3-D Nucleus Architecture in Oat × Maize Addition Lines

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
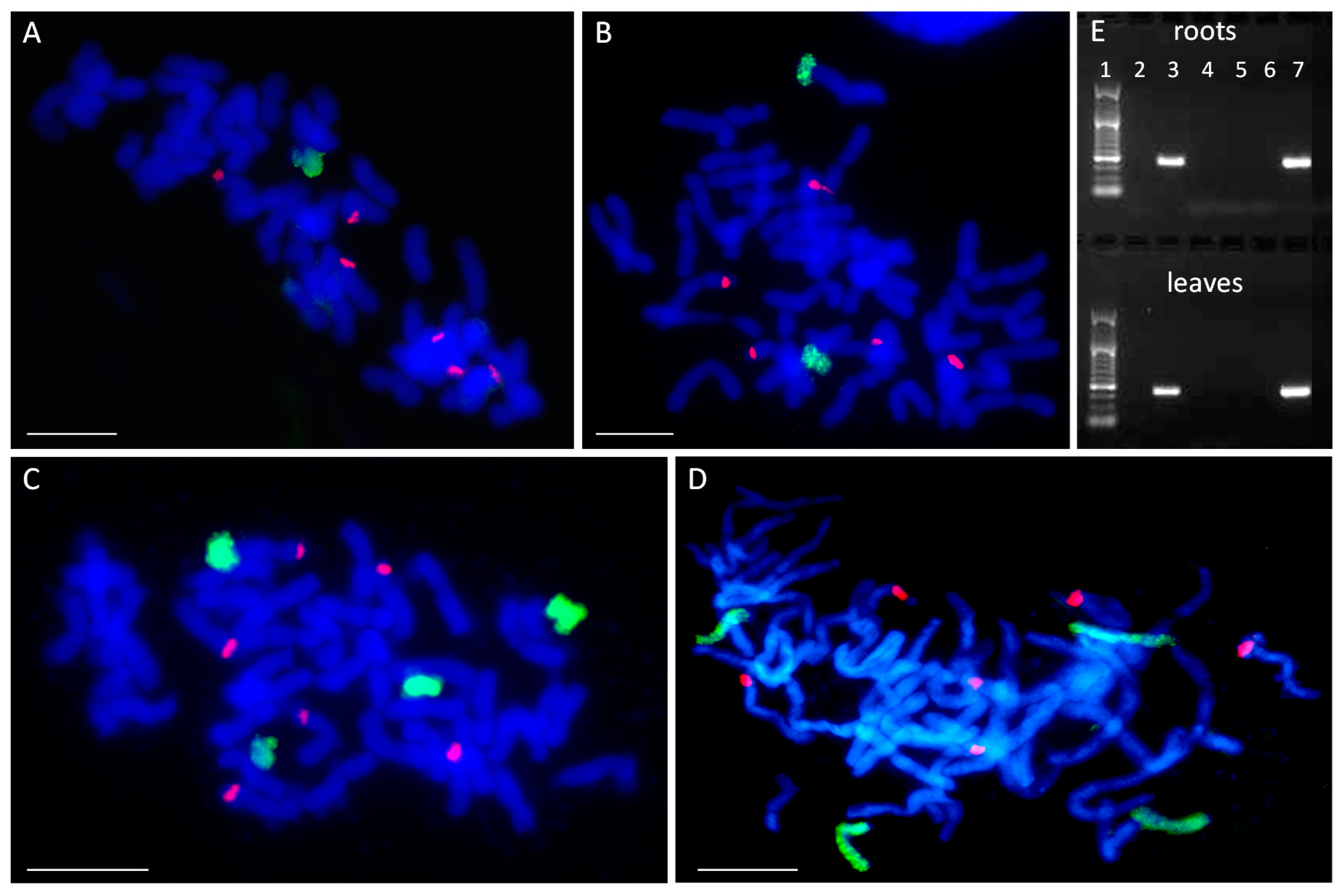
2.1. Number and Identification of Maize Chromosome Additions
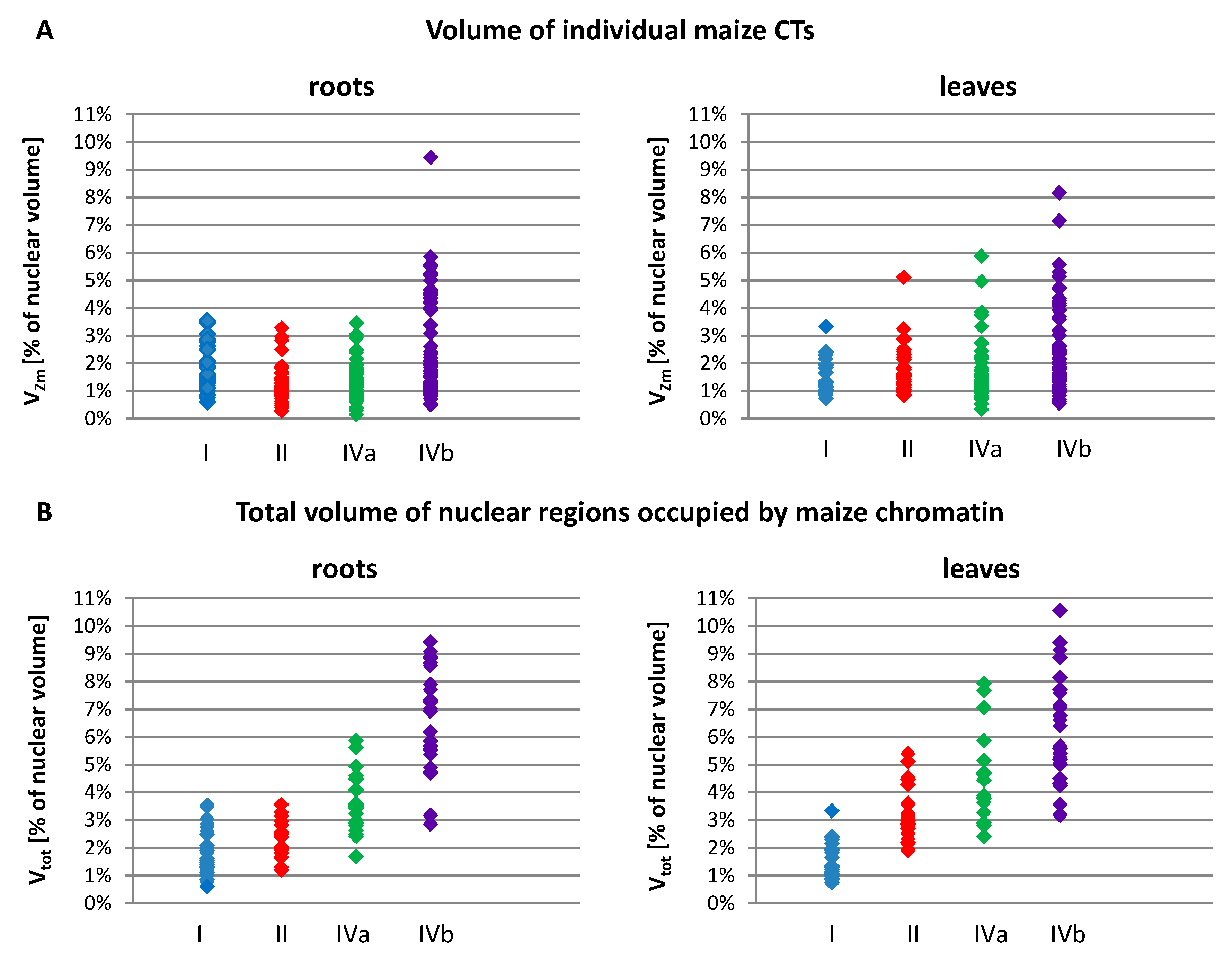
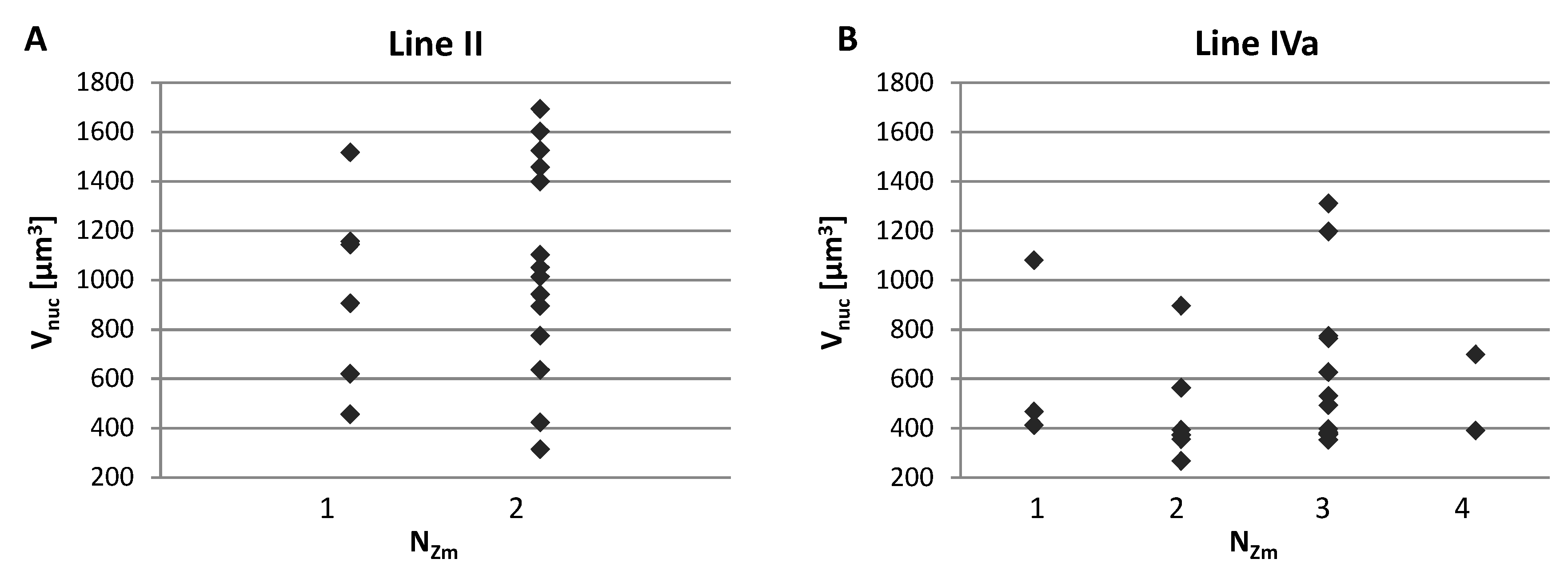
2.2. Size and Associations of Maize Chromosome Territories
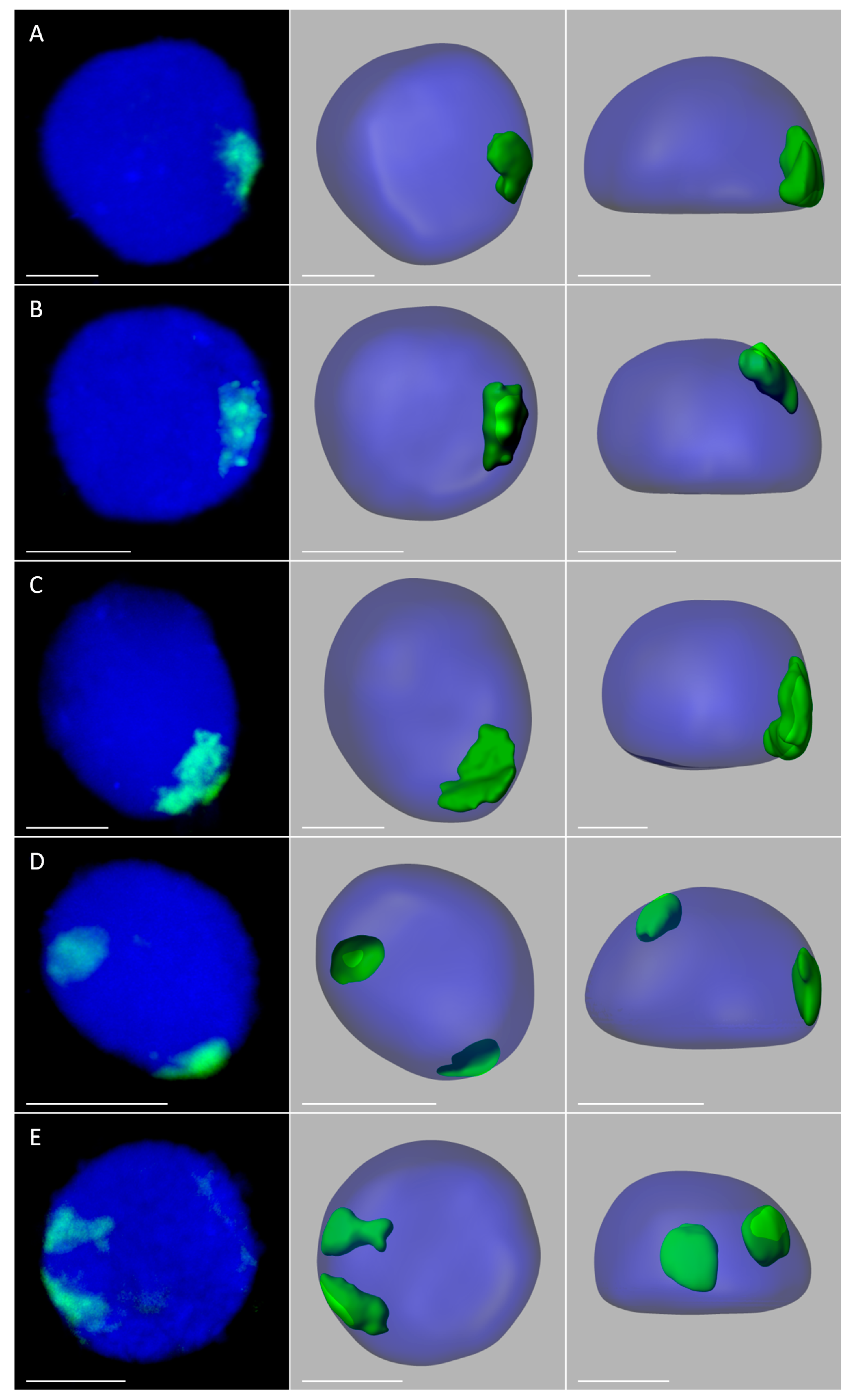
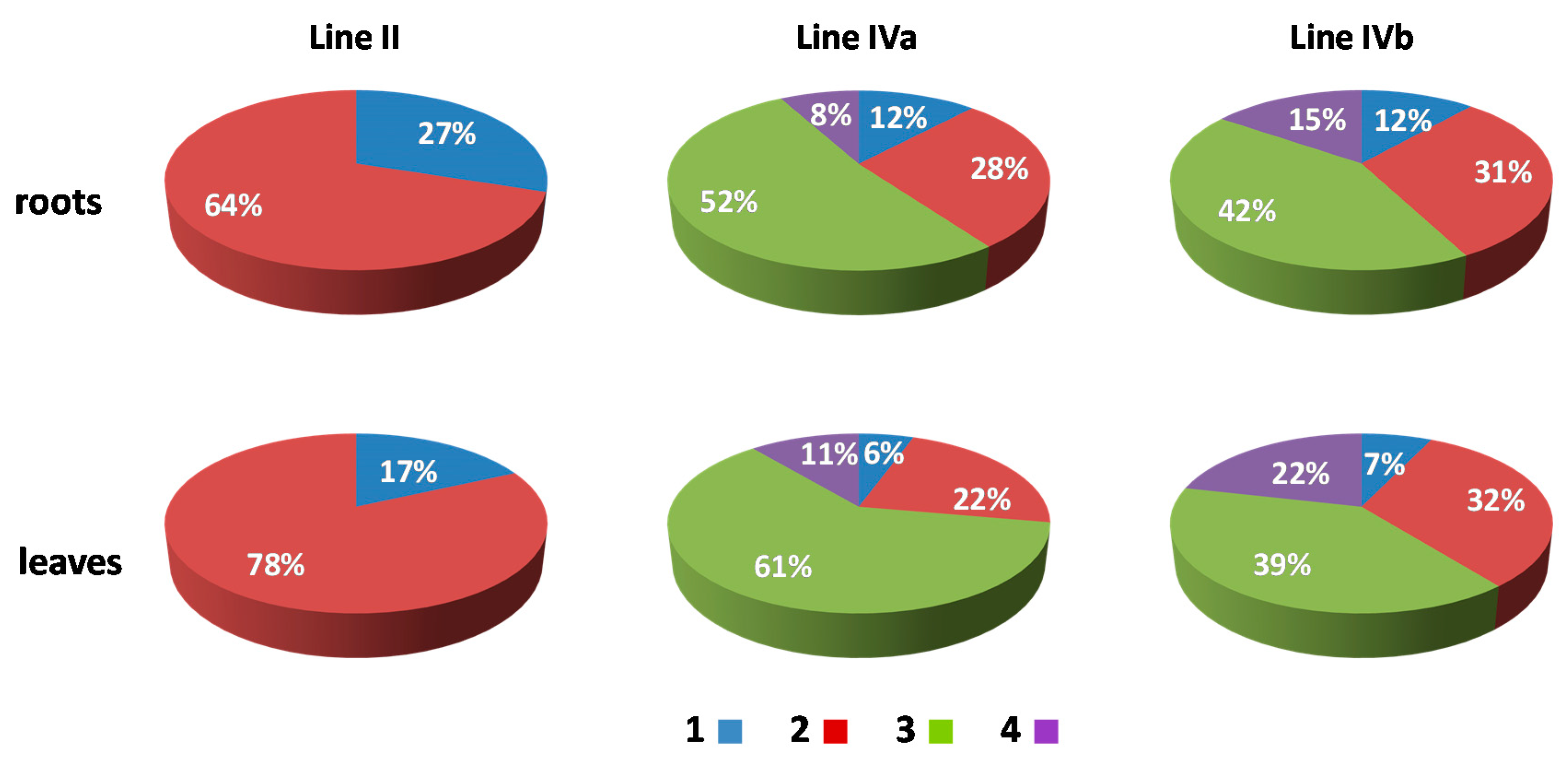
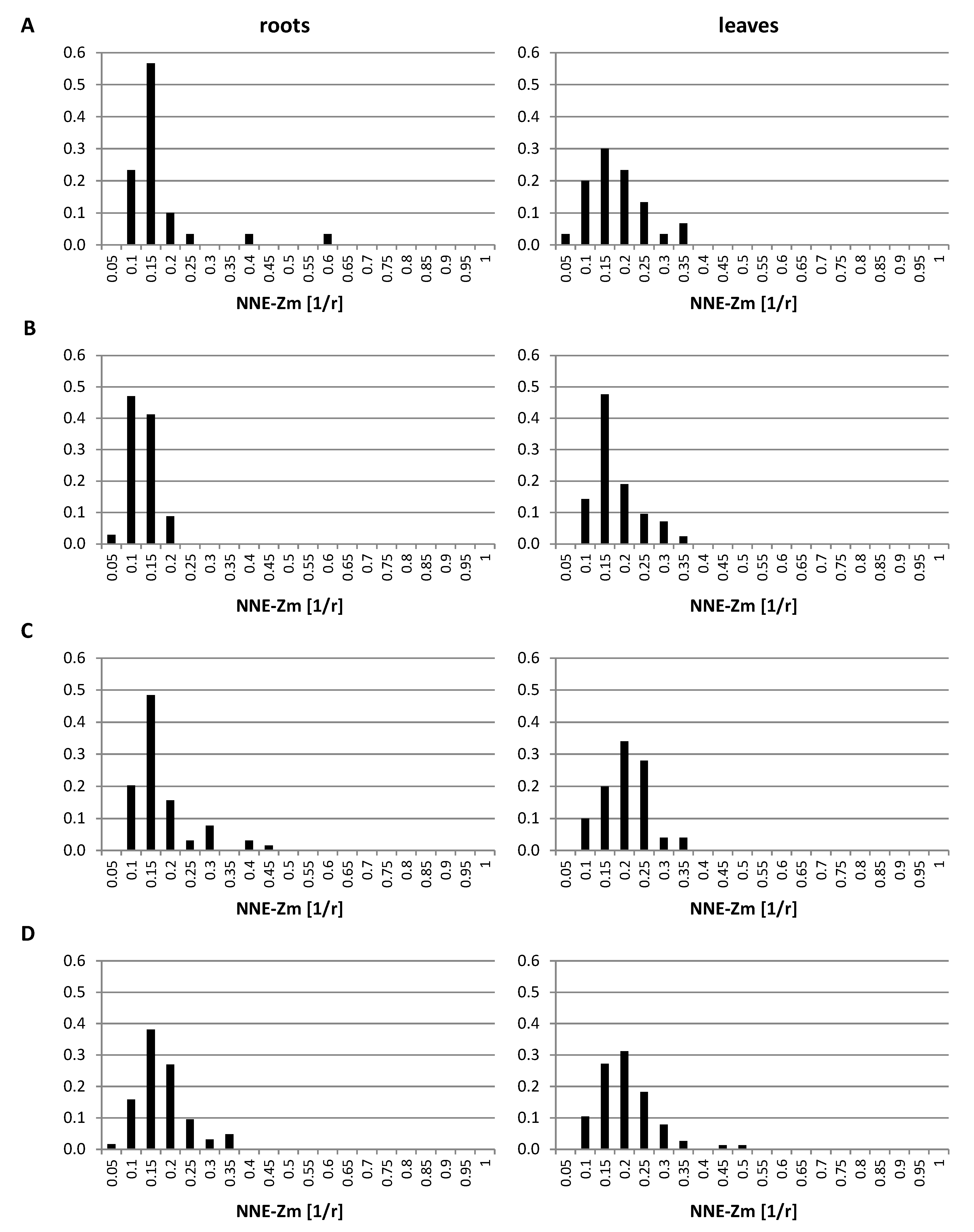
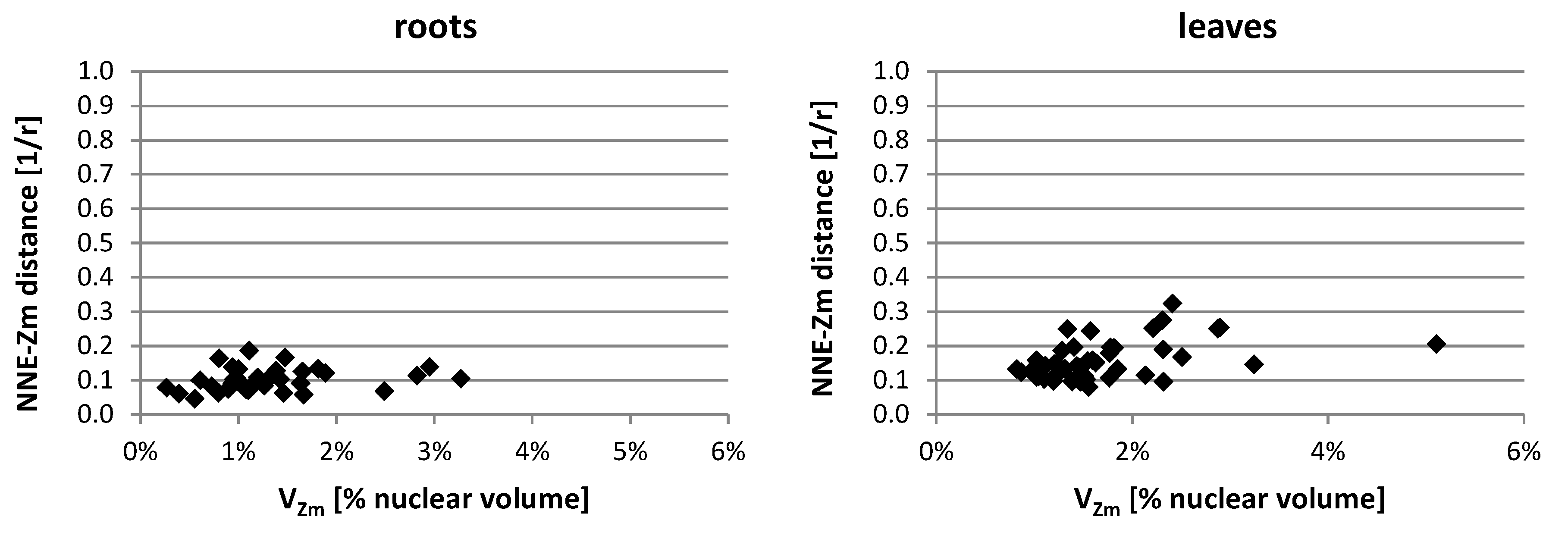
2.3. Positioning of Maize Chromatin-Occupied Regions
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Chromosome Preparations
4.3. Isolated Nuclei Preparations
4.4. DNA Extraction and PCR Analysis for Chromosome Identification
4.5. DNA Labeling and In Situ Hybridization
4.6. Image Acquisition and Analysis
4.7. Statistical Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BAC | Bacterial Artificial Chromosome |
| CT | Chromosome Territory |
| FISH | Fluorescence In Situ Hybridization |
| GISH | Genomic In Situ Hybridization |
| NNE | Nearest Nuclear Edge |
| NOR | Nucleolar Organizing Region |
| OMA | Oat × Maize Addition line |
| SSR | Simple Sequence Repeat |
Appendix A

References
- Cremer, T.; Cremer, M. Chromosome territories. Cold Spring Harb. Perspect. Biol. 2010, 2, a003889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meaburn, K.J.; Misteli, T. Cell biology: Chromosome territories. Nature 2007, 445, 379–781. [Google Scholar] [CrossRef] [PubMed]
- Branco, M.R.; Pombo, A. Intermingling of chromosome territories in interphase suggests role in translocations and transcription-dependent associations. PLoS Biol. 2006, 4, e138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visser, A.E.; Jaunin, F.; Fakan, S.; Aten, J.A. High resolution analysis of interphase chromosome domains. J. Cell Sci. 2000, 113, 2585–2593. [Google Scholar] [PubMed]
- Heard, E.; Bickmore, W. The ins and outs of gene regulation and chromosome territory organisation. Curr. Opin. Cell Biol. 2007, 19, 311–316. [Google Scholar] [CrossRef]
- Misteli, T.; Soutoglou, E. The emerging role of nuclear architecture in DNA repair and genome maintenance. Nat. Rev. Mol. Cell Biol. 2009, 10, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Boyle, S.; Gilchrist, S.; Bridger, J.M.; Mahy, N.L.; Ellis, J.A.; Bickmore, W.A. The spatial organization of human chromosomes within the nuclei of normal and emerin-mutant cells. Hum. Mol. Genet. 2001, 10, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Bolzer, A.; Kreth, G.; Solovei, I.; Koehler, D.; Saracoglu, K.; Fauth, C.; Muller, S.; Eils, R.; Cremer, C.; Speicher, M.R.; et al. Three-dimensional maps of all chromosomes in human male fibroblast nuclei and prometaphase rosettes. PLoS Biol. 2005, 3, e157. [Google Scholar] [CrossRef] [Green Version]
- Hepperger, C.; Mannes, A.; Merz, J.; Peters, J.; Dietzel, S. Three-dimensional positioning of genes in mouse cell nuclei. Chromosoma 2008, 117, 535–551. [Google Scholar] [CrossRef] [Green Version]
- Chandley, A.C.; Speed, R.M.; Leitch, A.R. Different distributions of homologous chromosomes in adult human Sertoli cells and in lymphocytes signify nuclear differentiation. J. Cell Sci. 1996, 109, 773–776. [Google Scholar]
- Parada, L.A.; McQueen, P.G.; Misteli, T. Tissue-specific spatial organization of genomes. Genome Biol. 2004, 5, R44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federico, C.; Scavo, C.; Cantarella, C.D.; Motta, S.; Saccone, S.; Bernardi, G. Gene-rich and gene-poor chromosomal regions have different locations in the interphase nuclei of cold-blooded vertebrates. Chromosoma 2006, 115, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, H.; Muller, S.; Neusser, M.; von Hase, J.; Calcagno, E.; Cremer, M.; Solovei, I.; Cremer, C.; Cremer, T. Evolutionary conservation of chromosome territory arrangements in cell nuclei from higher primates. Proc. Natl. Acad. Sci. USA 2002, 99, 4424–4429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, F.; Jiang, J. Non-Rabl patterns of centromere and telomere distribution in the interphase nuclei of plant cells. Chromosome Res. Int. J. Mol. Supramol. Evol. Asp. Chromosome Biol. 1998, 6, 551–558. [Google Scholar] [CrossRef]
- Baroux, C.; Pecinka, A.; Fuchs, J.; Kreth, G.; Schubert, I.; Grossniklaus, U. Non-random chromosome arrangement in triploid endosperm nuclei. Chromosoma 2017, 126, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Prieto, P.; Santos, A.P.; Moore, G.; Shaw, P. Chromosomes associate premeiotically and in xylem vessel cells via their telomeres and centromeres in diploid rice (Oryza sativa). Chromosoma 2004, 112, 300–307. [Google Scholar] [CrossRef] [Green Version]
- Idziak, D.; Robaszkiewicz, E.; Hasterok, R. Spatial distribution of centromeres and telomeres at interphase varies among Brachypodium species. J. Exp. Bot. 2015, 66, 6623–6634. [Google Scholar] [CrossRef] [Green Version]
- Dumur, T.; Duncan, S.; Graumann, K.; Desset, S.; Randall, R.S.; Scheid, O.M.; Prodanov, D.; Tatout, C.; Baroux, C. Probing the 3D architecture of the plant nucleus with microscopy approaches: Challenges and solutions. Nucleus 2019, 10, 181–212. [Google Scholar] [CrossRef] [Green Version]
- Leitch, A.R.; Mosgoller, W.; Schwarzacher, T.; Bennett, M.D.; Heslop-Harrison, J.S. Genomic in situ hybridization to sectioned nuclei shows chromosome domains in grass hybrids. J. Cell Sci. 1990, 95, 335–341. [Google Scholar]
- Han, J.; Zhou, B.; Shan, W.; Yu, L.; Wu, W.; Wang, K. A and D genomes spatial separation at somatic metaphase in tetraploid cotton: Evidence for genomic disposition in a polyploid plant. Plant J. 2015, 84, 1167–1177. [Google Scholar] [CrossRef]
- Leitch, A.R.; Schwarzacher, T.; Mosgoller, W.; Bennett, M.D.; Heslop-Harrison, J.S. Parental genomes are separated throughout the cell cycle in a plant hybrid. Chromosoma 1991, 101, 206–213. [Google Scholar] [CrossRef]
- Kolackova, V.; Pernickova, K.; Vrana, J.; Duchoslav, M.; Jenkins, G.; Phillips, D.; Turkosi, E.; Samajova, O.; Sedlarova, M.; Samaj, J.; et al. Nuclear Disposition of Alien Chromosome Introgressions into Wheat and Rye Using 3D-FISH. Int. J. Mol. Sci. 2019, 20, 4143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gernand, D.; Rutten, T.; Varshney, A.; Rubtsova, M.; Prodanovic, S.; Bruss, C.; Kumlehn, J.; Matzk, F.; Houben, A. Uniparental chromosome elimination at mitosis and interphase in wheat and pearl millet crosses involves micronucleus formation, progressive heterochromatinization, and DNA fragmentation. Plant Cell 2005, 17, 2431–2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kynast, R.G.; Riera-Lizarazu, O.; Vales, M.I.; Okagaki, R.J.; Maquieira, S.B.; Chen, G.; Ananiev, E.V.; Odland, W.E.; Russell, C.D.; Stec, A.O.; et al. A complete set of maize individual chromosome additions to the oat genome. Plant Physiol. 2001, 125, 1216–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurie, D.A.; Bennett, M.D. The timing of chromosome elimination in hexaploid wheat x maize crosses. Genome 1989, 32, 953–961. [Google Scholar] [CrossRef]
- Riera-Lizarazu, O.; Rines, H.W.; Phillips, R.L. Cytological and molecular characterization of oat x maize partial hybrids. Appl. Genet. 1996, 93, 123–135. [Google Scholar] [CrossRef]
- Zenkteler, M.; Nitzsche, W. Wide hybridization experiments in cereals. Appl. Genet. 1984, 68, 311–315. [Google Scholar] [CrossRef]
- Soltis, P.S.; Marchant, D.B.; Van de Peer, Y.; Soltis, D.E. Polyploidy and genome evolution in plants. Curr. Opin. Genet. Dev. 2015, 35, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Gill, B.S. Different species-specific chromosome translocations in Triticum timopheevii and T. turgidum support the diphyletic origin of polyploid wheats. Chromosome Res. 1994, 2, 59–64. [Google Scholar] [CrossRef]
- Lim, K.Y.; Matyasek, R.; Kovarik, A.; Leitch, A.R. Genome evolution in allotetraploid Nicotiana. Biol. J. Linn. Soc. 2004, 82, 599–606. [Google Scholar] [CrossRef]
- Pernickova, K.; Kolackova, V.; Lukaszewski, A.J.; Fan, C.; Vrana, J.; Duchoslav, M.; Jenkins, G.; Phillips, D.; Samajova, O.; Sedlarova, M.; et al. Instability of Alien Chromosome Introgressions in Wheat Associated with Improper Positioning in the Nucleus. Int. J. Mol. Sci. 2019, 20, 1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rines, H.W.; Phillips, R.L.; Kynast, R.G.; Okagaki, R.J.; Galatowitsch, M.W.; Huettl, P.A.; Stec, A.O.; Jacobs, M.S.; Suresh, J.; Porter, H.L.; et al. Addition of individual chromosomes of maize inbreds B73 and Mo17 to oat cultivars Starter and Sun II: Maize chromosome retention, transmission, and plant phenotype. Appl. Genet. 2009, 119, 1255–1264. [Google Scholar] [CrossRef] [PubMed]
- Kynast, R.G.; Okagaki, R.J.; Rines, H.W.; Phillips, R.L. Maize individualized chromosome and derived radiation hybrid lines and their use in functional genomics. Funct. Integr. Genom. 2002, 2, 60–69. [Google Scholar] [CrossRef]
- Okagaki, R.J.; Kynast, R.G.; Livingston, S.M.; Russell, C.D.; Rines, H.W.; Phillips, R.L. Mapping maize sequences to chromosomes using oat-maize chromosome addition materials. Plant Physiol. 2001, 125, 1228–1235. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Yu, J.; Li, H.; Huang, W.; Xu, L.; Zhao, Y.; Zhang, T.; Xu, W.; Jiang, J.; Su, Z.; et al. Transcriptional and epigenetic adaptation of maize chromosomes in Oat-Maize addition lines. Nucleic Acids Res. 2018, 46, 5012–5028. [Google Scholar] [CrossRef] [PubMed]
- Kowles, R.V.; Walch, M.D.; Minnerath, J.M.; Bernacchi, C.J.; Stec, A.O.; Rines, H.W. Expression of C4 photosynthetic enzymes in oat–maize chromosome addition lines. Maydica 2008, 53, 69–78. [Google Scholar]
- Muehlbauer, G.J.; Riera-Lizarazu, O.; Kynast, R.G.; Martin, D.; Phillips, R.L.; Rines, H.W. A maize chromosome 3 addition line of oat exhibits expression of the maize homeobox gene liguleless3 and alteration of cell fates. Genome 2000, 43, 1055–1064. [Google Scholar] [CrossRef]
- Jin, W.; Melo, J.R.; Nagaki, K.; Talbert, P.B.; Henikoff, S.; Dawe, R.K.; Jiang, J. Maize centromeres: Organization and functional adaptation in the genetic background of oat. Plant Cell 2004, 16, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Wu, Y.; Zhang, W.; Dawe, R.K.; Jiang, J. Maize centromeres expand and adopt a uniform size in the genetic background of oat. Genome Res. 2014, 24, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Bass, H.W.; Riera-Lizarazu, O.; Ananiev, E.V.; Bordoli, S.J.; Rines, H.W.; Phillips, R.L.; Sedat, J.W.; Agard, D.A.; Cande, W.Z. Evidence for the coincident initiation of homolog pairing and synapsis during the telomere-clustering (bouquet) stage of meiotic prophase. J. Cell Sci. 2000, 113, 1033–1042. [Google Scholar]
- Skrzypek, E.; Warzecha, T.; Noga, A.; Warchol, M.; Czyczylo-Mysza, I.; Dziurka, K.; Marcinska, I.; Kaploniak, K.; Sutkowska, A.; Nita, Z.; et al. Complex characterization of oat (Avena sativa L.) lines obtained by wide crossing with maize (Zea mays L.). PeerJ 2018, 6, e5107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Ma, C.; Tian, X.; Zhu, C.; Wang, G.; Lv, Y.; Friebe, B.; Li, H.; Liu, W. Genome-wide impacts of alien chromatin introgression on wheat gene transcriptions. Sci. Rep. 2020, 10, 4801. [Google Scholar] [CrossRef] [PubMed]
- Kynast, R.G.; Davis, D.W.; Phillips, R.L.; Rines, H.W. Gamete formation via meiotic nuclear restitution generates fertile amphiploid F1 (oat × maize) plants. Sex. Plant Reprod. 2012, 25, 111–122. [Google Scholar] [CrossRef]
- Tolley, B.J.; Sage, T.L.; Langdale, J.A.; Hibberd, J.M. Individual maize chromosomes in the C(3) plant oat can increase bundle sheath cell size and vein density. Plant Physiol. 2012, 159, 1418–1427. [Google Scholar] [CrossRef] [Green Version]
- Kynast, R.G.; Okagaki, R.J.; Galatowitsch, M.W.; Granath, S.R.; Jacobs, M.S.; Stec, A.O.; Rines, H.W.; Phillips, R.L. Dissecting the maize genome by using chromosome addition and radiation hybrid lines. Proc. Natl. Acad. Sci. USA 2004, 101, 9921–9926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pernickova, K.; Linc, G.; Gaal, E.; Kopecky, D.; Samajova, O.; Lukaszewski, A.J. Out-of-position telomeres in meiotic leptotene appear responsible for chiasmate pairing in an inversion heterozygote in wheat (Triticum aestivum L.). Chromosoma 2019, 128, 31–39. [Google Scholar] [CrossRef]
- Naranjo, T. Dynamics of rye telomeres in a wheat background during early meiosis. Cytogenet Genome Res. 2014, 143, 60–68. [Google Scholar] [CrossRef]
- Naranjo, T. Variable Patterning of Chromatin Remodeling, Telomere Positioning, Synapsis, and Chiasma Formation of Individual Rye Chromosomes in Meiosis of Wheat-Rye Additions. Front. Plant Sci. 2018, 9, 880. [Google Scholar] [CrossRef]
- Oleszczuk, S.; Rabiza-Swider, J.; Zimny, J.; Lukaszewski, A.J. Aneuploidy among androgenic progeny of hexaploid triticale (XTriticosecale Wittmack). Plant Cell Rep. 2011, 30, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Misteli, T. Beyond the sequence: Cellular organization of genome function. Cell 2007, 128, 787–800. [Google Scholar] [CrossRef] [Green Version]
- Heride, C.; Ricoul, M.; Kieu, K.; von Hase, J.; Guillemot, V.; Cremer, C.; Dubrana, K.; Sabatier, L. Distance between homologous chromosomes results from chromosome positioning constraints. J. Cell Sci. 2010, 123, 4063–4075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berr, A.; Pecinka, A.; Meister, A.; Kreth, G.; Fuchs, J.; Blattner, F.R.; Lysak, M.A.; Schubert, I. Chromosome arrangement and nuclear architecture but not centromeric sequences are conserved between Arabidopsis thaliana and Arabidopsis lyrata. Plant J. 2006, 48, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Pecinka, A.; Schubert, V.; Meister, A.; Kreth, G.; Klatte, M.; Lysak, M.A.; Fuchs, J.; Schubert, I. Chromosome territory arrangement and homologous pairing in nuclei of Arabidopsis thaliana are predominantly random except for NOR-bearing chromosomes. Chromosoma 2004, 113, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Aragon-Alcaide, L.; Reader, S.; Beven, A.; Shaw, P.; Miller, T.; Moore, G. Association of homologous chromosomes during floral development. Curr. Biol. 1997, 7, 905–908. [Google Scholar] [CrossRef] [Green Version]
- Robaszkiewicz, E.; Idziak-Helmcke, D.; Tkacz, M.A.; Chrominski, K.; Hasterok, R. The arrangement of Brachypodium distachyon chromosomes in interphase nuclei. J. Exp. Bot. 2016, 67, 5571–5583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danilova, T.V.; Birchler, J.A. Integrated cytogenetic map of mitotic metaphase chromosome 9 of maize: Resolution, sensitivity, and banding paint development. Chromosoma 2008, 117, 345–356. [Google Scholar] [CrossRef]
- Kato, A.; Lamb, J.C.; Birchler, J.A. Chromosome painting using repetitive DNA sequences as probes for somatic chromosome identification in maize. Proc. Natl. Acad. Sci. USA 2004, 101, 13554–13559. [Google Scholar] [CrossRef] [Green Version]
- Albert, P.S.; Zhang, T.; Semrau, K.; Rouillard, J.M.; Kao, Y.H.; Wang, C.R.; Danilova, T.V.; Jiang, J.; Birchler, J.A. Whole-chromosome paints in maize reveal rearrangements, nuclear domains, and chromosomal relationships. Proc. Natl. Acad. Sci. USA 2019, 116, 1679–1685. [Google Scholar] [CrossRef] [Green Version]
- Cowan, C.R.; Carlton, P.M.; Cande, W.Z. The polar arrangement of telomeres in interphase and meiosis. Rabl organization and the bouquet. Plant Physiol. 2001, 125, 532–538. [Google Scholar] [CrossRef] [Green Version]
- Rabl, C. Über Zelltheilung. Morphol. Jahrb. 1885, 10, 214–330. [Google Scholar]
- Fransz, P.; De Jong, J.H.; Lysak, M.; Castiglione, M.R.; Schubert, I. Interphase chromosomes in Arabidopsis are organized as well defined chromocenters from which euchromatin loops emanate. Proc. Natl. Acad. Sci. USA 2002, 99, 14584–14589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, H.; Zhou, Z.; Wang, Y.; Wang, J.; Kallgren, S.P.; Kurchuk, T.; Miller, E.A.; Chang, F.; Jia, S. Csi1 links centromeres to the nuclear envelope for centromere clustering. J. Cell Biol. 2012, 199, 735–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.S.; Armstrong, K.C.; Fedak, G.; Ho, K.; Park, N.I. A microsatellite sequence from the rice blast fungus (Magnaporthe grisea) distinguishes between the centromeres of Hordeum vulgare and H. bulbosum in hybrid plants. Genome 2002, 45, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Carvalho, C.R.; Clarindo, W.R. Updating the maize karyotype by chromosome DNA sizing. PLoS ONE 2018, 13, e0190428. [Google Scholar] [CrossRef] [PubMed]
- Idziak-Helmcke, D.; Betekhtin, A. Methods for Cytogenetic Chromosome Barcoding and Chromosome Painting in Brachypodium distachyon and Its Relative Species. Methods Mol. Biol. 2018, 1667, 1–19. [Google Scholar]
- Dolezel, J.; Binaroca, P.; Lucretti, S. Analysis of nuclear DNA content in plant cells by flow cytometry. Biol. Plant. 1989, 31, 113–120. [Google Scholar] [CrossRef]
- Jenkins, G.; Hasterok, R. BAC ‘landing’ on chromosomes of Brachypodium distachyon for comparative genome alignment. Nat. Protoc. 2007, 2, 88–98. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype of the Addition Line | Symbol of the Addition Line | No. of Maize Chromosomes Added to the Oat Genome | Chromosome ID |
|---|---|---|---|
| STH 6.8636 | I | 1 | 5 |
| STH 5.8436 b | II | 2 | 3, 8 |
| STH 6.8661 | IVa | 4 | 3, 9 |
| STH 4.4690 p | IVb | 4 | 1, 2 |
| Maize Chromosome | SSR Marker | Maize Chromosome | SSR Marker |
|---|---|---|---|
| 1 | p-bnlg421 | 6 | phi070 |
| 2 | p-bnlg125 | 7 | phi112 |
| 3 | p-phi047 | 8 | phi080 |
| 4 | p-phi093, p-blng490 | 9 | phi032 |
| 5 | p-nc130 | 10 | phi059 |
| Parameter | I Roots | I Leaves | II Roots | II Leaves | IVa Roots | IVa Leaves | IVb Roots | IVb Leaves |
|---|---|---|---|---|---|---|---|---|
| No. of nuclei | 30 | 30 | 20 | 23 | 25 | 18 | 25 | 28 |
| Vnuc _max [µm3] | 1401 | 1849 | 1693 | 1176 | 1311 | 732 | 1375 | 857 |
| Vnuc_min [µm3] | 533 | 611 | 314 | 527 | 268 | 238 | 576 | 372 |
| Vnuc_av [µm3] | 1023 | 966 | 1031 | 795 | 570 | 463 | 939 | 544 |
| Vnuc_med [µm3] | 1050 | 873 | 1032 | 753 | 412 | 451 | 875 | 492 |
| Vnuc_SD [µm3] | 193 | 344 | 413 | 152 | 285 | 105 | 254 | 116 |
| VZm_max [µm3] | 36.50 | 33.60 | 32.70 | 37.0 | 18.10 | 42.90 | 67.20 | 47.70 |
| VZm_min [µm3] | 6.49 | 6.57 | 3.09 | 6.12 | 0.97 | 1.16 | 4.14 | 2.45 |
| VZm_av [µm3] | 19.31 | 14.55 | 12.99 | 13.67 | 7.20 | 7.65 | 23.96 | 12.16 |
| VZm med [µm3] | 18.35 | 13.45 | 12.00 | 13.00 | 6.46 | 5.43 | 19.30 | 8.57 |
| VZm_SD [µm3] | 7.66 | 6.47 | 7.55 | 5.95 | 3.82 | 6.66 | 15.62 | 9.12 |
| VZm_max [%] | 3.53% | 3.33% | 3.27% | 5.11% | 3.45% | 5.86% | 9.44% | 8.16% |
| VZm_min [%] | 0.61% | 0.73% | 0.27% | 0.82% | 0.14% | 0.33% | 0.50% | 0.55% |
| VZm_av [%] | 1.94% | 1.55% | 1.33% | 1.73% | 1.35% | 1.64% | 2.59% | 2.24% |
| VZm_med [%] | 1.86% | 1.33% | 1.15% | 1.54% | 1.28% | 1.26% | 1.94% | 1.76% |
| VZm_SD [%] | 0.80% | 0.61% | 0.70% | 0.79% | 0.66% | 1.11% | 1.78% | 1.52% |
| Vtot_max [µm3] | 36.50 | 33.60 | 43.60 | 46.10 | 40.85 | 42.90 | 113.40 | 71.50 |
| Vtot_min [µm3] | 6.49 | 6.57 | 7.57 | 13.60 | 6.67 | 8.01 | 27.81 | 15.70 |
| Vtot_av [µm3] | 19.75 | 14.55 | 22.08 | 24.97 | 18.44 | 21.26 | 60.38 | 33.45 |
| Vtot_med [µm3] | 18.35 | 13.45 | 19.42 | 24.50 | 17.94 | 18.59 | 55.00 | 31.83 |
| Vtot_SD [µm3] | 7.66 | 6.47 | 9.30 | 8.27 | 7.41 | 9.39 | 22.56 | 13.63 |
| Vtot_max [%] | 3.53% | 3.33% | 3.55% | 5.39% | 5.86% | 7.94% | 9.44% | 10.56% |
| Vtot_min [%] | 0.61% | 0.73% | 1.19% | 1.89% | 1.68% | 2.42% | 2.85% | 3.18% |
| Vtot_av [%] | 1.93% | 1.55% | 2.26% | 3.17% | 3.47% | 4.56% | 6.53% | 6.15% |
| Vtot_med [%] | 1.86% | 1.33% | 2.21% | 2.90% | 3.45% | 4.16% | 6.18% | 5.62% |
| Vtot_SD [%] | 0.80% | 0.61% | 0.68% | 0.98% | 1.05% | 1.64% | 1.83% | 1.92% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Idziak-Helmcke, D.; Warzecha, T.; Sowa, M.; Warchoł, M.; Dziurka, K.; Czyczyło-Mysza, I.; Skrzypek, E. 3-D Nucleus Architecture in Oat × Maize Addition Lines. Int. J. Mol. Sci. 2020, 21, 4280. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124280
Idziak-Helmcke D, Warzecha T, Sowa M, Warchoł M, Dziurka K, Czyczyło-Mysza I, Skrzypek E. 3-D Nucleus Architecture in Oat × Maize Addition Lines. International Journal of Molecular Sciences. 2020; 21(12):4280. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124280
Chicago/Turabian StyleIdziak-Helmcke, Dominika, Tomasz Warzecha, Marta Sowa, Marzena Warchoł, Kinga Dziurka, Ilona Czyczyło-Mysza, and Edyta Skrzypek. 2020. "3-D Nucleus Architecture in Oat × Maize Addition Lines" International Journal of Molecular Sciences 21, no. 12: 4280. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124280