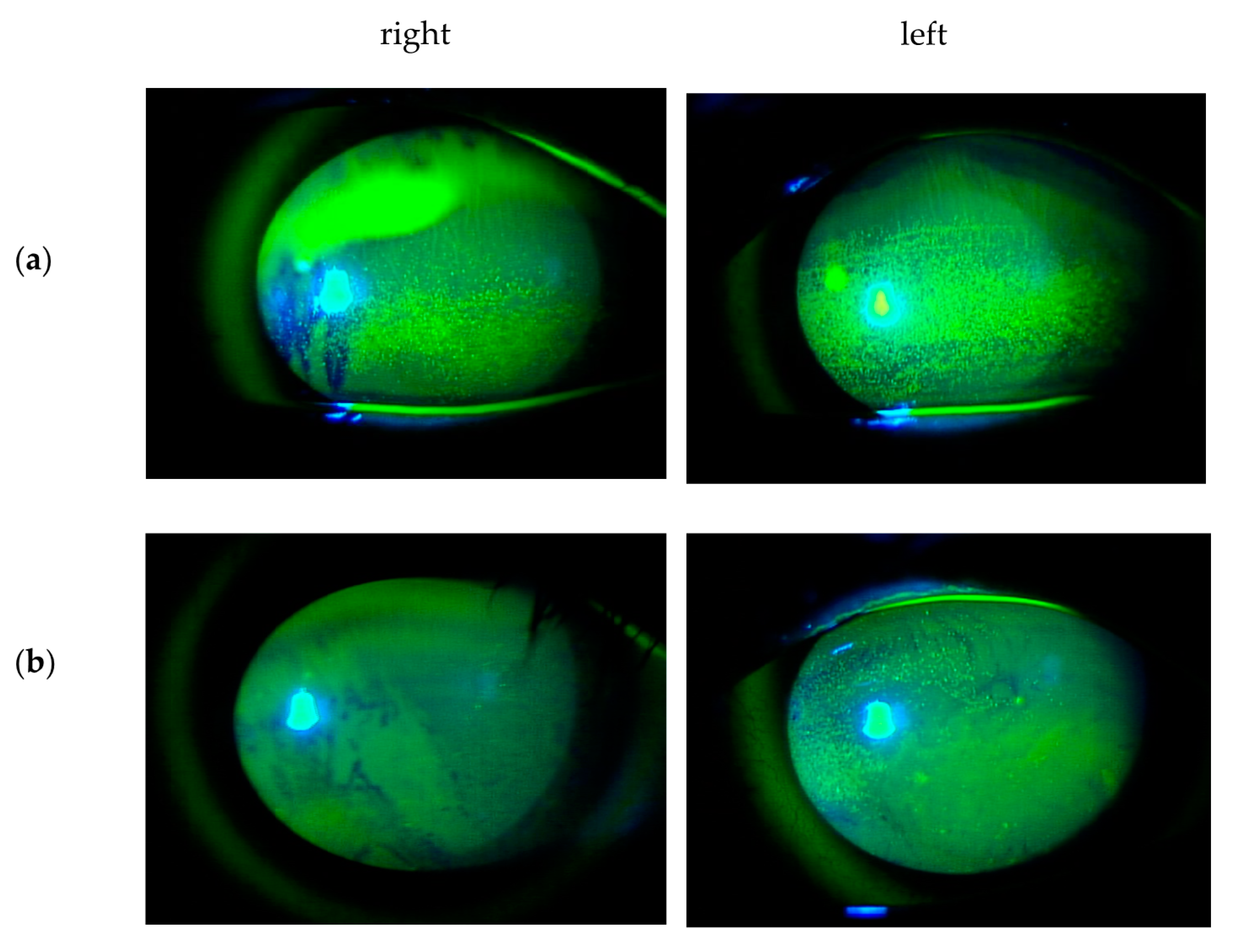
4.2. Immunopathologic Mechanisms of Dry Eye Associated with Checkpoint Inhibitor Therapy
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ACAID | anterior chamber-associated immune deviation |
| α-MSH | alpha-Melanocyte stimulating hormone |
| VIP | vasoactive intestinal peptide |
| CGRP | calcitonin gene-related peptide |
| TGF-β2 | transforming growth factor-beta 2 |
| TSP-1 | thrombospondin-1 |
| MIF | macrophage migrating inhibitory factor |
| IL-1Ra | interleukin 1 receptor antagonist |
| sFas L | soluble Fas ligand |
| PD-1 | programmed death-1 |
| PD-L1 | programmed death ligand-1 |
| Gal-9 | Galectin-9 |
| Tim-3 | T-cell immunoglobulin and mucin domain-3 |
| ICOS | inducible costimulatory molecule |
| ICOSL | inducible costimulatory molecule ligand |
| VISTA | V-domain Ig suppressor of T cell activation |
| GITR | glucocorticoid-induced tumor necrosis factor receptor family-related protein |
| GITRL | glucocorticoid-induced tumor necrosis factor receptor family-related protein ligand |
| TRAIL | tumor necrosis factor related apoptosis-inducing ligand |
| CTLA4 | cytotoxic T lymphocyte-associated antigen 4 |
| CTLA-2α | cytotoxic T lymphocyte-associated antigen-2 alpha |
| APCs | antigen presenting cells |
| mAb | monoclonal antibody |
| Treg | regulatory T cells |
| irAE | immune-related adverse event |
References
- Niederkorn, J.Y. See no evil, hear no evil, do no evil: The lessons of immune privilege. Nat. Immunol. 2006, 7, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Streilein, J.W. Ocular immune privilege: Therapeutic opportunities from an experiment of nature. Nat. Rev. Immunol. 2003, 3, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Hori, J.; Yamaguchi, T.; Keino, H.; Hamrah, P.; Maruyama, K. Immune privilege in corneal transplantation. Prog. Retin. Eye Res. 2019, 72, 100758. [Google Scholar] [CrossRef] [PubMed]
- Medawar, P.B. Immunity to homologous grafted skin; the fate of skin homografts transplanted to the brain, to subcutaneous tissue, and to the anterior chamber of the eye. Br. J. Exp. Pathol. 1948, 29, 58–69. [Google Scholar]
- Barker, C.F.; Billingham, R.E. Immunologically privileged sites. Adv. Immunol. 1977, 25, 1–54. [Google Scholar]
- Streilein, J.W. Immune regulation and the eye: A dangerous compromise. FASEB J. 1987, 1, 199–208. [Google Scholar] [CrossRef]
- Niederkorn, J.Y. Immune privilege and immune regulation in the eye. Adv. Immunol. 1990, 48, 191–226. [Google Scholar]
- Forrester, J.V. Privilege revisited: An evaluation of the eye’s defence mechanism. Eye 2009, 23, 756–766. [Google Scholar] [CrossRef] [Green Version]
- Pleyer, U.; Schlickeiser, S. The taming of the shrew? The immunology of corneal transplantation. Acta Ophthalmol. 2009, 87, 488–497. [Google Scholar] [CrossRef]
- Apte, R.S.; Sinha, D.; Mayhew, E.; Wistow, G.J.; Niederkorn, J.Y. Cutting edge: Role of macrophage migration inhibitory factor in inhibiting NK cell activity and preserving immune privilege. J. Immunol. 1998, 160, 5693–5696. [Google Scholar]
- Griffith, T.S.; Brunner, T.; Fletcher, S.M.; Green, D.R.; Ferguson, T.A. Fas ligand-induced apoptosis as a mechanism of immune privilege. Science 1995, 270, 1189–1192. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, M.C.; Rosenbaum, J.T.; Brown, J.; Planck, S.R.; Huang, X.; Armstrong, C.A.; Ansel, J.C. Novel production of interleukin-1 receptor antagonist peptides in normal human cornea. J. Clin. Investig. 1995, 95, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Namba, K.; Kitaichi, N.; Nishida, T.; Taylor, A.W. Induction of regulatory T cells by the immunomodulating cytokines alpha-melanocyte-stimulating hormone and transforming growth factor-beta2. J. Leukoc. Biol. 2002, 72, 946–952. [Google Scholar] [PubMed]
- Sheibani, N.; Sorenson, C.M.; Cornelius, L.A.; Frazier, W.A. Thrombospondin-1, a natural inhibitor of angiogenesis, is present in vitreous and aqueous humor and is modulated by hyperglycemia. Biochem. Biophys. Res. Commun. 2000, 267, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Sohn, J.H.; Kaplan, H.J.; Suk, H.J.; Bora, P.S.; Bora, N.S. Complement regulatory activity of normal human intraocular fluid is mediated by MCP, DAF, and CD59. Investig. Ophthalmol. Vis. Sci. 2000, 41, 4195–4202. [Google Scholar]
- Stuart, P.M.; Griffith, T.S.; Usui, N.; Pepose, J.; Yu, X.; Ferguson, T.A. CD95 ligand (FasL)-induced apoptosis is necessary for corneal allograft survival. J. Clin. Investig. 1997, 99, 396–402. [Google Scholar] [CrossRef]
- Sugita, S.; Streilein, J.W. Iris pigment epithelium expressing CD86 (B7-2) directly suppresses T cell activation in vitro via binding to cytotoxic T lymphocyte-associated antigen 4. J. Exp. Med. 2003, 198, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Sugita, S.; Yamada, Y.; Horie, S.; Nakamura, O.; Ishidoh, K.; Yamamoto, Y.; Yamagami, S.; Mochizuki, M. Induction of T regulatory cells by cytotoxic T-lymphocyte antigen-2alpha on corneal endothelial cells. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2598–2605. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.W.; Yee, D.G.; Streilein, J.W. Suppression of nitric oxide generated by inflammatory macrophages by calcitonin gene-related peptide in aqueous humor. Investig. Ophthalmol. Vis. Sci. 1998, 39, 1372–1378. [Google Scholar]
- Wilbanks, G.A.; Mammolenti, M.; Streilein, J.W. Studies on the induction of anterior chamber-associated immune deviation (ACAID). III. Induction of ACAID depends upon intraocular transforming growth factor-beta. Eur. J. Immunol. 1992, 22, 165–173. [Google Scholar] [CrossRef]
- Yamagami, S.; Kawashima, H.; Tsuru, T.; Yamagami, H.; Kayagaki, N.; Yagita, H.; Okumura, K.; Gregerson, D.S. Role of Fas-Fas ligand interactions in the immunorejection of allogeneic mouse corneal transplants. Transplantation 1997, 64, 1107–1111. [Google Scholar] [CrossRef] [PubMed]
- Niederkorn, J.Y. The induction of anterior chamber-associated immune deviation. Chem. Immunol. Allergy 2007, 92, 27–35. [Google Scholar] [PubMed]
- Stein-Streilein, J.; Streilein, J.W. Anterior chamber associated immune deviation (ACAID): Regulation, biological relevance, and implications for therapy. Int. Rev. Immunol. 2002, 21, 123–152. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, H.J.; Streilein, J.W. Immune response to immunization via the anterior chamber of the eye. I. F. lymphocyte-induced immune deviation. J. Immunol. 1977, 118, 809–814. [Google Scholar]
- Kaplan, H.J.; Streilein, J.W. Immune response to immunization via the anterior chamber of the eye. II. An analysis of F1 lymphocyte-induced immune deviation. J. Immunol. 1978, 120, 689–693. [Google Scholar]
- Ksander, B.R.; Streilein, J.W. Analysis of cytotoxic T cell responses to intracameral allogeneic tumors. Investig. Ophthalmol. Vis. Sci. 1989, 30, 323–329. [Google Scholar]
- Wilbanks, G.A.; Streilein, J.W. Distinctive humoral immune responses following anterior chamber and intravenous administration of soluble antigen. Evidence for active suppression of IgG2-secreting B lymphocytes. Immunology 1990, 71, 566–572. [Google Scholar]
- Hori, J.; Wang, M.; Miyashita, M.; Tanemoto, K.; Takahashi, H.; Takemori, T.; Okumura, K.; Yagita, H.; Azuma, M. B7-H1-induced apoptosis as a mechanism of immune privilege of corneal allografts. J. Immunol. 2006, 177, 5928–5935. [Google Scholar] [CrossRef] [Green Version]
- Kunishige, T.; Taniguchi, H.; Terada, M.; Akiba, H.; Yagita, H.; Abe, R.; Hori, J. Protective Role of ICOS and ICOS Ligand in Corneal Transplantation and in Maintenance of Immune Privilege. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6815–6823. [Google Scholar] [CrossRef]
- Kunishige, T.; Taniguchi, H.; Ohno, T.; Azuma, M.; Hori, J. VISTA Is Crucial for Corneal Allograft Survival and Maintenance of Immune Privilege. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4958–4965. [Google Scholar] [CrossRef] [Green Version]
- Hori, J.; Taniguchi, H.; Wang, M.; Oshima, M.; Azuma, M. GITR ligand-mediated local expansion of regulatory T cells and immune privilege of corneal allografts. Investig. Ophthal. Vis. Sci. 2010, 51, 6556–6565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimmura-Tomita, M.; Wang, M.; Taniguchi, H.; Akiba, H.; Yagita, H.; Hori, J. Galectin-9-mediated protection from allo-specific T cells as a mechanism of immune privilege of corneal allografts. PLoS ONE 2013, 8, e63620. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S.; Golstein, P. The Fas death factor. Science 1995, 267, 1449–1456. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A.; Dixit, V.M. Death receptors: Signaling and modulation. Science 1998, 281, 1305–1308. [Google Scholar] [CrossRef] [Green Version]
- Ashkenazi, A.; Dixit, V.M. Apoptosis control by death and decoy receptors. Curr. Opin. Cell Biol. 1999, 11, 255–260. [Google Scholar] [CrossRef]
- Wolf, B.B.; Green, D.R. Suicidal tendencies: Apoptotic cell death by caspase family proteinases. J. Biol. Chem. 1999, 274, 20049–20052. [Google Scholar] [CrossRef] [Green Version]
- Nagata, S. Apoptosis by death factor. Cell 1997, 88, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Hori, J.; Joyce, N.; Streilein, J.W. Epithelium-deficient corneal allografts display immune privilege beneath the kidney capsule. Investig. Ophthalmol. Vis. Sci. 2000, 41, 443–452. [Google Scholar]
- Hori, J.; Joyce, N.C.; Streilein, J.W. Immune privilege and immunogenicity reside among different layers of the mouse cornea. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3032–3042. [Google Scholar] [CrossRef]
- Ishida, Y.; Agata, Y.; Shibahara, K.; Honjo, T. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death. EMBO J. 1992, 11, 3887–3895. [Google Scholar] [CrossRef]
- Nishimura, H.; Nose, M.; Hirai, H.; Minato, N.; Honjo, T. Development of lupus-like autoimmune diseases by disruption of the PD-1 gene encoding and ITIM motif-carrying immunoreceptor. Immunity 1999, 11, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Sharpe, A.H.; Freeman, G.J. The B7-CD28 superfamily. Nat. Rev. Immunol. 2002, 2, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Zhu, G.; Tamada, K.; Chen, L. B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion. Nat. Med. 1999, 5, 1365–1369. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Strome, S.E.; Salomao, D.R.; Tamura, H.; Hirano, F.; Flies, D.B.; Roche, P.C.; Lu, J.; Zhu, G.; Tamada, K. Tumor-associated B7-H1 promotes T-cell apoptosis: A potential mechanism of immune evasion. Nat. Med. 2002, 8, 793–800. [Google Scholar] [CrossRef]
- Dong, H.; Zhu, G.; Tamada, K.; Flies, D.B.; van Deursen, J.M.; Chen, L. B7-H1 determines accumulation and deletion of intrahepatic CD8(+) T lymphocytes. Immunity 2004, 20, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Manzanet, R.; DeKruyff, R.; Kuchroo, V.K.; Umetsu, D.T. The costimulatory role of TIM molecules. Immunol. Rev. 2009, 229, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Ueno, T.; Habicht, A.; Clarkson, M.R.; Albin, M.J.; Yamaura, K.; Boenisch, O.; Popoola, J.; Wang, Y.; Yagita, H.; Akiba, H.; et al. The emerging role of T cell Ig mucin 1 in alloimmune responses in an experimental mouse transplant model. J. Clin. Investig. 2008, 118, 742–751. [Google Scholar] [CrossRef]
- Xiao, S.; Najafian, N.; Reddy, J.; Albin, M.; Zhu, C.; Jensen, E.; Imitola, J.; Korn, T.; Anderson, A.C.; Zhang, Z.; et al. Differential engagement of Tim-1 during activation can positively or negatively costimulate T cell expansion and effector function. J. Exp. Med. 2007, 204, 1691–1702. [Google Scholar] [CrossRef]
- Monney, L.; Sabatos, C.A.; Gaglia, J.L.; Ryu, A.; Waldner, H.; Chernova, T.; Manning, S.; Greenfield, E.A.; Coyle, A.J.; Sobel, R.A.; et al. Th1-specific cell surface protein Tim-3 regulates macrophage activation and severity of an autoimmune disease. Nature 2002, 415, 536–541. [Google Scholar] [CrossRef]
- Sanchez-Fueyo, A.; Tian, J.; Picarella, D.; Domenig, C.; Zheng, X.X.; Sabatos, C.A.; Manlongat, N.; Bender, O.; Kamradt, T.; Kuchroo, V.K.; et al. Tim-3 inhibits T helper type 1-mediated auto- and alloimmune responses and promotes immunological tolerance. Nat. Immunol. 2003, 4, 1093–1101. [Google Scholar] [CrossRef]
- Sabatos, C.A.; Chakravarti, S.; Cha, E.; Schubart, A.; Sanchez-Fueyo, A.; Zheng, X.X.; Coyle, A.J.; Strom, T.B.; Freeman, G.J.; Kuchroo, V.K. Interaction of Tim-3 and Tim-3 ligand regulates T helper type 1 responses and induction of peripheral tolerance. Nat. Immunol. 2003, 4, 1102–1110. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.C.; Anderson, D.E.; Bregoli, L.; Hastings, W.D.; Kassam, N.; Lei, C.; Chandwaskar, R.; Karman, J.; Su, E.W.; Hirashima, M.; et al. Promotion of tissue inflammation by the immune receptor Tim-3 expressed on innate immune cells. Science 2007, 318, 1141–1143. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Anderson, A.C.; Schubart, A.; Xiong, H.; Imitola, J.; Khoury, S.J.; Zheng, X.X.; Strom, T.B.; Kuchroo, V.K. The Tim-3 ligand galectin-9 negatively regulates T helper type 1 immunity. Nat. Immunol. 2005, 6, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Hutloff, A.; Dittrich, A.M.; Beier, K.C.; Eljaschewitsch, B.; Kraft, R.; Anagnostopoulos, I.; Kroczek, R.A. ICOS is an inducible T-cell co-stimulator structurally and functionally related to CD28. Nature 1999, 397, 263–266. [Google Scholar] [CrossRef]
- Yoshinaga, S.K.; Whoriskey, J.S.; Khare, S.D.; Sarmiento, U.; Guo, J.; Horan, T.; Shih, G.; Zhang, M.; Coccia, M.A.; Kohno, T.; et al. T-cell co-stimulation through B7RP-1 and ICOS. Nature 1999, 402, 827–832. [Google Scholar] [CrossRef]
- Tezuka, K.; Tsuji, T.; Hirano, D.; Tamatani, T.; Sakamaki, K.; Kobayashi, Y.; Kamada, M.; Tafuri-Bladt, A. Identification and characterization of rat AILIM/ICOS, a novel T-cell costimulatory molecule, related to the CD28/CTLA4 family. Biochem. Biophys. Res. Commun. 2000, 276, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Swallow, M.M.; Wallin, J.J.; Sha, W.C. B7h, a novel costimulatory homolog of B7.1 and B7.2, is induced by TNFalpha. Immunity 1999, 11, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Rubinstein, R.; Lines, J.L.; Wasiuk, A.; Ahonen, C.; Guo, Y.; Lu, L.-F.; Gondek, D.; Wang, Y.; Fava, R.A.; et al. VISTA, a novel mouse Ig superfamily ligand that negatively regulates T cell responses. J. Exp. Med. 2011, 208, 577–592. [Google Scholar] [CrossRef]
- Flies, D.B.; Wang, S.; Xu, H.; Chen, L. Cutting edge: A monoclonal antibody specific for the programmed death-1 homolog prevents graft-versus-host disease in mouse models. J. Immunol. 2011, 187, 1537–1541. [Google Scholar] [CrossRef]
- Flies, D.B.; Han, X.; Higuchi, T.; Zheng, L.; Sun, J.; Ye, J.J.; Chen, L. Coinhibitory receptor PD-1H preferentially suppresses CD4(+) T cell-mediated immunity. J. Clin. Investig. 2014, 124, 1966–1975. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Le Mercier, I.; Kuta, A.; Noelle, R.J. A New VISTA on combination therapy for negative checkpoint regulator blockade. J. Immunother. Cancer 2016, 4, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurney, A.L.; Marsters, S.A.; Huang, R.M.; Pitti, R.M.; Mark, D.T.; Baldwin, D.T.; Gray, A.M.; Dowd, A.D.; Brush, A.D.; Heldens, A.D. Identification of a new member of the tumor necrosis factor family and its receptor, a human ortholog of mouse GITR. Curr. Biol. 1999, 9, 215–218. [Google Scholar] [CrossRef] [Green Version]
- Nocentini, G.; Giunchi, L.; Ronchetti, S.; Krausz, L.T.; Bartoli, A.; Moraca, R.; Migliorati, G.; Riccardi, C. A new member of the tumor necrosis factor/nerve growth factor receptor family inhibits T cell receptor-induced apoptosis. Proc. Natl. Acad. Sci. USA 1997, 94, 6216–6221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawashima, H.; Prasad, S.A.; Gregerson, D.S. Corneal endothelial cells inhibit T cell proliferation by blocking IL-2 production. J. Immunol. 1994, 153, 1982–1989. [Google Scholar]
- Obritsch, W.F.; Kawashima, H.; Evangelista, A.; Ketcham, J.M.; Holland, E.J.; Gregerson, D.S. Inhibition of in vitro T cell activation by corneal endothelial cells. Cell. Immunol. 1992, 144, 80–94. [Google Scholar] [CrossRef]
- Sugita, S.; Usui, Y.; Horie, S.; Futagami, Y.; Yamada, Y.; Ma, J.; Kezuka, T.; Hamada, H.; Usui, T.; Mochizuki, M. Human corneal endothelial cells expressing programmed death-ligand 1 (PD-L1) suppress PD-1+ T helper 1 cells by a contact-dependent mechanism. Investig. Ophthalmol. Vis. Sci. 2009, 50, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Sugita, S.; Horie, S.; Yamagami, S.; Mochizuki, M. Mechanisms of immune suppression for CD8+ T cells by human corneal endothelial cells via membrane-bound TGFbeta. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2548–2557. [Google Scholar] [CrossRef] [Green Version]
- Zou, W.; Chen, L. Inhibitory B7-family molecules in the tumour microenvironment. Nat. Rev. Immunol. 2008, 8, 467–477. [Google Scholar] [CrossRef]
- Pardoll, D.; Drake, C. Immunotherapy earns its spot in the ranks of cancer therapy. J. Exp. Med. 2012, 209, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Allison, J.P. The future of immune checkpoint therapy. Science 2015, 348, 56–61. [Google Scholar] [CrossRef]
- Postow, M.A.; Callahan, M.K.; Wolchok, J.D. Immune Checkpoint Blockade in Cancer Therapy. J. Clin. Oncol. 2015, 33, 1974–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Allison, J.P. Immune checkpoint targeting in cancer therapy: Toward combination strategies with curative potential. Cell 2015, 161, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Hieu, T.; Malarkannan, S.; Wang, L. The structure, expression, and multifaceted role of immune-checkpoint protein VISTA as a critical regulator of anti-tumor immunity, autoimmunity, and inflammation. Cell. Mol. Immunol. 2018, 15, 438–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxi, S.; Yang, A.; Gennarelli, R.L.; Khan, N.; Wang, Z.; Boyce, L.; Korenstein, D. Immune-related adverse events for anti-PD-1 and anti-PD-L1 drugs: Systematic review and meta-analysis. BMJ 2018, 360, k793. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Wahab, N.; Suarez-Almazor, M.E. Frequency and distribution of various rheumatic disorders associated with checkpoint inhibitor therapy. Rheumatology 2019, 58, vii40–vii48. [Google Scholar] [CrossRef] [Green Version]
- Dalvin, L.A.; Shields, C.L.; Orloff, M.; Sato, T.; Shields, J.A. CHECKPOINT INHIBITOR IMMUNE THERAPY: Systemic Indications and Ophthalmic Side Effects. Retina 2018, 38, 1063–1078. [Google Scholar] [CrossRef]
- Antoun, J.; Titah, C.; Cochereau, I. Ocular and orbital side-effects of checkpoint inhibitors: A review article. Curr. Opin. Oncol. 2016, 28, 288–294. [Google Scholar] [CrossRef]
- Nguyen, A.T.; Elia, M.; Materin, M.A.; Sznol, M.; Chow, J. Cyclosporine for Dry Eye Associated with Nivolumab: A Case Progressing to Corneal Perforation. Cornea 2016, 35, 399–401. [Google Scholar] [CrossRef]
- Ileana Dumbrava, E.; Smith, V.; Alfattal, R.; El-Naggar, A.K.; Penas-Prado, M.; Tsimberidou, A.M. Autoimmune Granulomatous Inflammation of Lacrimal Glands and Axonal Neuritis Following Treatment with Ipilimumab and Radiation Therapy. J. Immunother. 2018, 41, 336–339. [Google Scholar] [CrossRef]
- Eckert, A.; Schoeffler, A.; Dalle, S.; Phan, A.; Kiakouama, L.; Thomas, L. Anti-CTLA4 monoclonal antibody induced sarcoidosis in a metastatic melanoma patient. Dermatology 2008, 218, 69–70. [Google Scholar] [CrossRef]
- Suozzi, K.C.; Stahl, M.; Ko, C.J.; Chiang, A.; Gettinger, S.N.; Siegel, M.D.; Bunick, C.G. Immune-related sarcoidosis observed in combination ipilimumab and nivolumab therapy. JAAD Case Rep. 2016, 2, 264–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegenhagen, M.W.; Müller-Quernheim, J. The cytokine network in sarcoidosis and its clinical relevance. J. Intern. Med. 2003, 253, 18–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soluble Factors in the Anterior Chamber (Target Cells/Factors to Suppress) | Cell Surface Molecules of the Cornea and Iris–Ciliary Body (Target Cells/Factors to Suppress) |
|---|---|
| α-MSH (T cells, macrophages, neutrophils) | PD-L1 (B7-H1) (T cells) |
| VIP (T cells) | ICOSL (B7-H2, B7RP-1) (T cells) |
| Somatostatin (T cells) | VISTA (APCs, T cells) |
| CGRP (macrophages) | GITR ligand (T cells) |
| TGF-β2 (T cells, macrophages, NK cells) | Galectin-9 (T cells) |
| TSP-1 (macrophages) | TRAIL (T cells) |
| MIF (NK cells) | B7-2 (via CTLA4) (T cells) |
| IL-1Ra (IL-1) | CTLA-2α (T cells) |
| sFas L (T cells, neutrophils) | Fas L (CD95 L) (T cells, neutrophils) |
| CD46, CD55, CD59, C3ib (complement) | MHC class Ib (T cells, NK cells) |
| CD46, CD55, CD59 (complement) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hori, J.; Kunishige, T.; Nakano, Y. Immune Checkpoints Contribute Corneal Immune Privilege: Implications for Dry Eye Associated with Checkpoint Inhibitors. Int. J. Mol. Sci. 2020, 21, 3962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113962
Hori J, Kunishige T, Nakano Y. Immune Checkpoints Contribute Corneal Immune Privilege: Implications for Dry Eye Associated with Checkpoint Inhibitors. International Journal of Molecular Sciences. 2020; 21(11):3962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113962
Chicago/Turabian StyleHori, Junko, Tomoyuki Kunishige, and Yuji Nakano. 2020. "Immune Checkpoints Contribute Corneal Immune Privilege: Implications for Dry Eye Associated with Checkpoint Inhibitors" International Journal of Molecular Sciences 21, no. 11: 3962. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113962