Anticancer and Antiangiogenic Activities of Novel α-Mangostin Glycosides in Human Hepatocellular Carcinoma Cells via Downregulation of c-Met and HIF-1α


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of α-Mangostin Glycosides on the Growth of HCC Cells
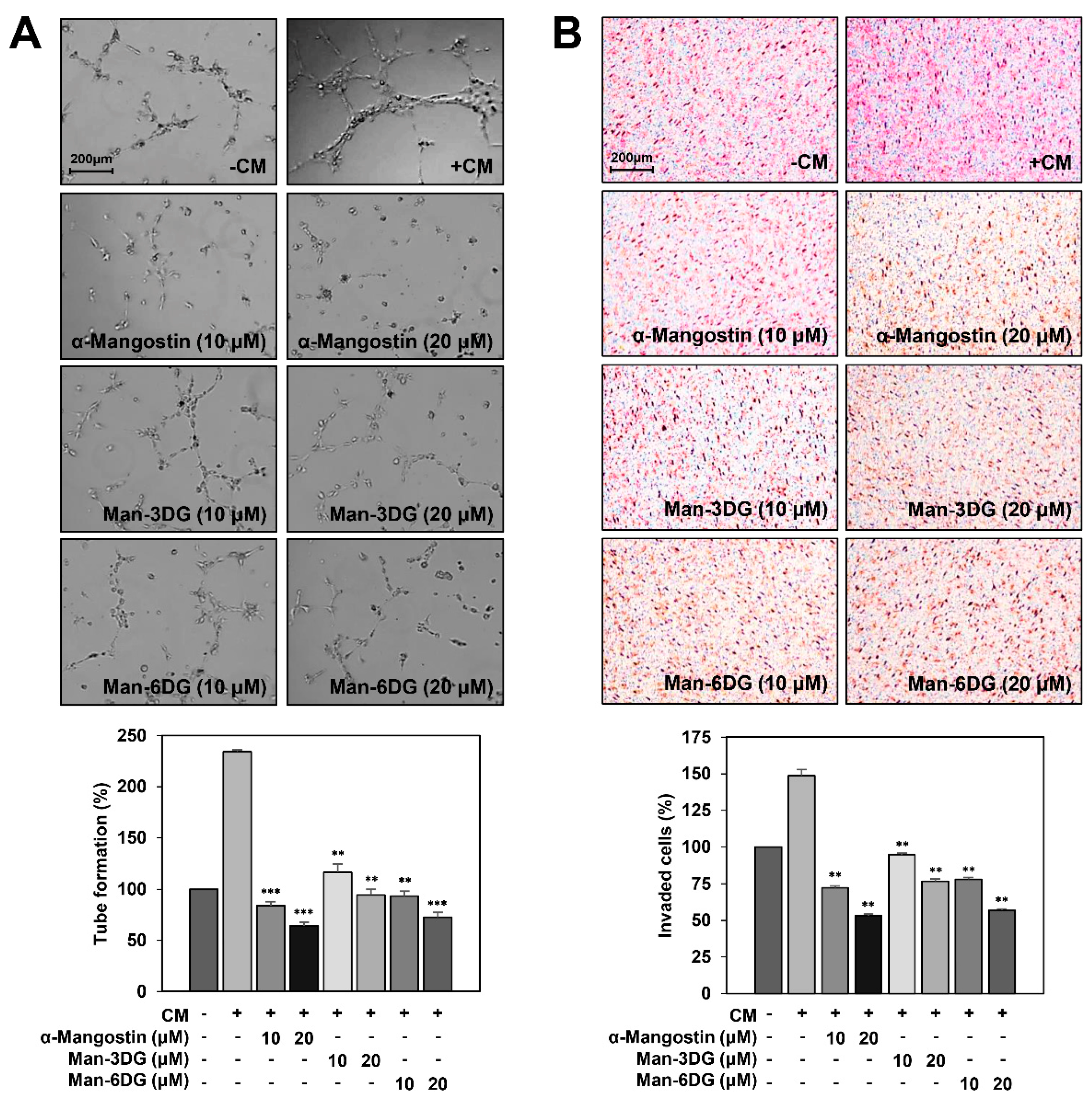
2.2. Effects of α-Mangostin Glycosides on the Migration of Hep3B Cells
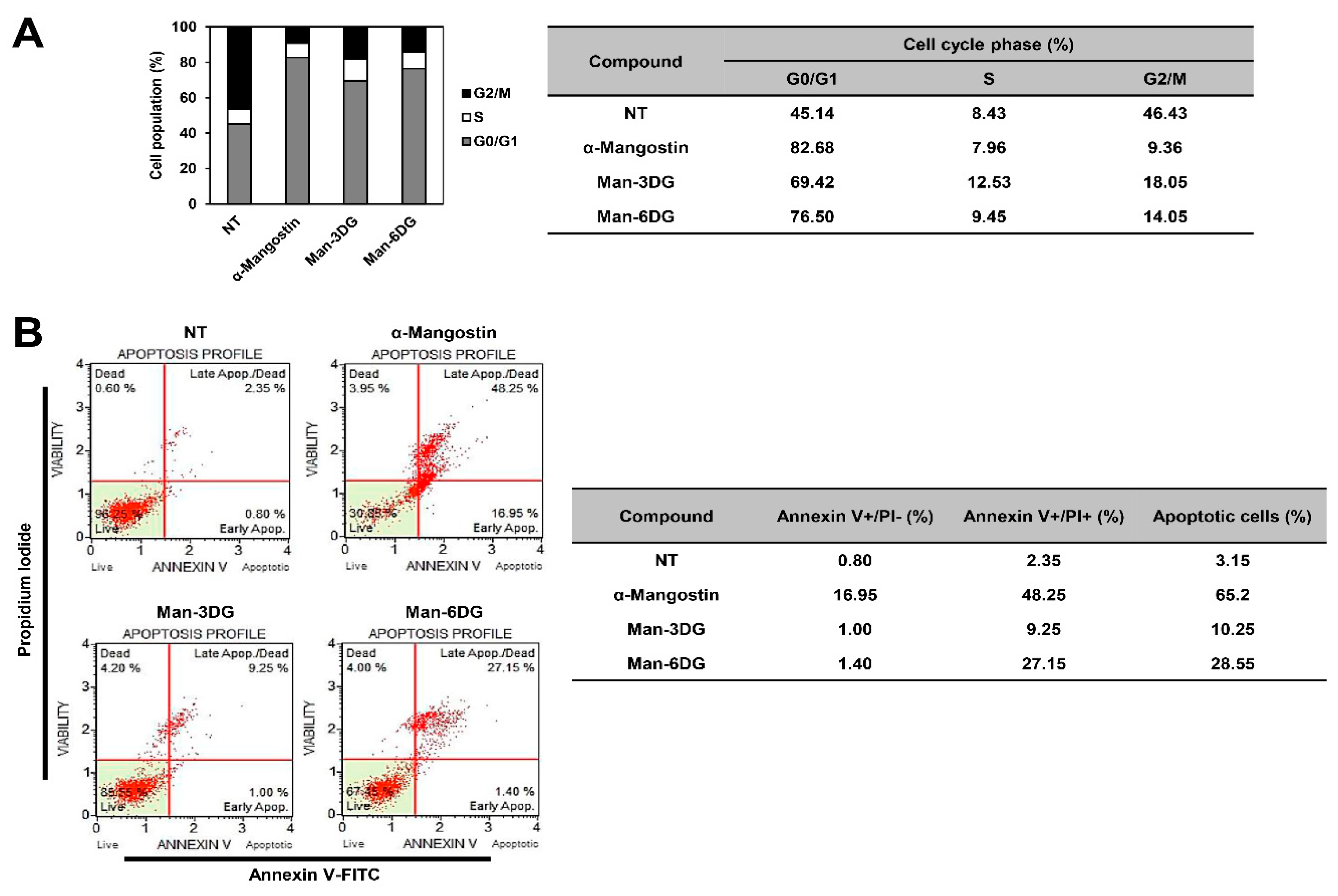
2.3. Effects of α-Mangostin Glycosides on the Apoptosis of Hep3B Cells
2.4. Effects of α-Mangostin Glycosides on the Autophagy of Hep3B Cells
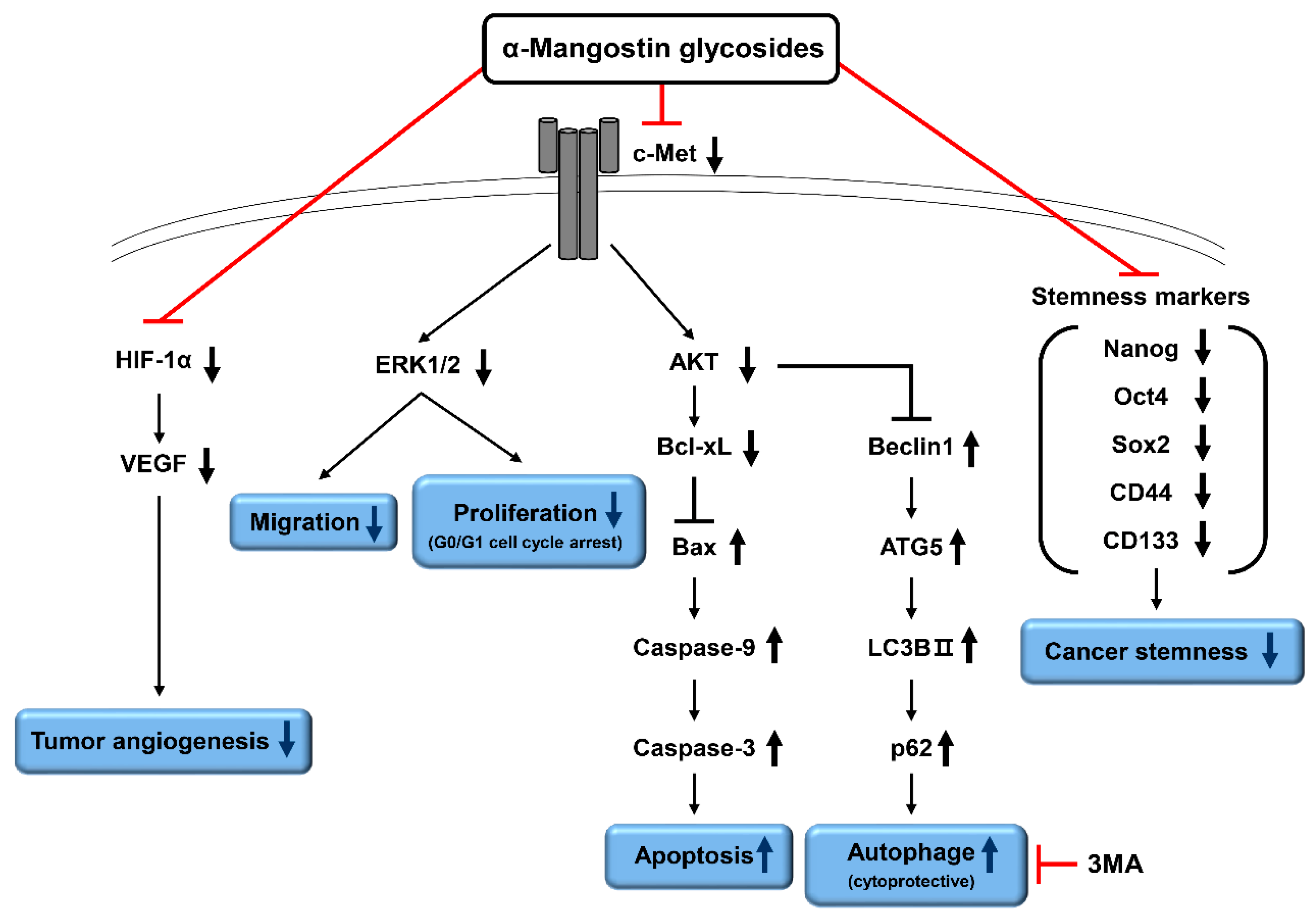
2.5. Effects of α-Mangostin Glycosides on the c-Met Signaling Pathway in Hep3B Cells
2.6. Effects of α-Mangostin Glycosides on the Hep3B Cell-Induced Angiogenesis
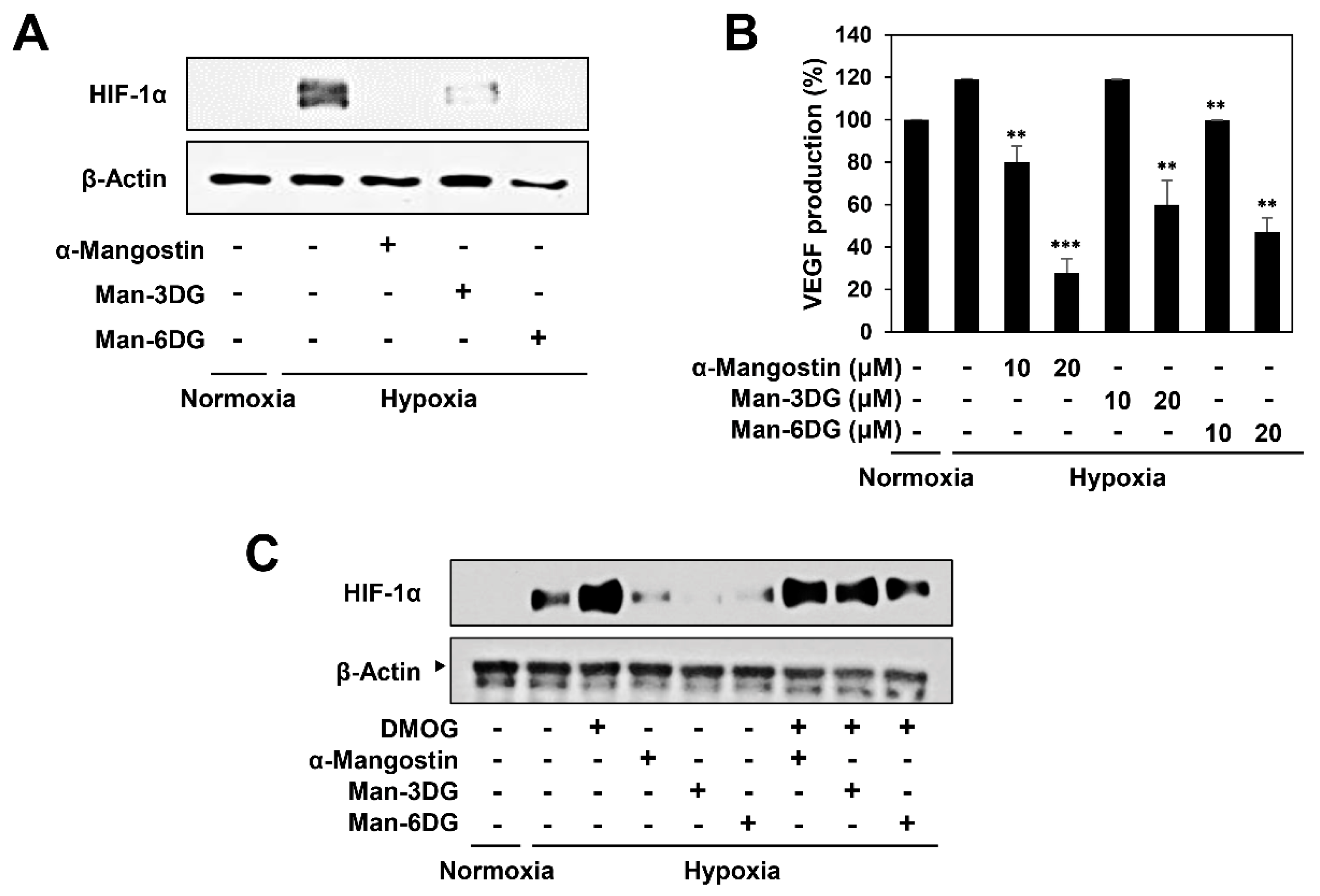
2.7. Effects of α-Mangostin Glycosides on the Hypoxia-Induced Accumulation of HIF-1α Protein
2.8. Effects of α-Mangostin Glycosides on the Tumorsphere Formation and Cancer Stemness of Hep3B Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Hypoxic Conditions
4.3. Cell Growth Assay
4.4. Colony Formation Assay
4.5. Wound Healing Assay
4.6. Cell Cycle Analysis
4.7. Apoptosis Analysis
4.8. Western Blot Analysis
4.9. Tumor Cell-Induced Angiogenesis Assay
4.10. VEGF Measurement by Enzyme-Linked Immunosorbent Assay (ELISA)
4.11. 3D Cell Culture
4.12. CellTiter-Glo® Luminescent Cell Viability Assay
4.13. Tumorsphere Forming Assay
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| HCC | Hepatocellular carcinoma |
| Man-3DG | α-Mangostin-3-O-β-D-2-deoxyglucopyranoside |
| Man-6DG | α-Mangostin 6-O-β-D-2-deoxyglucopyranoside |
| HIF-1α | Hypoxia-inducible factor-1α |
| VEGF | Vascular endothelial growth factor |
| HGF | Hepatic growth factor |
| MAPK | Mitogen activated protein kinase |
| PI3K | Phosphatidylinositol-3 kinase |
| AKT | Protein kinase B |
| STAT | Signal transducer and activator of transcription |
| FITC | Fluorescein isothiocyanate |
| PI | Propidium iodide |
| 3-MA | 3-Methyladenine |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| ERK | Extracellular signal-regulated kinase |
| HUVEC | Human umbilical vein endothelial cell |
| CM | Conditioned medium |
| AMPK | AMP-activated protein kinase |
| mTORC1 | Mammalian target of rapamycin complex 1 |
| HGFR | Hepatocyte growth factor receptor |
| PHD | Prolyl-hydroxylase |
| DMOG | Dimethyloxalylglycine |
| CSC | Cancer stem cell |
| Bcl-XL | B-cell lymphoma-extra large |
| Bax | Bcl-2-associated X protein |
| Atg5 | Autophagy related 5 |
| LC3B | Microtubule-associated proteins 1A/1B light chain 3B |
| p62 | Nucleoporin p62 |
| Oct4 | Octamer-binding transcription factor 4 |
| Sox2 | Sex-determining region Y-box 2 |
References
- El-Serag, H.B. Hepatocellular carcinoma. N. Engl. J. Med. 2011, 365, 1118–1127. [Google Scholar] [CrossRef]
- Chacko, S.; Samanta, S. Hepatocellular carcinoma: A life-threatening disease. Biomed. Pharmacother. 2016, 84, 1679–1688. [Google Scholar] [CrossRef]
- Yu, M.C.; Yuan, J.M. Environmental factors and risk for hepatocellular carcinoma. Gastroenterology 2004, 127, S72–S78. [Google Scholar] [CrossRef]
- Villanueva, A.; Hernandez-Gea, V.; Llovet, J.M. Medical therapies for hepatocellular carcinoma: A critical view of the evidence. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 34–42. [Google Scholar] [CrossRef]
- Kondo, S.; Ojima, H.; Tsuda, H.; Hashimoto, J.; Morizane, C.; Ikeda, M.; Ueno, H.; Tamura, K.; Shimada, K.; Kanai, Y.; et al. Clinical impact of c-Met expression and its gene amplification in hepatocellular carcinoma. Int. J. Clin. Oncol. 2013, 18, 207–213. [Google Scholar] [CrossRef]
- Kaposi-Novak, P.; Lee, J.S.; Gomez-Quiroz, L.; Coulouarn, C.; Factor, V.M.; Thorgeirsson, S.S. Met-regulated expression signature defines a subset of human hepatocellular carcinomas with poor prognosis and aggressive phenotype. J. Clin. Investig. 2006, 116, 1582–1595. [Google Scholar] [CrossRef] [PubMed]
- Goyal, L.; Muzumdar, M.D.; Zhu, A.X. Targeting the HGF/c-MET Pathway in Hepatocellular Carcinoma. Clin. Cancer. Res. 2013, 19, 2310–2318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, X.S.; Guo, X.Z.; Han, G.H.; Li, H.Y.; Chen, J. MET inhibitors for treatment of advanced hepatocellular carcinoma: A. review. World J. Gastroenterol. 2015, 21, 5445–5453. [Google Scholar] [CrossRef]
- Okuma, H.S.; Kondo, S. Trends in the development of MET inhibitors for hepatocellular carcinoma. Future Oncol. 2016, 12, 1275–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, A.X.; Duda, D.G.; Sahani, D.V.; Jain, R.K. HCC and angiogenesis: Possible targets and future directions. Nat. Rev. Clin. Oncol. 2011, 8, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.F.; de Oliveira, A.C.; Santoro, A.; Raoul, J.L.; Forner, A.; et al. Sorafenib in advanced hepatocellular carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Wang, Z.; Wu, J.; Jiang, C.; Wu, J. The role of hypoxia inducible factor-1 in hepatocellular carcinoma. BioMed Res. Int. 2014, 2014, 409272. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Wu, J. Hypoxia inducible factor in hepatocellular carcinoma: A therapeutic target. World J. Gastroenterol. 2015, 21, 12171–12178. [Google Scholar] [CrossRef]
- Chen, C.; Lou, T. Hypoxia inducible factors in hepatocellular carcinoma. Oncotarget 2017, 8, 46691–46703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jindarat, S. Xanthones from mangosteen (Garcinia mangostana): Multi-targeting pharmacological properties. J. Med. Assoc. Thail. 2014, 97 (Suppl. 2), S196–S201. [Google Scholar]
- Pedraza-Chaverri, J.; Cárdenas-Rodríguez, N.; Orozco-Ibarra, M.; Pérez-Rojas, J.M. Medicinal properties of mangosteen (Garcinia mangostana). Food Chem. Toxicol. 2008, 46, 3227–3239. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.J.; Petiwala, S.M.; Syed, D.N.; Rasmussen, J.T.; Adhami, V.M.; Siddiqui, I.A.; Kohl, A.M.; Mukhtar, H. α-mangostin, a xanthone from mangosteen fruit, promotes cell cycle arrest in prostate cancer and decreases xenograft tumor growth. Carcinogenesis 2012, 33, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Watanapokasin, R.; Jarinthanan, F.; Nakamura, Y.; Sawasjirakij, N.; Jaratrungtawee, A.; Suksamrarn, S. Effects of α-mangostin on apoptosis induction of human colon cancer. World J. Gastroenterol. 2011, 17, 2086–2095. [Google Scholar] [CrossRef]
- Zhang, K.J.; Gu, Q.L.; Yang, K.; Ming, X.J.; Wang, J.X. Anticarcinogenic Effects of α-Mangostin: A Review. Planta Med. 2017, 83, 188–202. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Brunner, I.; Han, A.R.; Hamburger, M.; Kinghorn, A.D.; Frye, R.; Butterweck, V. Pharmacokinetics of Alpha-Mangostin in Rats After Intravenous and Oral Application. Mol. Nutr. Food Res. 2011, 55 (Suppl. 1), S67–S74. [Google Scholar] [CrossRef]
- Choi, Y.H.; Han, S.Y.; Kim, Y.J.; Kim, Y.M.; Chin, Y.W. Absorption, tissue distribution, tissue metabolism and safety of α-mangostin in mangosteen extract using mouse models. Food Chem. Toxicol. 2014, 66, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; You, B.H.; Kim, Y.C.; Chin, Y.W.; Choi, Y.H. Dose-Independent ADME Properties and Tentative Identification of Metabolites of α-Mangostin from Garcinia mangostana in Mice by Automated Microsampling and UPLC-MS/MS Methods. PLoS ONE 2015, 10, e0131587. [Google Scholar] [CrossRef] [PubMed]
- Moradi, S.V.; Hussein, W.M.; Varamini, P.; Simerska, P.; Toth, I. Glycosylation, an effective synthetic strategy to improve the bioavailability of therapeutic peptides. Chem. Sci. 2016, 7, 2492–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Wu, Z.; Zhang, L. Clinical applications of the naturally occurring or synthetic glycosylated low molecular weight drugs. Prog. Mol. Biol. Transl. Sci. 2019, 163, 487–522. [Google Scholar]
- Rodriguez, M.C.; Cudic, M. Optimization of physicochemical and pharmacological properties of peptide drugs by glycosylation. Methods Mol. Biol. 2013, 1081, 107–136. [Google Scholar]
- Le, T.T.; Pandey, R.P.; Gurung, R.B.; Dhakal, D.; Sohng, J.K. Efficient enzymatic systems for synthesis of novel α-mangostin glycosides exhibiting antibacterial activity against Gram-positive bacteria. Appl. Microbiol. Biotechnol. 2014, 98, 8527–8538. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Li, Y.; Zhang, G. Cell cycle regulation and anticancer drug discovery. Cancer Biol. Med. 2017, 14, 348–362. [Google Scholar]
- Hsieh, S.C.; Huang, M.H.; Cheng, C.W.; Hung, J.H.; Yang, S.F.; Hsieh, Y.H. α-mangostin induces mitochondrial dependent apoptosis in human hepatoma SK-Hep-1 cells through inhibition of p38 MAPK pathway. Apoptosis 2013, 18, 1548–1560. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Peng, Z.L. Programmed cell death and cancer. Postgrad. Med. J. 2009, 85, 134–140. [Google Scholar] [CrossRef]
- El-Khattouti, A.; Selimovic, D.; Haikel, Y.; Hassan, M. Crosstalk between apoptosis and autophagy: Molecular mechanisms and therapeutic strategies in cancer. J. Cell Death. 2013, 6, 37–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Feng, Y. Elaborating the role of natural products-induced autophagy in cancer treatment: Achievements and artifacts in the state of the art. BioMed Res. Int. 2015, 2015, 934207. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Liang, Y.; Ma, X. Alpha-mangostin induces endoplasmic reticulum stress and autophagy which count against fatty acid synthase inhibition mediated apoptosis in human breast cancer cells. Cancer Cell Int. 2019, 19, 151. [Google Scholar] [CrossRef] [PubMed]
- Jittiporn, K.; Suwanpradid, J.; Patel, C.; Rojas, M.; Thirawarapan, S.; Moongkarndi, P.; Suvitayavat, W.; Caldwell, R.B. Anti-angiogenic actions of the mangosteen polyphenolic xanthone derivative α-mangostin. Microvasc. Res. 2014, 93, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, Ł.; Lamperska, K. 2D and 3D cell cultures—a comparison of different types of cancer cell cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar] [CrossRef]
- Shin, H.J.; Han, J.M.; Choi, Y.S.; Jung, H.J. Pterostilbene suppresses both cancer cells and cancer stem-like cells in cervical cancer with superior bioavailability to resveratrol. Molecules 2020, 25, 228. [Google Scholar] [CrossRef] [Green Version]
- Afify, S.M.; Sanchez, C.A.; Hassan, G.; Kumon, K.; Nawara, H.M.; Zahra, M.H.; Mansour, H.M.; Khayrani, A.C.; Alam, M.J.; Du, J.; et al. A novel model of liver cancer stem cells developed from induced pluripotent stem cells. Br. J. Cancer 2020, 122, 1378–1390. [Google Scholar] [CrossRef] [Green Version]
- Bielecka, Z.F.; Maliszewska-Olejniczak, K.; Safir, I.J.; Szczylik, C.; Czarnecka, A.M. Three-dimensional cell culture model utilization in cancer stem cell research. Biol. Rev. 2017, 92, 1505–1520. [Google Scholar] [CrossRef]
- Cragg, G.M.; Pezzuto, J.M. Natural Products as a Vital Source for the Discovery of Cancer Chemotherapeutic and Chemopreventive Agents. Med. Princ. Pract. 2016, 25 (Suppl. 2), 41–59. [Google Scholar] [CrossRef]
- Zeng, Z.P.; Jiang, J.G. Analysis of the adverse reactions induced by natural product-derived drugs. Br. J. Pharmacol. 2010, 159, 1374–1391. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Morris-Natschke, S.L.; Lee, K.H. Strategies for the Optimization of Natural Leads to Anticancer Drugs or Drug Candidates. Med. Res. Rev. 2016, 36, 32–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fei, X.; Jo, M.; Lee, B.; Han, S.B.; Lee, K.; Jung, J.K.; Seo, S.Y.; Kwak, Y.S. Synthesis of xanthone derivatives based on α-mangostin and their biological evaluation for anti-cancer agents. Bioorg. Med. Chem. Lett. 2014, 24, 2062–2065. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tang, G.; Tang, Q.; Zhang, J.; Hou, Y.; Cai, E.; Liu, S.; Lei, D.; Zhang, L.; Wang, S. A Method of Effectively Improved α-Mangostin Bioavailability. Eur. J. Drug Metab. Pharmacokinet. 2016, 41, 605–613. [Google Scholar] [CrossRef]
- Gurung, R.B.; Gong, S.Y.; Dhakal, D.; Le, T.T.; Jung, N.R.; Jung, H.J.; Oh, T.J.; Sohng, J.K. Synthesis of Curcumin Glycosides with Enhanced Anticancer Properties Using One-Pot Multienzyme Glycosylation Technique. J. Microbiol. Biotechnol. 2017, 27, 1639–1648. [Google Scholar] [CrossRef] [Green Version]
- Xiong, S.; Mu, T.; Wang, G.; Jiang, X. Mitochondria-mediated apoptosis in mammals. Protein Cell 2014, 5, 737–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.J.; Long, Z.J.; Xu, D.F.; Xiao, R.Z.; Liu, L.L.; Xu, Z.F.; Qiu, S.X.; Lin, D.J.; Liu, Q. Inhibition of autophagy augments the anticancer activity of α-mangostin in chronic myeloid leukemia cells. Leuk. Lymphoma 2014, 55, 628–638. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, Y.; Tooze, S.A. Autophagy pathway: Cellular and molecular mechanisms. Autophagy 2018, 14, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.A.; Choy, H.; Habib, A.A.; Saha, D. SNS-032 prevents tumor cell-induced angiogenesis by inhibiting vascular endothelial growth factor. Neoplasia (New York, NY) 2007, 9, 370. [Google Scholar] [CrossRef] [Green Version]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.M.; Han, J.M.; Le, T.T.; Sohng, J.K.; Jung, H.J. Anticancer and Antiangiogenic Activities of Novel α-Mangostin Glycosides in Human Hepatocellular Carcinoma Cells via Downregulation of c-Met and HIF-1α. Int. J. Mol. Sci. 2020, 21, 4043. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21114043
Kim SM, Han JM, Le TT, Sohng JK, Jung HJ. Anticancer and Antiangiogenic Activities of Novel α-Mangostin Glycosides in Human Hepatocellular Carcinoma Cells via Downregulation of c-Met and HIF-1α. International Journal of Molecular Sciences. 2020; 21(11):4043. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21114043
Chicago/Turabian StyleKim, Sung Min, Jang Mi Han, Tuoi Thi Le, Jae Kyung Sohng, and Hye Jin Jung. 2020. "Anticancer and Antiangiogenic Activities of Novel α-Mangostin Glycosides in Human Hepatocellular Carcinoma Cells via Downregulation of c-Met and HIF-1α" International Journal of Molecular Sciences 21, no. 11: 4043. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21114043