Meta-Analysis of Gene Expressions in Testicular Germ Cell Tumor Histologies


1
Center of Tobacco Control Research, DK-5230 Odense M, Denmark
2
Facultad de Ciencias Informaticas, Universidad Tecnica de Manabi, Portoviejo 130105, Ecuador
3
Departamento de Ingenieria Informatica, Universidad de Santiago de Chile, Santiago 9170020, Chile
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(12), 4487; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124487
Submission received: 22 May 2020
/
Revised: 18 June 2020
/
Accepted: 19 June 2020
/
Published: 24 June 2020
(This article belongs to the Special Issue Germ Cells and Genitourinary Cancers)
Abstract
:There is no consensus as to how a precursor lesion, germ cell neoplasia in situ (GCNIS), develops into the histologic types of testicular germ cell tumor type II (TGCT). The present meta-analysis examined RNA expressions of 24 candidate genes in three datasets. They included 203 samples of normal testis (NT) and histologic types of TGCT. The Fisher’s test for combined p values was used for meta-analysis of the RNA expressions in the three datasets. The histologic types differed in RNA expression of PRAME, KIT, SOX17, NANOG, KLF4, POU5F1, RB1, DNMT3B, and LIN28A (p < 0.01). The histologic types had concordant differences in RNA expression of the genes in the three datasets. Eight genes had overlap with a high RNA expression in at least two histologic types. In contrast, only seminoma (SE) had a high RNA expression of KLF4 and only embryonal carcinoma (EC) had a high RNA expression of DNMT3B. In conclusion, the meta-analysis showed that the development of the histologic types of TGCT was driven by changes in RNA expression of candidate genes. According to the RNA expressions of the ten genes, TGCT develops from NT over GCNIS, SE, EC, to the differentiated types of TGCT.
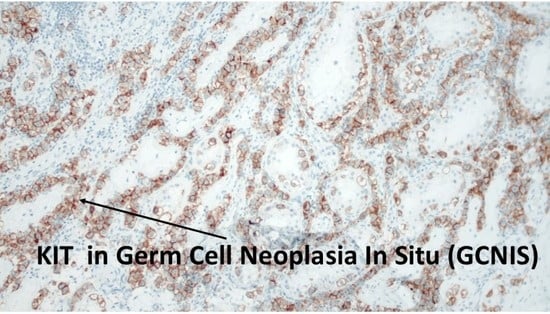
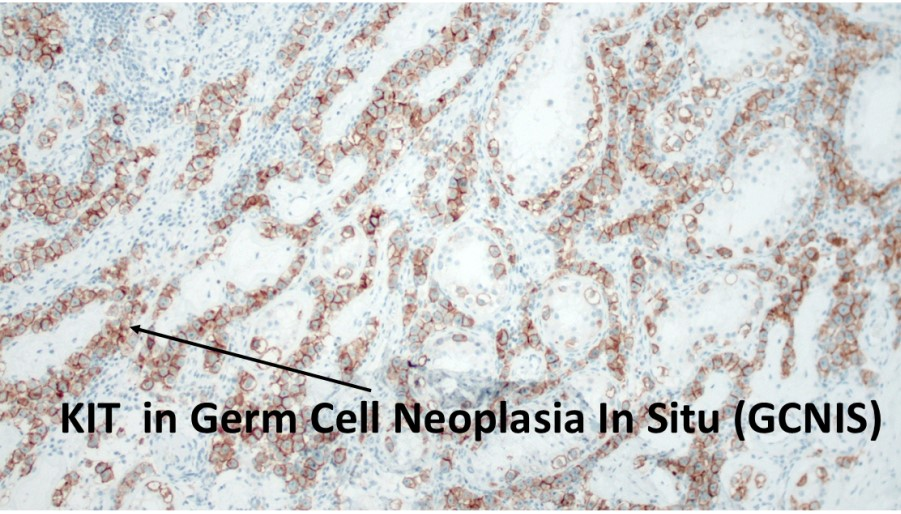
1. Introduction
Testicular germ cell tumor type II (TGCT) is the most frequent malignancy in young adult men. Worldwide, oncologists follow the TNM (T for primary tumor, N for regional lymph node metastases, M for distant metastases) classification that separates TGCT into seminoma and nonseminomatous germ cell tumors (NSGCT) [1].
The WHO classification of tumors of the urogenital system and male genital organs 2016 [2] acknowledges that TGCT develops from a common precursor lesion, germ cell neoplasia in situ (GCNIS). The WHO classification is based on extensive previous research. The research started in 1896, in which Wilms reported that TGCT was derived from normal testicular germ cells [3].
In 1950, Dixon and Moore described that TGCT developed along two lines, i.e., one line for seminoma and the other line for NSCGT [4]. With regards to NSGCT, embryonal carcinoma (EC) could differentiate into teratoma (TER) and choriocarcinoma (CC). In 1959, Teilum described a separate histologic type of NSGCT, YST (also called endodermal sinus tumor) [5]. In 1990, de Jong summarized studies that showed TGCT had a characteristic isochromosome of the short arm of chromosome 12, 12p, and i(12p) [6].
In 2004, in a meta-analysis of genes in TGCT, von Eyben pointed out that undifferentiated types of TGCT had high levels of CCND2 and low levels of RB1 which combined, programed the malignant germ cells for a high proliferation [7]. A meta-analysis by Alagaratnam, in 2011, pointed out that 92 genes had roles for the pathogenesis of TGCT [8]. From 2012 to 2016, Nettersheim in studies of xenografts of TCam2 to nude mice showed that seminoma (SE) can develop into EC [9].
After the WHO classification in 2016, an immunohistochemical study, in 2017, showed that precursor lesions for TGCT had a seminomatous immunophenotype [10]. The Genome Cancer Atlas (TGCA) project, in 2018, found that TGCT did not have mandatory somatic mutations [11].
In general, solid tumors develop as a result of an accumulation of mutations or epigenetic changes in oncogenes or tumor suppressor gene. As TGCT does not have mandatory mutations, the development of the histologic types should be caused by regulation of gene expression of candidate genes. To further study how TGCT develops, we aimed to investigate whether the histologic types of TGCT differed significantly in RNA expression of candidate genes.
2. Results
2.1. Comparisons of Three Datasets
Our meta-analysis examined RNA expression of 24 candidate genes in three datasets of TGCT. The three datasets are publicly available [12,13,14]. Previous publications had reported that the gene products of the 24 candidate genes were significant in the pathogenesis or treatment of TGCT.
The three datasets were from studies that differed with respect to time, country, and methodology for the RNA measurements. Nevertheless, the histologic types had concordant RNA expressions of eight significant genes. Therefore, our meta-analysis combined the evaluation of the three datasets.
2.2. Significant Genes
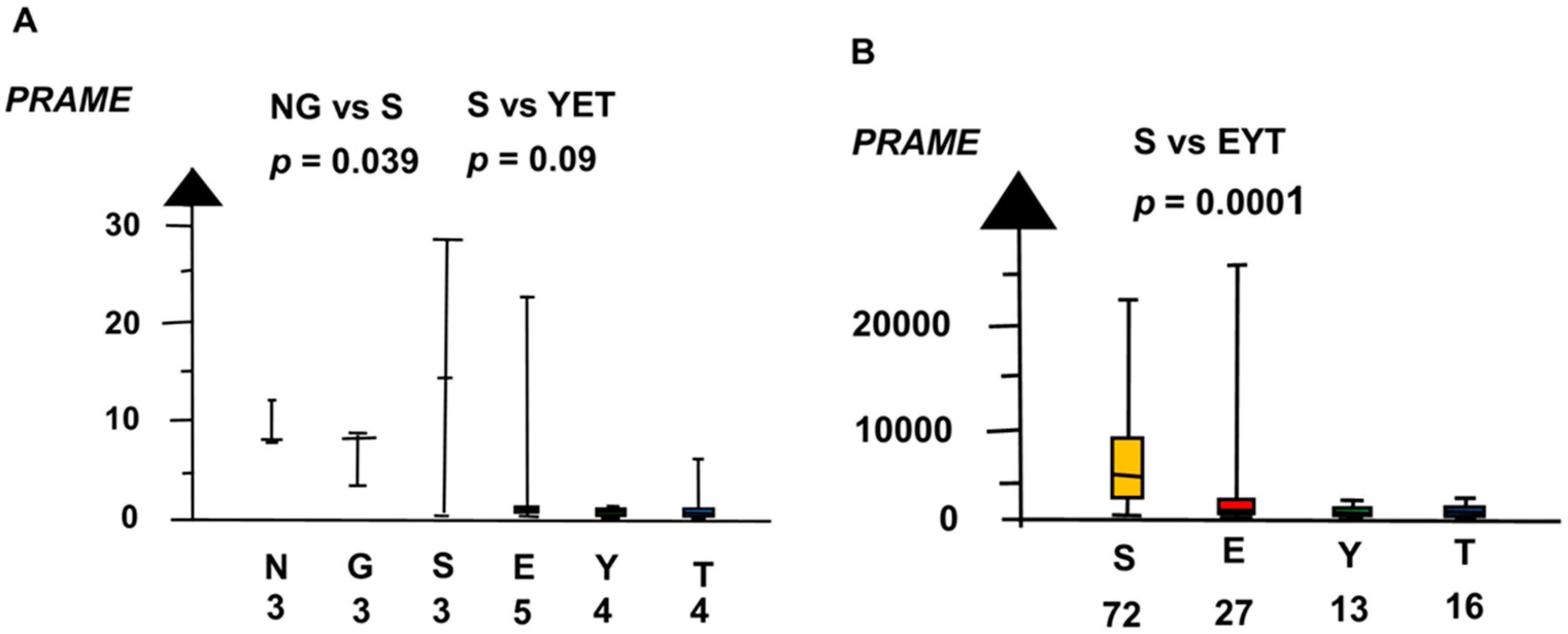
Our meta-analysis found the histologic types had overlaps and differences in RNA expressions of the candidate genes. Normal testis (NT), GCNIS, and SE had overlap with high RNA expression of PRAME, as shown in Figure 1. In the second microarray, NT, GCNIS, and SE combined had a sixteen times higher RNA expression of PRAME than EC, YST, and TER combined (median 8.4 vs. 0.4, p = 0.003, Kruskal–Wallis test).
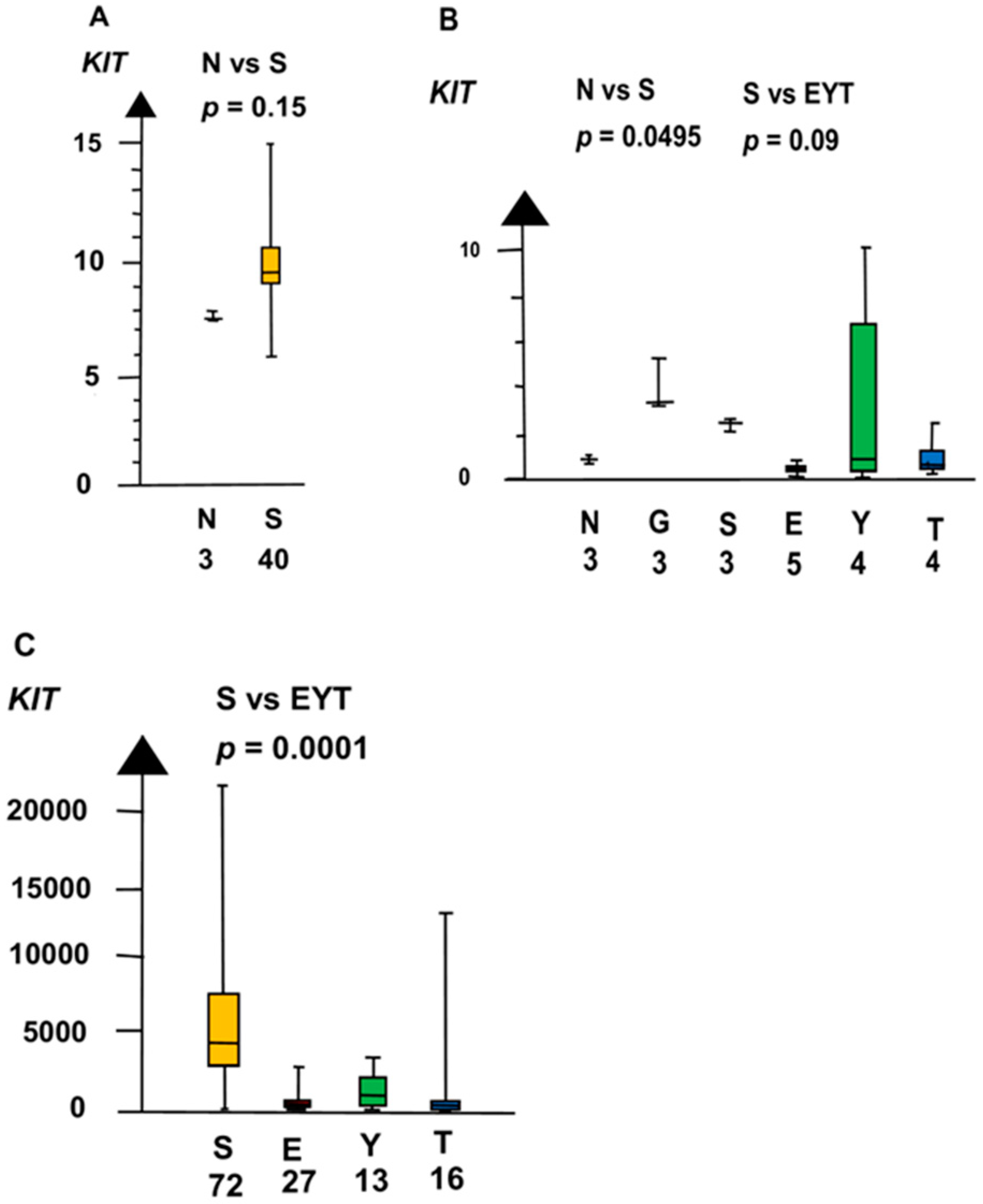
GCNIS had a higher RNA expression than NT of three genes, i.e., KIT, SOX17, and NANOG. In the second microarray, GCNIS and SE combined had a four times higher RNA expression of KIT than NT (median 4.3 vs. 0.89, p = 0.02, Kruskal–Wallis test), as shown in Figure 2.
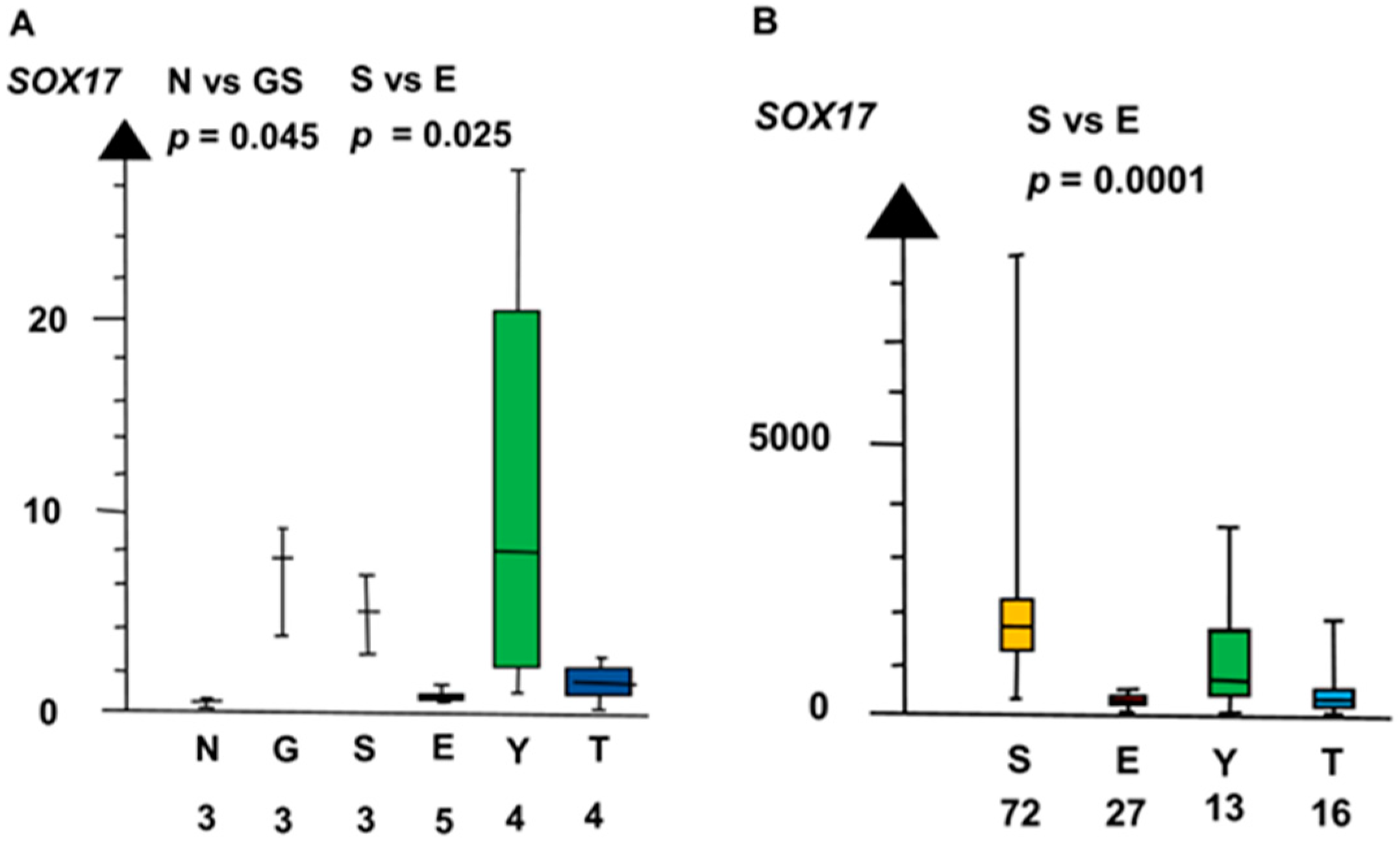
GCNIS and SE combined had a four times higher RNA expression of SOX17 than EC, YST, and TER combined (median 3.9 vs. 1.0, p = 0.023, Kruskal–Wallis test), as shown in Figure 3.
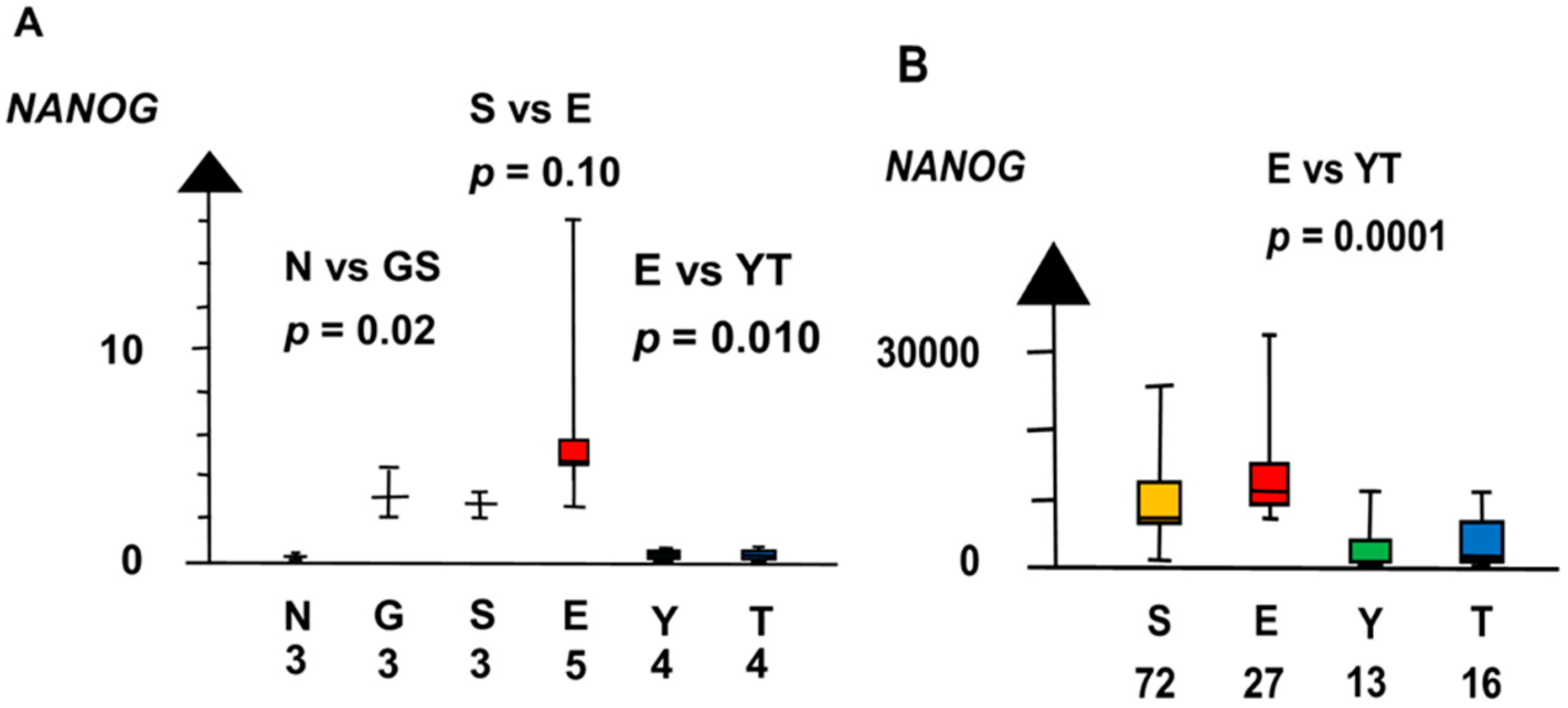
GCNIS and SE had a ten times higher RNA expression of NANOG than NT (median 2.9 vs. 0.29, p = 0.02, Kruskal–Wallis test), as shown in Figure 4. NANOG differed stepwise among histologic types. SE and EC combined had a three times higher RNA expression of NANOG than NT and GCNIS combined (median 3.8 vs. 1.2, p = 0.039, Kruskal–Wallis test).
SE had a set of overexpressed genes that differed from that in GCNIS. The RNA expression of CCND2, KLF4, and POU5F1 was high in SE but low in GCNIS. Furthermore, SE had a downregulated tumor suppressor gene, RB1, not downregulated in GCNIS. In contrast, both GCNIS and SE had high RNA expression of KIT, SOX17, and NANOG.
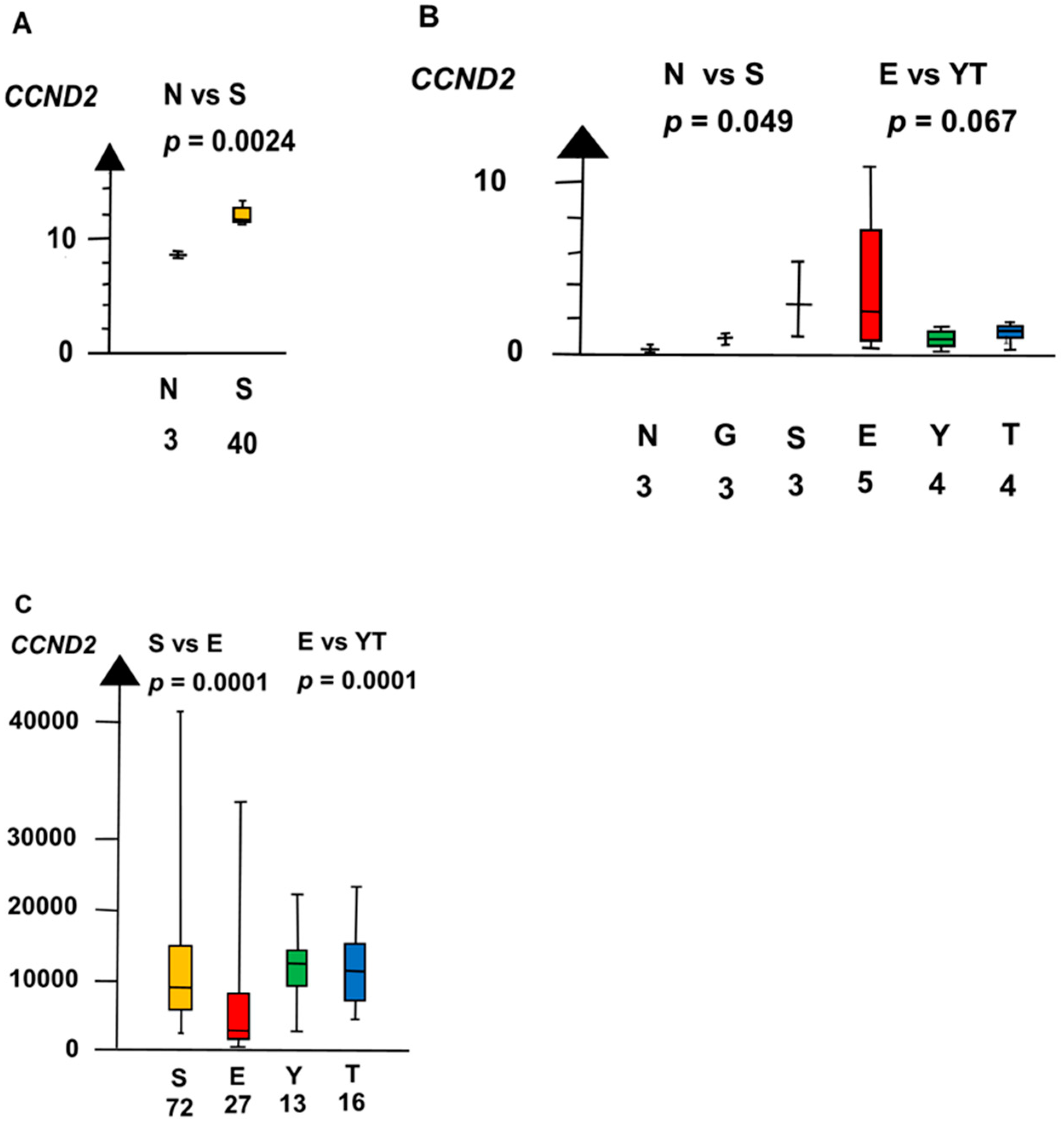
In the second microarray, SE had a twelve times higher RNA expression of CCND2 than NT (median 3.1 vs. 0.26, p = 0.0495, Kruskal–Wallis test), as shown in Figure 5.
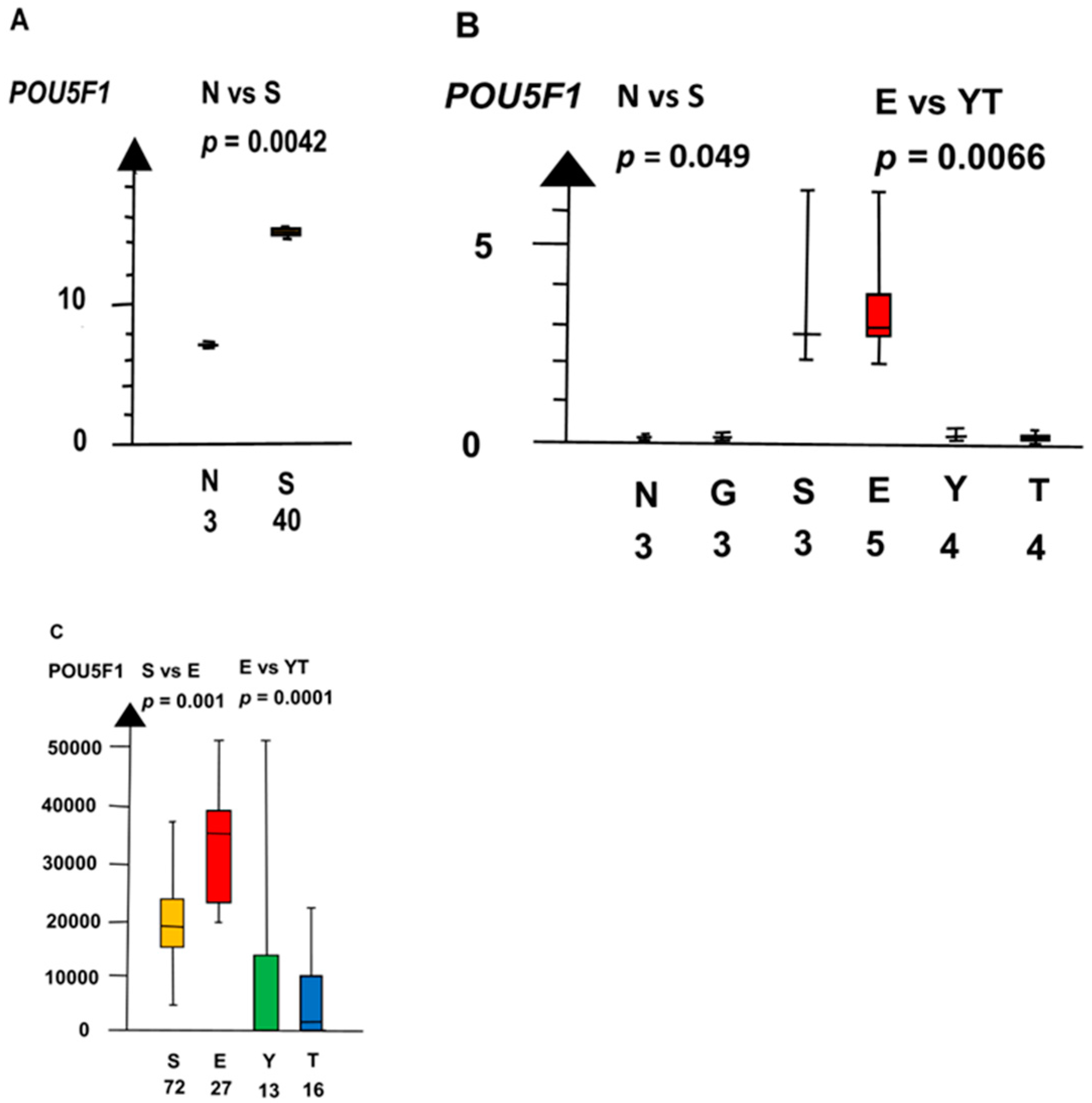
SE and EC combined had a four times higher RNA expression of POU5F1 than NT and GCNIS combined (median 3.5 vs. 0.72, p = 0.0045, Kruskal–Wallis test), as shown in Figure 6.
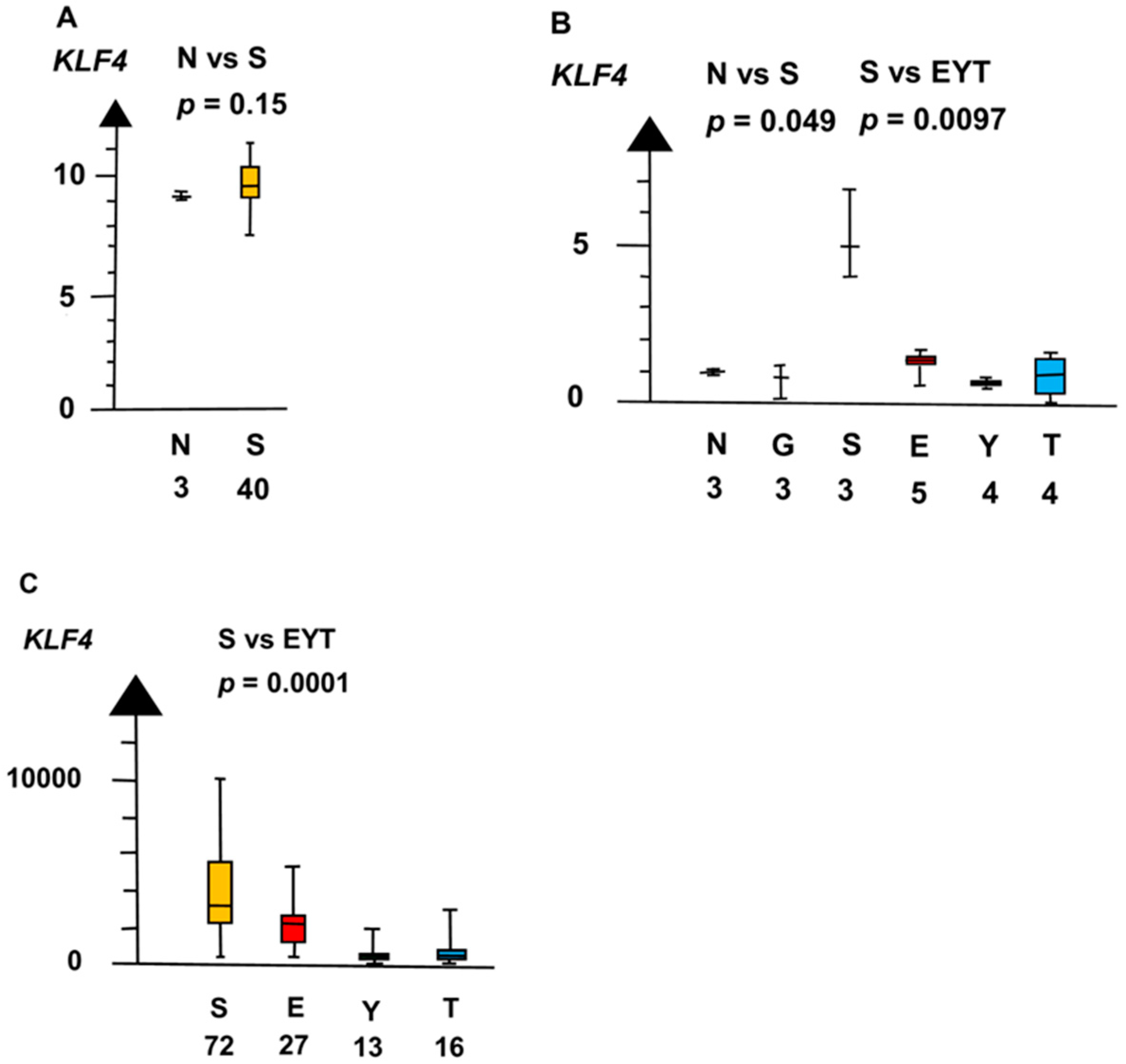
SE and EC had a seven times higher RNA expression of KLF4 than all other histologic types combined (median 4.9 vs. 0.71, p = 0.0073, Kruskal–Wallis test), as indicated in Figure 7.
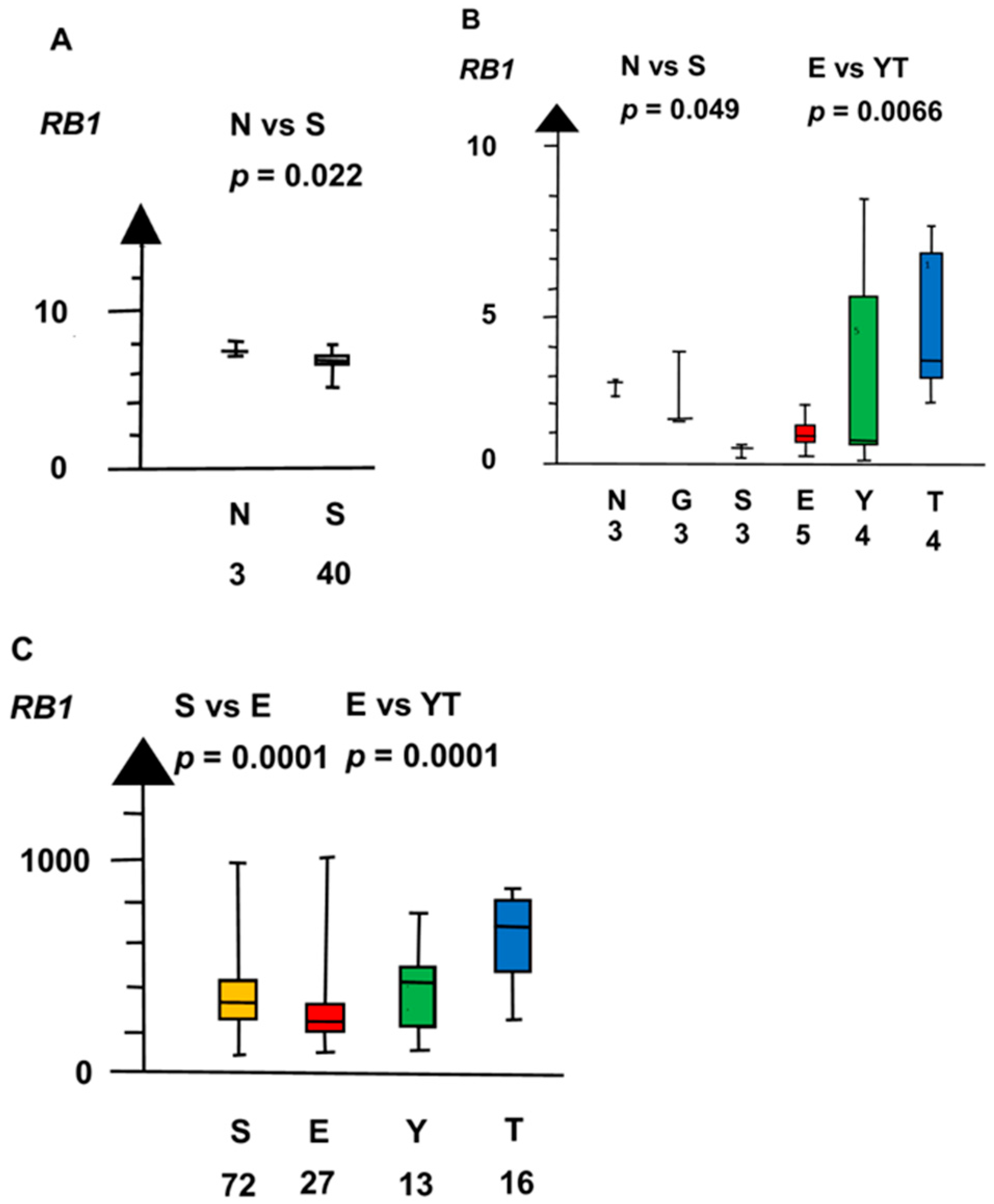
SE and EC combined had an RNA expression of RB1 that was a quarter of that of NT and GCNIS combined (median 0.5 vs. 2.3, p = 0.0495, Kruskal–Wallis test), as shown in Figure 8.
EC had a set of genes with overexpressed RNA that differed from that in SE. EC had high RNA expressions of DMT3B and LIN28A, but the RNA expressions were low in SE.
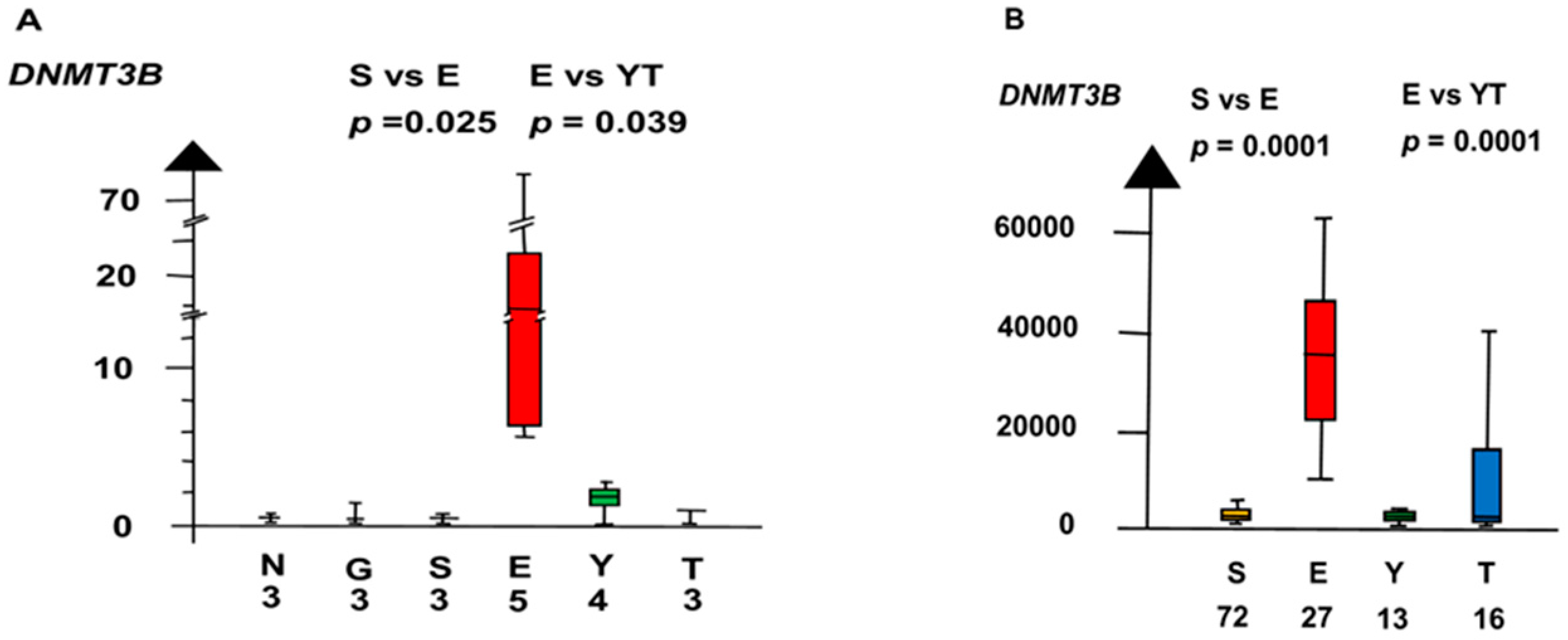
In the second microarray, EC had an 18 times higher RNA expression of DNMT3B than all other histologic types combined (median 18.3 vs. 1, p = 0.0013, Kruskal–Wallis test), as shown in Figure 9.
EC had a three times higher RNA expression of LIN28A than SE (2.6 vs. 0.85, p = 0.18, Kruskal–Wallis test), as shown in Figure 10. LIN28A differed stepwise among the histologic types. SE and EC combined had a six times higher RNA expression of LIN28A than NT and GCNIS combined (median 2.4 vs. 0.36, p = 0.0019, Kruskal–Wallis test).
Differentiated NSGCT, TER, YST, and CC, had genes that were overexpressed, whereas EC did not overexpress the genes. TER had a nine times higher RNA expression of RB1 than EC (median 4.6 vs. 0.5, p = 0.028, Kruskal–Wallis test), as shown in Figure 8. YST had a 21 times higher RNA expression of AFP than EC (median 9.4 vs. 0.4, p = 0.02, Kruskal–Wallis test).
CC had an eleven times higher RNA expression of CGB5 than EC (9.0 vs. median 0.8, p = 0.14, Kruskal–Wallis test).
2.3. Nonsignificant Genes
In the second microarray, the histologic types did not differ significantly in RNA expression of CDKN1A, CDKN2C. CCNE1, FOXD0, MYC, MYCN, MDM2, and PTTG1.
2.4. Role of Genes for the Development of Testicular Germ Cell Tumor Type II (TGCT)
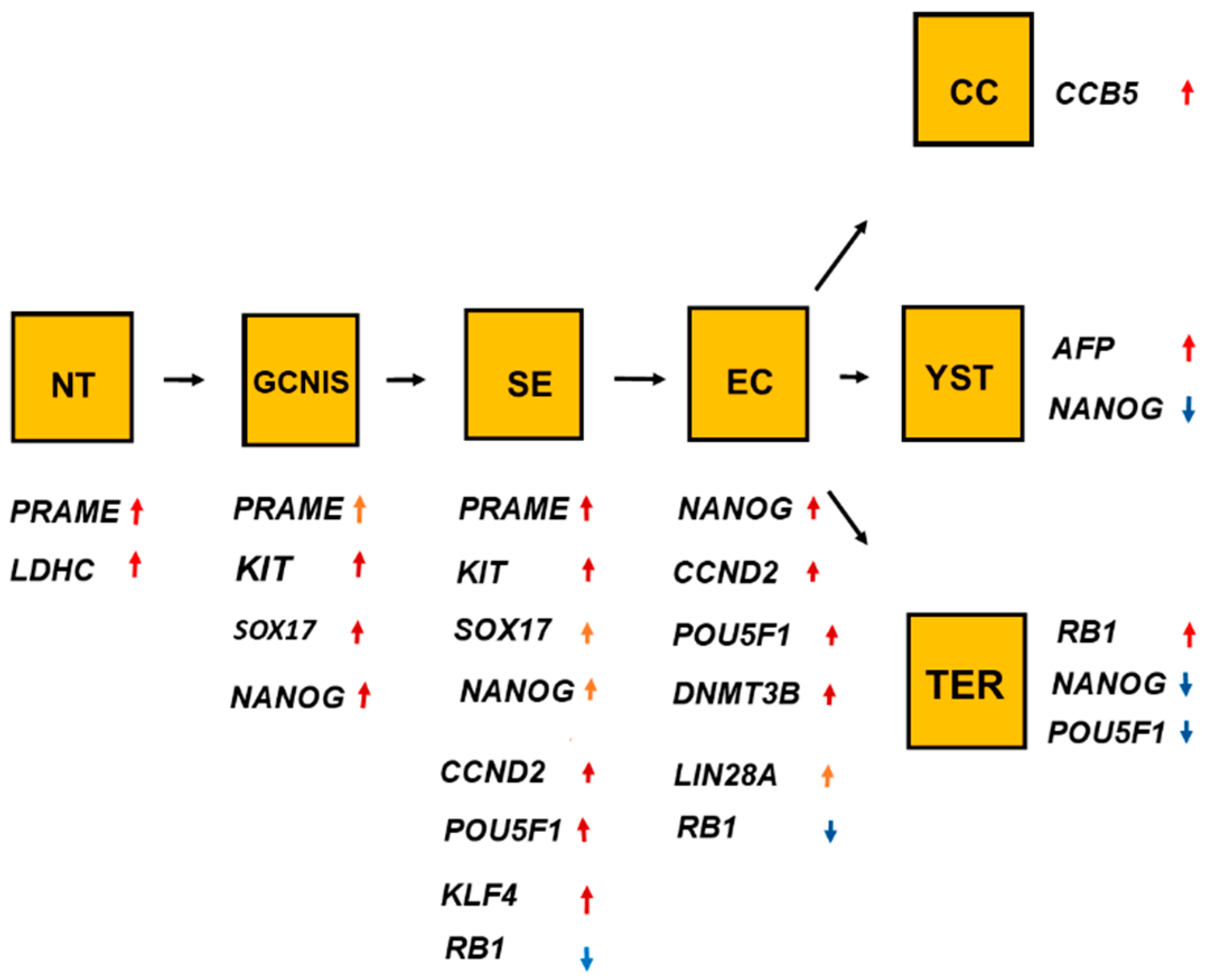
The histologic types of TGCT had considerable overlaps of high RNA expressions of many genes, as shown in Table 1 and Figure 11. The overlaps indicate the histologic types have close relations during the development.
The histologic types also differed in RNA expression of significant genes, as shown in Table 2 and Figure 11. The following two genes were genetic signatures for a histologic type: KLF4 for SE and DNMT3B for EC. The significant differences for RNA expressions of genes among histologic types indicate that the histologic types had different roles for the genes.
In undifferentiated histologic types of TGCT, SE, and EC, high RNA expressions of significant genes are associated with the main clinical features such as self-renewal, hyperproliferation, and sensitivity to platin-based chemotherapy, as indicated in Figure 12.
3. Discussion
Our meta-analysis of cancer genomics of 24 candidate genes of TGCT fulfilled the major goal to identify driver genes for initiation of GCNIS, the precursor lesion, and progression to the histologic types of TGCT. The histologic types differed significantly in RNA expression of PRAME, KIT, SOX17, NANOG, CCND2, KLF4, POU5F1, RB1, DNMT3B, and LIN28A.
Eight of the ten significant candidate genes differed concordantly in the three datasets regarding RNA expressions among the histologic types. Thus, our gene analyses indisputably support that NT develops over GCNIS to SE, EC, and the differentiated types of NSGCT.
Our meta-analysis evaluated the individual tissue samples in the three datasets for the RNA expressions. Combined in a meta-analysis of the p values of the three datasets by the Fisher’s test [15], the summary p findings were highly significant.
Undifferentiated types of TGCT had a high RNA expression of CCND2 and NANOG and both genes have their gene locus on chromosome 12p (12p13.32 and 12p13.31, respectively). Thus, the RNA expressions of the two genes in part explains how the high copy numbers of chromosome 12p in TGCT contribute to the pathogenesis of TGCT.
Although the recent WHO classification 2016 gave the precursor lesion an ambiguous name, GCNIS, the precursor lesion in reality is a localized malignancy. Half the GCNIS progress to microinvasive germ cell tumors within five years [16]. GCNIS was associated with high RNA expression of three genes, and SE was associated with high expression of two other genes and a reduced expression of a tumor suppressor gene.
SE can progress to EC. Both SE and EC, had high RNA expression of CCND2, NANOG, and POU5F1 and, furthermore, both SE and EC had reduced RNA expression of RB1. The stepwise increase among GCNIS and the undifferentiated types of TGCT in levels of RNA expression of the three genes supports that the genes are driver genes in the pathogenesis of TGCT.
SE and EC also differed. SE had high RNA expressions of PRAME, KIT, and SOX17, not found in EC. EC had a high RNA expression of LIN28A not found in SE. SE had KLF4 as a genetic signatures and EC had DMNT3B as a genetic signature. However, the differences between SE and EC do not prove that SE and EC develop according to separate lines.
The genes have two roles in the pathogenesis of TGCT. Overlap of high RNA expression of many genes in the histologic types bind the histologic types together. Differences in genes among the histologic types can be primary or secondary. Two genes had a high RNA expression only in one histologic type as a genetic signature. The two genes contribute to the specific characteristics of SE and EC.
For SE, KLF4 is a genetic signature. KLF4 could block SE for differentiation into differentiated types of NSGCT, TER, YST, and CC. RNA expression of KLF4 had a longer half-life when the cells expressed NANOG and POU5F1 [17].
DNMT3B methylates DNA. For EC, DNMT3B is a genetic signature. The high RNA expression of DNMT3B causes EC to have more methylation of DNA than SE. Similarly, a proteomic study found that EC had high protein expression of DNMT3B [18]. Methylation of an upstream region for POU5F1 regulated expression of POU5F1 [18]. Downregulation of DNMT3B is part of the difference in gene expressions as EC differentiates into TER, YST, and CC.
Our results are in accordance with previous research. Undifferentiated and differentiated histologic types of TGCT differed in RNA expression of genes for pluripotency and, similarly, the two histologic types differed in protein levels of the genes in previous publications. Thus, TGCT mainly regulates the proteins at the step of transcription of the genes, like eukaryotic cells, in general, regulate the protein levels.
Our findings are consistent with an experimental study that xenografted a seminoma cell line, TCam2, to the flanks of nude mice, and therefore the TCam2 thereby developed into EC [19,20,21]. During the transition of TCam2 into EC, the bromodomain family of genes upregulated DNMT3B and LIN28A [20,21]. Both SE and EC had high RNA expression of NANOG and POU5F1.
The mechanisms caused EC, in our meta-analysis, to have similarly higher RNA expression of DNMT3B, LIN28A, NANOG, and POU5F1 than SE, similar to the mechanisms reported in the experimental study.
In addition, a previous systematic review [7] showed that TGCT upregulated CCND2 and downregulated RB1. Our findings were also consistent with the observations that GCNIS overexpressed genes for pluripotency [22], that MYC did not have a significant role in the development of the histologic types of TGCT [23], and that EC and the differentiated types of NSGCT differed in the level of the proteins for pluripotency [24].
Further previous studies also showed that SE and NSGCT had higher expression of NANOG and POU5F1 than GCNIS, YST, and TER [25,26]. The differences between EC and differentiated NSGCT in our meta-analysis for NANOG, POU5F1, and SOX17 are like the findings in another study as EC differentiates into TER. Methylations of CpG dinucleotides in NANOG regulatory regions downregulated gene expression of NANOG [27].
Importantly, our meta-analysis showed that genes for pluripotency were upregulated in SE and EC and not only in GCNIS. The genes had a stepwise increase in RNA expression from NT over GCNIS, and SE to EC and a reduction in TER. The changes indicate that epigenetics regulates the RNA expression of the genes.
Epigenetics for GCT can involve that methylations and acetylations of specific lysines on histone 3 (H3K2, H3K9, and H3K27) in promoters and enhancers of the significant candidate genes regulate the RNA expression of the genes [28,29,30].
NANOG can act together with SOX2 and POU5F1 and the complex of transcription factors can bind to a motif in DNA [31]. The complex of NANOG, POU5F1, and SOX2 especially increased RNA expression of NANOG in cells with a bivalent promoter for NANOG.
A panel of four genes for pluripotency including KLF4, LIN28A, MYC, NANOG, and POU5F1 can reprogram somatic cells into induced pluripotent stem cells, iPSC [32,33]. The panel of genes for pluripotency is crucial in developmental biology. The genes for pluripotency can also be driver genes in the pathogenesis of TGCT type II in general.
As proof of the impact in patients with TGCT of genes for pluripotency, a set of genes for pluripotency could reprogram fibroblasts into iPSC in a report of a seminoma patient [34].
Both EC and embryonal stem cells (ESC) have a high expression of genes for pluripotency [35], but iPSC has similarly high RNA expression of the genes. Both EC and iPSC, but not ESC, can differentiate into TER. A study showed that xenograft with iPSC could develop into TER or into EC depending on the expression level of genes for pluripotency [36]. Therefore, the data favors the importance of the biologic similarity between EC and iPSC.
Knockdown studies support that our candidate genes are important. Knockdown of PRAME showed that PRAME inhibited that SE differentiated into TER [37]. Knockdown of LIN28A showed that LIN28A stimulates expression of LIN28A, NANOG, and POU5F1 [38].
Nettersheim argued that TGCT developed based on a bidirectional plasticity [9,10]. The plasticity implies that SE develops into EC, as well as EC develops into SE. However, GCNIS progresses into TGCT [2] and EC progresses into the differentiated histologic types of NSGCT as indicated by Dixon and Moore [4]. Therefore, the histologic types of TGCT are linked by a unidirectional progression.
The Genomic Cancer Atlas (TGCA) extensively reported RNA findings in relation to the development of TGCT [12]. TGCA supported that the histologic types of TGCT develop in two lines for the development. However, our analysis of 24 candidate genes in the TGCA RNAseq dataset supports that TCGT develops in one-line progression among the histologic types.
The Vogelstein group indicates that a total of three mutations of oncogenes and tumor suppressor genes are sufficient to drive the malignant transformation [39], and that epigenetic regulations can substitute for the mutations in the malignant transformation. The iPSC shows that a panel of four genes of pluripotency can transform somatic cells into iPSC [32,33]. Therefore, four genes for pluripotency can reprogram NT into EC at least in hypotetraploid germ cells.
In TGCT, NSGCT often includes a combination of the histologic types [40]. However, a multivariate analysis showed that the various combinations of histologic types had no prognostic implications.
Future studies could further investigate how epigenetics regulates our candidate genes. It would be interesting to analyze whether RNA expression of the candidate genes among the histologic types of TGCT are associated with methylations and acetylations of lysines on histone 3 of promoters and enhancers for the candidate genes.
Our meta-analysis has limitations. It is not a systematic review. Pathologists accept that genes for pluripotency are important in elder patients for the development of GCT [41,42,43,44]. But most pathologists are not convinced that young adult patients, in general, develop TGCT due to a combined impact from a panel of genes for pluripotency.
In conclusion, PRAME, KIT, SOX17, CCND2, KLF4, NANOG, POU5F1, DNMT3B, RB1, and LIN28A are important in the pathogenesis of TGCT.
4. Materials and Methods
4.1. Data Sources
The meta-analysis evaluated two microarray datasets and an RNA sequencing (seq) dataset. The first microarray [13] used Affymetrix HG-U95A v2 (Thermo Fisher Scientific, Waltman, MA, USA) to analyze RNA expression. The microarray was available in Gene Expression Omnibus (GEO) at ascension number GSE 8607 (http:www.ncbi.gov.gen/query/ace.cgi?acc.GSE 8607). The first microarray included three samples of NT and 40 samples of SE.
The second microarray [14] used Agilent Human 1 A oligo assay (Agilent Technologies Inc, Palo Alto, CA, USA) to analyze RNA expression and was available in GEO at ascension number GSE 1818 (http:ww.ncbi.gov.gen/query/ace.cgi?acc.GSE 1818) and included three samples of NT, three samples of GCNIS, three samples of SE, five samples of EC, four samples of YST, four samples of YST, and one sample of CC.
The microarray summarized gene expression on log2 scale relative to a reference standard.
The TCGA Research Network made the RNAseq database [12] available at (http://portal.gdc.cancer.gov/legacy-archieve/).
The RNAseq dataset included 72 samples of SE, 27 samples of EC, 13 samples of YST, and 16 samples of TER.
Regarding the second microarray, we carried out inverse logarithmic transformation before we analyzed and reported the RNA expressions.
The original studies of the three datasets followed the Declaration of Helsinki.
4.2. Candidate Genes
The meta-analysis selected genes that previous publications had indicated were significant for the development of TGCT. Overall, our meta-analysis included six genes for pluripotency, five proto-oncogenes, four tumor suppressor genes, five genes for the three serum tumor markers, and four genes for biomarkers and a testis cancer antigen.
The six genes for pluripotency, KLF4, LIN28, MYC, NANOG, POU5F1, and SOX17 were among the genes with the highest gene expression in GCNIS [22]. Of these genes, SE expresses SOX17 but not SOX2, whereas EC expresses SOX2 and not SOX17. Unfortunately, the two microarrays did not include SOX2, therefore, we were unable to undertake a meta-analysis of SOX2.
TGCT can include upregulated proto-oncogenes such as CCND2, MDM2, MYC, NMYC, PTTG1, and CCNE1 [45,46,47,48]. TGCT can include downregulated tumor suppressor genes such as CDK1N1A, CDKN2C, PTEN, and RB1 [49,50,51,52,53,54,55,56,57]. DNMT3B as a biomarker of TGCT.
Our list of candidate genes included AFP, CGB5, LDHA, LDHB, and LDHC for the genes for the serum tumor markers, alpha fetoprotein (AFP), human chorionic gonadotropin (hCG), and lactate dehydrogenase (EC 1.1.1.27, LDH), because the tumor markers are important for the treatment of TGCT. AFP and CGB5 were secondary to the histologic types, and therefore the present meta-analysis included only limited analysis of the tumor marker genes.
4.3. Definitions and Evaluations
The meta-analysis defined KLF4, LIN28A, NANOG, and POU5F1 as genes for pluripotency. We took a low combined p value from more than one dataset to indicate a role of the gene for the development of TGCT. We defined a gene as a driver gene if RNA expression of the gene increased stepwise from NT to SE/EC. We defined other significant genes as passenger genes.
We defined RNA expression of a gene as insignificant if the RNA expression of the gene did not differ statistically significantly among NT and the histologic types of TGCT.
A high RNA expression in two or more histologic types indicated an overlap of gene expression as a link as a histologic type progresses to the next. A significant change in RNA expression among histologic types indicated that the gene contributed to the development among the histologic types. We defined a significantly high RNA expression in only one histologic type as a genetic signature.
4.4. Statistical Analyses
The meta-analysis gave priority to comparisons among gene expressions in histologic types of TGCT that could be based on more than one dataset. We showed the combined p values based on the p values for comparisons of RNA expression of histologic types of the individual datasets using the meta-analytic method of the Fisher’s test [15].
The meta-analysis did not substitute missing data in the datasets of RNA expression in a tissue sample. The RNA expressions of the candidate genes had non-parametric distributions. Therefore, the meta-analysis used the median values for the RNA expressions of the genes and the Kruskal–Wallis test as we compared two groups of histologic types. All statistical tests were two sided. We considered a p value < 0.05 as statistically significant.
We carried out the statistical analyses with Stata 14.2 (Stata Corp., TX, USA).
Author Contributions
F.E.v.E. developed the concept for the study and wrote the first draft of the manuscript; J.P.-A. published an analysis of the first microarray study, searched for other microarray datasets, and was active in the writing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
No external funding.
Acknowledgments
We thank Rolf Inge Skotheim, the Department of molecular oncology, Institute of Cancer Research, Oslo University, Norway, for help regarding the second microarray dataset.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hermanek, P.; International Union Against Cancer. The TNM Atlas: Illustrated Guide to the TNM/pTNM Classification of Malignant Tumours, 4th ed.; Springer: Berlin, NY, USA, 1987. [Google Scholar]
- Looijenga, L.H.J.; Kao, C.S.; Idrees, M.T. Predicting Gonadal Germ Cell Cancer in People with Disorders of Sex Development; Insights from Developmental Biology. Int. J. Mol. Sci. 2019, 20, 5017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilms, M. The teratoid testicular tumors including so called cystoid and enchondroma, In German. Beitr. Der Pathol. Anat. 1896, 36, 266–337. [Google Scholar]
- Dixon, F.J.; Moore, R.A. Testicular tumors: A clinicopathological study. Cancer 1953, 6, 427–454. [Google Scholar] [CrossRef]
- Teilum, G. Endodermal sinus tumors of the ovary and testis. Comparative morphogenesis of the so-called mesoephroma ovarii (Schiller) and extraembryonic (yolk sac-allantoic) structures of the rat’s placenta. Cancer 1959, 12, 1092–1105. [Google Scholar] [CrossRef]
- De Jong, B.; Oosterhuis, J.W.; Castedo, S.M.; Vos, A.; te Meerman, G.J. Pathogenesis of adult testicular germ cell tumors: A cytogenetic model. Cancer Genet. Cytogenet. 1990, 48, 143–167. [Google Scholar] [CrossRef]
- Von Eyben, F.E. Chromosomes, Genes, and Development of Testicular Germ Cell Tumors. Cancer Genet. Cytogenet. 2004, 151, 93–138. [Google Scholar] [CrossRef]
- Alagaratnam, S.; Lind, G.E.; Kraggerud, S.M.; Lothe, R.A.; Skotheim, R.I. The testicular germ cell tumour transcriptome. Int. J. Androl. 2011, 34, e133–e150, discussion e50–e51. [Google Scholar] [CrossRef]
- Nettersheim, D.; Westernstroer, B.; Haas, N.; Leinhaas, A.; Brustle, O.; Schlatt, S.; Schorle, H. Establishmlent of a versatile seminoma model indicates cellular plasticity of germ cell tumor cells. Genes Chromosomes Cancer 2012, 51, 717–726. [Google Scholar] [CrossRef]
- Nettersheim, D.; Jostes, S.; Schneider, S.; Schorle, H. Elucidating human male germ cell development by studying germ cell cancer. Reproduction 2016, 152, R101–R113. [Google Scholar] [CrossRef] [Green Version]
- von Eyben, F.E.; Jensen, M.B.; Hoyer, S. Frequency and Markers of Precursor Lesions and Implications for the Pathogenesis of Testicular Germ Cell Tumors. Clin. Genitourin. Cancer. 2018, 16, e211–e221. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Shih, J.; Hollern, D.P.; Wang, L.; Bowlby, R.; Tickoo, S.K.; Thorsson, V.; Mungall, A.J.; Newton, Y.; Hedge, A.M.; et al. Integrated Molecular Characterization of Testicular Germ Cell Tumors. Cell Rep. 2018, 23, 3392–3406. [Google Scholar] [CrossRef] [PubMed]
- Gashaw, I.; Grummer, R.; Klein-Hitpass, L.; Dushaj, O.; Bergmann, M.; Brehm, R.; Grobholz, R.; Kliesch, S.; Neuvians, T.P.; Schmid, K.V.; et al. Gene signatures of testicular seminoma with emphasis on expression of ets variant gene 4. Cell. Mol. Life Sci. 2005, 62, 2359–2368. [Google Scholar] [CrossRef]
- Skotheim, R.I.; Lind, G.E.; Monni, O.; Nesland, J.M.; Abeler, V.M.; Fossa, S.D.; Duale, N.; Brunborg, G.; Kallioniemi, O.; Andrews, P.W.; et al. Differentiation of human embryonal carcinomas in vitro and in vivo reveals expression profiles relevant Duale, to normal development. Cancer Res. 2005, 65, 5588–5598. [Google Scholar] [CrossRef] [Green Version]
- Tobias, V. Meta-analysis of p values. Stata technical Bulletin. Stata Tech. Bull. 1999, 49, 15–17. [Google Scholar]
- Von Eyben, F.E.; Jacobsen, G.K.; Skotheim, R.I. Microinvasive germ cell tumor of the testis. Virchows Arch. 2005, 447, 610–625. [Google Scholar] [CrossRef] [PubMed]
- Dhaliwal, N.K.; Abatti, L.E.; Mitchell, J.A. KLF4 protein stability regulated by interaction with pluripotency transcription factors overrides transcriptional control. Genes Dev. 2019, 33, 1069–1082. [Google Scholar] [CrossRef] [Green Version]
- Chaerkady, R.; Kerr, C.L.; Kandasamy, K.; Marimuthu, A.; Gearhart, J.D.; Pandey, A. Comparative proteomics of human embryonic stem cells and embryonal carcinoma cells. Proteomics 2010, 10, 1359–1373. [Google Scholar] [CrossRef] [Green Version]
- De Jong, J.; Weeda, S.; Gillis, A.J.; Oosterhuis, J.W.; Looijenga, L.H. Differential methylation of the OCT3/4 upstream region in primary human testicular germ cell tumors. Oncol. Rep. 2007, 18, 127–132. [Google Scholar] [CrossRef]
- Nettersheim, D.; Jostes, S.; Sharma, R.; Schneider, S.; Hofmann, A.; Ferreira, H.J.; Hoffman, P.; Kristiansen, G.; Esteller, M.B.; Schorle, H. BMP Inhibition in Seminomas Initiates Acquisition of Pluripotency via NODAL Signaling Resulting in Reprogramming to an Embryonal Carcinoma. PLoS Genet. 2015, 11, e1005415. [Google Scholar] [CrossRef] [Green Version]
- Jostes, S.; Nettersheim, D.; Fellermeyer, M.; Schneider, S.; Hafezi, F.; Honecker, F.; Schumacher, V.; Geyer, M.; Kristiansen, G.; Schorle, H. The bromodomain inhibitor JQ1 triggers growth arrest and apoptosis in testicular germ cell tumours in vitro and in vivo. J. Cell. Mol. Med. 2017, 21, 1300–1314. [Google Scholar] [CrossRef]
- Almstrup, K.; Leffers, H.; Lothe, R.A.; Skakkebaek, N.E.; Sonne, S.B.; Nielsen, J.E.; Rajpert-De Meyts, E.; Skotheim, R.I. Improved gene expression signature of testicular carcinoma in situ. Int. J. Androl. 2007, 30, 292–302, discussion 3. He. [Google Scholar] [CrossRef] [PubMed]
- Buganim, Y.; Markoulaki, S.; van Wietmarschen, N.; Hoke, H.; Wu, T.; Ganz, K.; Akhtar-Zaidi, B.; He, Y.; Abraham, B.J.; Porubsky, D.; et al. The developmental potential of iPSCs is greatly influenced by reprogramming factor selection. Cell Stem Cell 2014, 15, 295–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajpert-De Meyts, E.; Hanstein, R.; Jorgensen, N.; Graem, N.; Vogt, P.H.; Skakkebaek, N.E. Developmental expression of POU5F1 (OCT-3/4) in normal and dysgenetic human gonads. Hum. Reprod. 2004, 19, 1338–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gainetdinov, I.V.; Kondratieva, S.A.; Skvortsova, Y.V.; Zinovyeva, M.V.; Stukacheva, E.A.; Klimov, A.; Tryakin, A.A.; Azhikina, T.L. Distinguishing epigenetic features of preneoplastic testis tissues adjacent to seminomas and nonseminomas. Oncotarget 2016, 7, 22439–22447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korkola, J.E.; Houldsworth, J.; Chadalavada, R.S.; Olshen, A.B.; Dobrzynski, D.; Reuter, V.E.; Bosl, G.J.; Chaganti, R.S. Down-regulation of stem cell genes, including those in a 200-kb gene cluster at 12p13.31, is associated with in vivo differentiation of human male germ cell tumors. Cancer Res. 2006, 66, 820–827. [Google Scholar] [CrossRef] [Green Version]
- Nettersheim, D.; Biermann, K.; Gillis, A.J.; Steger, K.; Looijenga, L.H.; Schorle, H. NANOG promoter methylation and expression correlation during normal and malignant human germ cell development. Epigenetics 2011, 6, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Almstrup, K.; Nielsen, J.E.; Mlynarska, O.; Jansen, M.T.; Jorgensen, A.; Skakkebaek, N.E.; Rajpert De Meyts, E. Carcinoma in situ testis displays permissive chromatin modifications similar to immature foetal germ cells. Br. J. Cancer 2010, 103, 1269–1276. [Google Scholar] [CrossRef]
- Kristensen, D.G.; Skakkebaek, N.E.; Rajpert-De Meyts, E.; Almstrup, K. Epigenetic features of testicular germ cell tumours in relation to epigenetic characteristics of foetal germ cells. Int. J. Dev. Biol. 2013, 57, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Van Der Zwan, Y.G.; Stoop, H.; Rossello, F.; White, S.J.; Looijenga, L.H. Role of epigenetics in the etiology of germ cell cancer. Int. J. Dev. Biol. 2013, 57, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Heurtier, V.; Owens, N.; Gonzalez, I.; Mueller, F.; Proux, C.; Mornico, D.; Clerc, P.; Dubois, A.; Navarro, P. The molecular logic of Nanog-induced self-renewal in mouse embryonic stem cells. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eldar-Geva, T.; Gross-Tsur, V.; Hirsch, H.J.; Altarescu, G.; Segal, R.; Zeligson, S.; Golomb, E.; Epsztejn-Litman, S.; Eiges, R. Incomplete methylation of a germ cell tumor (Seminoma) in a Prader-Willi male. Mol. Genet. Genomic Med. 2018, 6, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Almstrup, K.; Hoei-Hansen, C.E.; Wirkner, U.; Blake, J.; Schwager, C.; Ansorge, W.; Nielsen, J.E.; Skakkebaek, N.E.; Rajpert De Meyts, E. Embryonic stem cell-like features of testicular carcinoma in situ revealed by genome-wide gene expression profiling. Cancer Res. 2004, 64, 4736–4743. [Google Scholar] [CrossRef] [Green Version]
- Salvatori, D.C.F.; Dorssers, L.C.J.; Gillis, A.J.M.; Perretta, G.; van Agthoven, T.; Gomes Fernandes, M.; Stoop, H.; Prins, J.B.; Oosterhuis, J.W.; Mummery, C.; et al. The MicroRNA-371 Family as Plasma Biomarkers for Monitoring Undifferentiated and Potentially Malignant Human Pluripotent Stem Cells in Teratoma Assays. Stem. Cell Rep. 2018, 11, 1493–1505. [Google Scholar] [CrossRef] [Green Version]
- Nettersheim, D.; Arndt, I.; Sharma, R.; Riesenberg, S.; Jostes, S.; Schneider, S.; Holzel, M.; Kristiansen, G.; Schorle, H. The cancer/testis antigen PRAME supports the pluripotency network and expresses somatic and germ cell differentiation programs in seminomas. Br. J. Cancer 2016, 115, 454–464. [Google Scholar] [CrossRef] [Green Version]
- Gillis, A.J.; Stoop, H.; Biermann, K.; van Gurp, R.J.; Swartzman, E.; Cribbes, S.; Ferlinz, A.; Shannon, M.; Oosterhuis, J.W.; Looijenga, L.H. Expression 8and interdependencies of pluripotency factors LIN28, OCT3/4, NANOG and SOX2 in human testicular germ cells and tumours of the testis. Int. J. Androl. 2011, 34, e160–e174. [Google Scholar] [CrossRef]
- Tomasetti, C.; Marchionni, L.; Nowak, M.A.; Parmigiani, G.; Vogelstein, B. Only three driver gene mutations are required for the development of lung and colorectal cancers. Proc. Natl. Acad. Sci. USA 2015, 112, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Eyben, F.E.; Jacobsen, G.K.; Pedersen, H.; Jacobsen, M.; Clausen, P.P.; Zibrandtsen, P.C.; Gullberg, B. Multivariate analysis of risk factors in patients with metastatic testicular germ cell tumors treated with vinblastine and bleomycin. Invasion Metastasis 1982, 2, 125–135. [Google Scholar]
- Nogales, F.F.; Prat, J.; Schuldt, M.; Cruz-Viruel, N.; Kaur, B.; D’Angelo, E.; Matias-Guiu, X.; Vidal, A.; McCluggage, W.G.; Oosterhuis, J.W. Germ cell tumour growth patterns originating from clear cell carcinomas of the ovary and endometrium: A comparative immunohistochemical study favouring their origin from somatic stem cells. Histopathology 2018, 72, 634–647. [Google Scholar] [CrossRef]
- Lobo, J.; Gillis, A.J.M.; Jeronimo, C.; Henrique, R.; Looijenga, L.H.J. Human Germ Cell Tumors are Developmental Cancers: Impact of Epigenetics on Pathobiology and Clinic. Int. J. Mol. Sci. 2019, 20, 258. [Google Scholar] [CrossRef] [Green Version]
- Wermann, H.; Stoop, H.; Gillis, A.J.; Honecker, F.; van Gurp, R.J.; Ammerpohl, O.; Richter, J.; Oosterhuis, J.W.; Bokemeyer, C.; Looijenga, L.H. Global DNA methylation in fetal human germ cells and germ cell tumours: Association with differentiation and cisplatin resistance. J. Pathol. 2010, 221, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Oosterhuis, J.W.; Looijenga, L.H.J. Human germ cell tumours from a developmental perspective. Nat. Rev. Cancer 2019, 19, 522–537. [Google Scholar] [CrossRef] [PubMed]
- Sikora, K.; Evan, G.; Stewart, J.; Watson, J.V. Detection of the c-myc oncogene product in testicular cancer. Br. J. Cancer 1985, 52, 171–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godmann, M.; Gashaw, I.; Eildermann, K.; Schweyer, S.; Bergmann, M.; Skotheim, R.I.; Behr, R. The pluripotency transcription factor Kruppel-like factor 4 is strongly expressed in intratubular germ cell neoplasia unclassified and seminoma. Mol. Hum. Reprod. 2009, 15, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Jones, T.D.; Ulbright, T.M.; Eble, J.N.; Cheng, L. OCT4: A sensitive and specific biomarker for intratubular germ cell neoplasia of the testis. Clin. Cancer Res. 2004, 10, 8544–8547. [Google Scholar] [CrossRef] [Green Version]
- Datta, M.W.; Macri, E.; Signoretti, S.; Renshaw, A.A.; Loda, M. Transition from in situ to invasive testicular germ cell neoplasia is associated with the loss of p21 and gain of mdm-2 expression. Mod. Pathol. 2001, 14, 437–442. [Google Scholar] [CrossRef]
- Bartkova, J.; Rajpert-de Meyts, E.; Skakkebaek, N.E.; Bartek, J. D-type cyclins in adult human testis and testicular cancer: Relation to cell type, proliferation, differentiation, and malignancy. J. Pathol. 1999, 187, 573–581. [Google Scholar] [CrossRef]
- Pierconti, F.; Milardi, D.; Martini, M.; Grande, G.; Cenci, T.; Gulino, G.; Larocca, L.M.; Rindi, G.; Pontecorvi, A.; De Marinis, L. Pituitary-tumour-transforming-gene 1 expression in testicular cancer. Andrologia 2015, 47, 427–432. [Google Scholar] [CrossRef]
- Schmidt, B.A.; Rose, A.; Steinhoff, C.; Strohmeyer, T.; Hartmann, M.; Ackermann, R. Up-regulation of cyclin-Dependent kinase 4/cyclin D2 expression but down-regulation of cyclin-dependent kinase 2/cyclin E in testicular germ cell tumors. Cancer Res. 2001, 61, 4214–4221. [Google Scholar]
- Grande, G.; Milardi, D.; Martini, M.; Cenci, T.; Gulino, G.; Mancini, F.; Bianchi, A.; Pontecorvi, A.; Pierconti, F. Protein Expression of PTTG-1, OCT-4, and KLF-4 in Seminoma: A Pilot Study. Front. Endocrinol. (Lausanne) 2019, 10, 619. [Google Scholar] [CrossRef]
- Bartkova, J.; Lukas, C.; Sorensen, C.S.; Rajpert-De Meyts, E.; Skakkebaek, N.E.; Lukas, J.; Bartek, J. Deregulation of the RB pathway in human testicular germ cell tumours. J. Pathol. 2003, 200, 149–156. [Google Scholar] [CrossRef]
- Bartkova, J.; Thullberg, M.; Rajpert-De Meyts, E.; Skakkebaek, N.E.; Bartek, J. Cell cycle regulators in testicular cancer: Loss of p18INK4C marks progression from carcinoma in situ to invasive germ cell tumours. Int. J. Cancer 2000, 85, 370–375. [Google Scholar] [CrossRef]
- Di Vizio, D.; Cito, L.; Boccia, A.; Chieffi, P.; Insabato, L.; Pettinato, G.; Motti, M.L.; Shepis, F.; D’Amico, W.; Fabiani, F.; et al. Loss of the tumor suppressor gene PTEN marks the transition from intratubular germ cell neoplasias (ITGCN) to invasive germ cell tumors. Oncogene 2005, 24, 1882–1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartkova, J.; Thullberg, M.; Rajpert-De Meyts, E.; Skakkebaek, N.E.; Bartek, J. Lack of p19INK4d in human testicular germ-cell tumours contrasts with high expression during normal spermatogenesis. Oncogene 2000, 19, 4146–4150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strohmeyer, T.; Reissmann, P.; Cordon-Cardo, C.; Hartmann, M.; Ackermann, R.; Slamon, D. Correlation between retinoblastoma gene expression and differentiation in human testicular tumors. Proc. Natl. Acad. Sci. USA 1991, 88, 6662–6666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
RNA expression of PRAME in the two microarrays (A,B) show that normal testis (N), germ cell neoplasia in situ (G) and seminoma (S) had high RNA expression and embryonal carcinoma (E), yolk sac tumor (Y), and teratoma (T) had a low RNA expression. The boxes show the median and the upper and lower quartiles and the whiskers show the full range.
Figure 1.
RNA expression of PRAME in the two microarrays (A,B) show that normal testis (N), germ cell neoplasia in situ (G) and seminoma (S) had high RNA expression and embryonal carcinoma (E), yolk sac tumor (Y), and teratoma (T) had a low RNA expression. The boxes show the median and the upper and lower quartiles and the whiskers show the full range.

Figure 2.
RNA expression of KIT in the three datasets (A–C) show that germ cell neoplasia in situ (G) and seminoma (S) had high RNA expression and that embryonal carcinoma (E), yolk sac tumor (Y), and teratoma (T) had a low RNA expression.
Figure 2.
RNA expression of KIT in the three datasets (A–C) show that germ cell neoplasia in situ (G) and seminoma (S) had high RNA expression and that embryonal carcinoma (E), yolk sac tumor (Y), and teratoma (T) had a low RNA expression.

Figure 3.
RNA expression of SOX17 in the two microarrays (A,B) show that germ cell neoplasia in situ (G), seminoma (S), and yolk sac tumor (Y) had a high RNA expression and that embryonal carcinoma (E) and teratoma (T) had a low RNA expression.
Figure 3.
RNA expression of SOX17 in the two microarrays (A,B) show that germ cell neoplasia in situ (G), seminoma (S), and yolk sac tumor (Y) had a high RNA expression and that embryonal carcinoma (E) and teratoma (T) had a low RNA expression.

Figure 4.
RNA expression of NANOG in the second microarray and the RNAseq dataset (A,B) show that germ cell neoplasia in situ (G), seminoma (S), and embryonal carcinoma (E) had a high RNA expression and that yolk sac tumor (Y) and teratoma (T) had a low RNA expression.
Figure 4.
RNA expression of NANOG in the second microarray and the RNAseq dataset (A,B) show that germ cell neoplasia in situ (G), seminoma (S), and embryonal carcinoma (E) had a high RNA expression and that yolk sac tumor (Y) and teratoma (T) had a low RNA expression.

Figure 5.
RNA expression of CCND2 in the three datasets (A–C) show that germ cell neoplasia in situ (G), seminoma (S), and embryonal carcinoma (E) had a high RNA expression and that yolk sac tumor (Y) and teratoma (T) had a low RNA expression.
Figure 5.
RNA expression of CCND2 in the three datasets (A–C) show that germ cell neoplasia in situ (G), seminoma (S), and embryonal carcinoma (E) had a high RNA expression and that yolk sac tumor (Y) and teratoma (T) had a low RNA expression.

Figure 6.
RNA expression of POU5F1 in three datasets (A–C) show that seminoma (S) and embryonal carcinoma (E) had a high RNA expression and that germ cell neoplasia in situ (G), yolk sac tumor (Y), and teratoma (T) had a low RNA expression.
Figure 6.
RNA expression of POU5F1 in three datasets (A–C) show that seminoma (S) and embryonal carcinoma (E) had a high RNA expression and that germ cell neoplasia in situ (G), yolk sac tumor (Y), and teratoma (T) had a low RNA expression.

Figure 7.
RNA expression of KLF4 in the three datasets (A–C) show that seminoma (S) had a high RNA expression and that germ cell neoplasia in situ (G), embryonal carcinoma (E), yolk sac tumor (Y), and teratoma (T) had a low RNA expression.
Figure 7.
RNA expression of KLF4 in the three datasets (A–C) show that seminoma (S) had a high RNA expression and that germ cell neoplasia in situ (G), embryonal carcinoma (E), yolk sac tumor (Y), and teratoma (T) had a low RNA expression.

Figure 8.
RNA expression of RB1 in the three datasets (A–C) show that seminoma (S) and embryonal carcinoma (E) had a low RNA expression and that teratoma (T) had a high RNA expression.
Figure 8.
RNA expression of RB1 in the three datasets (A–C) show that seminoma (S) and embryonal carcinoma (E) had a low RNA expression and that teratoma (T) had a high RNA expression.

Figure 9.
RNA expression of DNMT3B. in the second microarray and the RNAseq dataset (A,B) show that embryonal carcinoma (E) had a high RNA expression and that germ cell neoplasia in situ (G), seminoma (S), yolk sac tumor (Y), and teratoma (T) had a low RNA expression.
Figure 9.
RNA expression of DNMT3B. in the second microarray and the RNAseq dataset (A,B) show that embryonal carcinoma (E) had a high RNA expression and that germ cell neoplasia in situ (G), seminoma (S), yolk sac tumor (Y), and teratoma (T) had a low RNA expression.

Figure 10.
RNA expression of LIN28A in the second microarray and the RNAseq dataset (A,B) show that embryonal carcinoma (E) had a high RNA expression and that yolk sac tumor (Y), and teratoma (T) had a low RNA expression.
Figure 10.
RNA expression of LIN28A in the second microarray and the RNAseq dataset (A,B) show that embryonal carcinoma (E) had a high RNA expression and that yolk sac tumor (Y), and teratoma (T) had a low RNA expression.

Figure 11.
The overlaps and differences of the ten genes shown in Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9 and Figure 10 showed that one gene was upregulated in normal testis (NT), and other genes were upregulated in germ cell neoplasia in situ (GCNIS). Undifferentiated histologic types, seminoma (SE) and embryonal carcinoma (EC), differed in RNA expression. EC differed in RNA expression from the differentiated histologic types such as choriocarcinoma (CC), yolk sac tumor (YST), and teratoma (TER). The figure indicates the linkage among the histologic types. The change from GNIS to SE was associated with a change in RNA expression of three significant genes. As for the genes, red arrows denote increased RNA expression and the blue arrows denote reduced RNA expression.
Figure 11.
The overlaps and differences of the ten genes shown in Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8, Figure 9 and Figure 10 showed that one gene was upregulated in normal testis (NT), and other genes were upregulated in germ cell neoplasia in situ (GCNIS). Undifferentiated histologic types, seminoma (SE) and embryonal carcinoma (EC), differed in RNA expression. EC differed in RNA expression from the differentiated histologic types such as choriocarcinoma (CC), yolk sac tumor (YST), and teratoma (TER). The figure indicates the linkage among the histologic types. The change from GNIS to SE was associated with a change in RNA expression of three significant genes. As for the genes, red arrows denote increased RNA expression and the blue arrows denote reduced RNA expression.

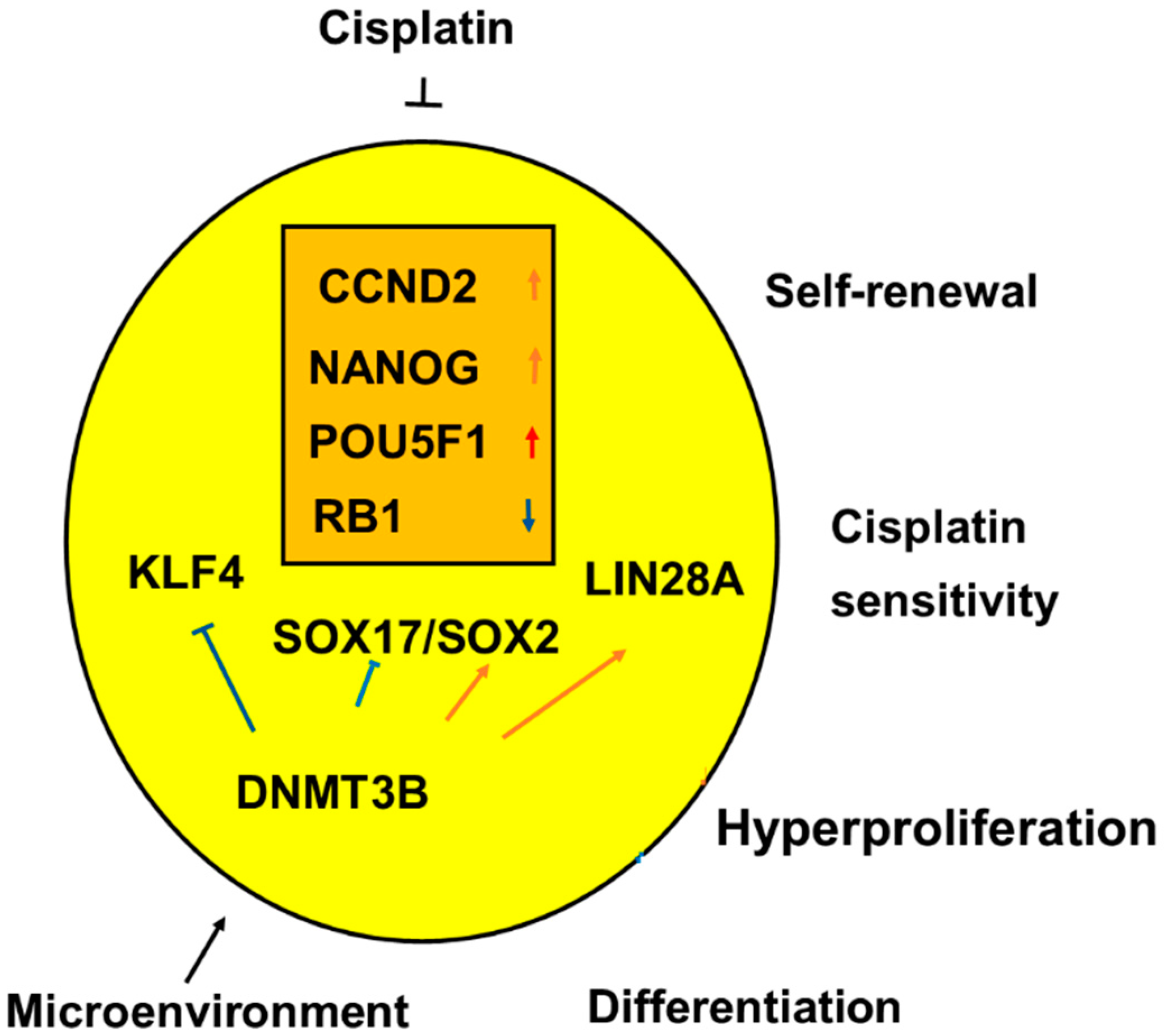
Figure 12.
The undifferentiated histologic types of testicular germ cell tumor type II (TGCT), seminoma (SE), and embryonal carcinoma (EC) had overlap with upregulation of three genes shown with red arrows and downregulation of one gene shown with a blue arrow in the orange box. Microenvironmental factors can upregulate DNMT3B that blocks SOX17 and KLF4 and upregulates SOX2 and LIN28A, genetic changes associated with the histologic transition from a seminomatous cell type to an embryonal carcinoma cell type. Significant genes are associated with the main features of the malignant undifferentiated germ cells such as self-renewal, hyperproliferation, and sensitivity to platin-based chemotherapy. As for the genes, red arrows denote increased RNA expressions or a stimulus of genes and the blue arrows denote reduced RNA expression or an inhibition of genes.
Figure 12.
The undifferentiated histologic types of testicular germ cell tumor type II (TGCT), seminoma (SE), and embryonal carcinoma (EC) had overlap with upregulation of three genes shown with red arrows and downregulation of one gene shown with a blue arrow in the orange box. Microenvironmental factors can upregulate DNMT3B that blocks SOX17 and KLF4 and upregulates SOX2 and LIN28A, genetic changes associated with the histologic transition from a seminomatous cell type to an embryonal carcinoma cell type. Significant genes are associated with the main features of the malignant undifferentiated germ cells such as self-renewal, hyperproliferation, and sensitivity to platin-based chemotherapy. As for the genes, red arrows denote increased RNA expressions or a stimulus of genes and the blue arrows denote reduced RNA expression or an inhibition of genes.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
RNA expressions of genes with overlaps among histologic types.
| Histologies | Genes | No Patients | Combined p Values |
|---|---|---|---|
| SvEYT | PRAME | 6 × 10−5 | |
| KLF4 | 1.4 × 10−5 | ||
| EvYT | CCND2 | 160 | 8.7 ×10−5 |
| POU5F1 | 160 | 1 × 10−5 | |
| RB1 | 160 | 1 × 10−5 | |
| DNMT3B | 160 | 5.2 × 10−5 |
Abbreviations: SvEYT, SE versus EC, YST, and TER; EvYT, EC versus YST and TER.
Table 2.
RNA expressions of genes that separate the histologic types.
| Histologies | Genes | No. of Patients | Combined p Values |
|---|---|---|---|
| NvS | KLF4 | 60 | 0.042 |
| KIT | 60 | 0.0022 | |
| RB1 | 60 | 0.008 | |
| POUF1 | 60 | 0.0019 | |
| CCND2 | 60 | 0.0011 | |
| SvE | SOX17 | 160 | 3.4 × 10−5 |
| KIT | 160 | 3.4 ×10−5 | |
| POU5F1 | 160 | 0.0019 | |
| LIN28A | 160 | 2.1 × 10−4 | |
| DNMT3B | 160 | 3.5 × 10−5 |
Abbreviations: NvS, NT versus SE; SvE, SE versus EC.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
von Eyben, F.E.; Parraga-Alava, J. Meta-Analysis of Gene Expressions in Testicular Germ Cell Tumor Histologies. Int. J. Mol. Sci. 2020, 21, 4487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124487
AMA Style
von Eyben FE, Parraga-Alava J. Meta-Analysis of Gene Expressions in Testicular Germ Cell Tumor Histologies. International Journal of Molecular Sciences. 2020; 21(12):4487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124487
Chicago/Turabian Stylevon Eyben, Finn Edler, and Jorge Parraga-Alava. 2020. "Meta-Analysis of Gene Expressions in Testicular Germ Cell Tumor Histologies" International Journal of Molecular Sciences 21, no. 12: 4487. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124487
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.