Self-Organized Liver Microtissue on a Bio-Functional Surface: The Role of Human Adipose-Derived Stromal Cells in Hepatic Function

Abstract
:1. Introduction
2. Results
2.1. Hepatic Phenotype of Mouse Induced Hepatic Progenitors
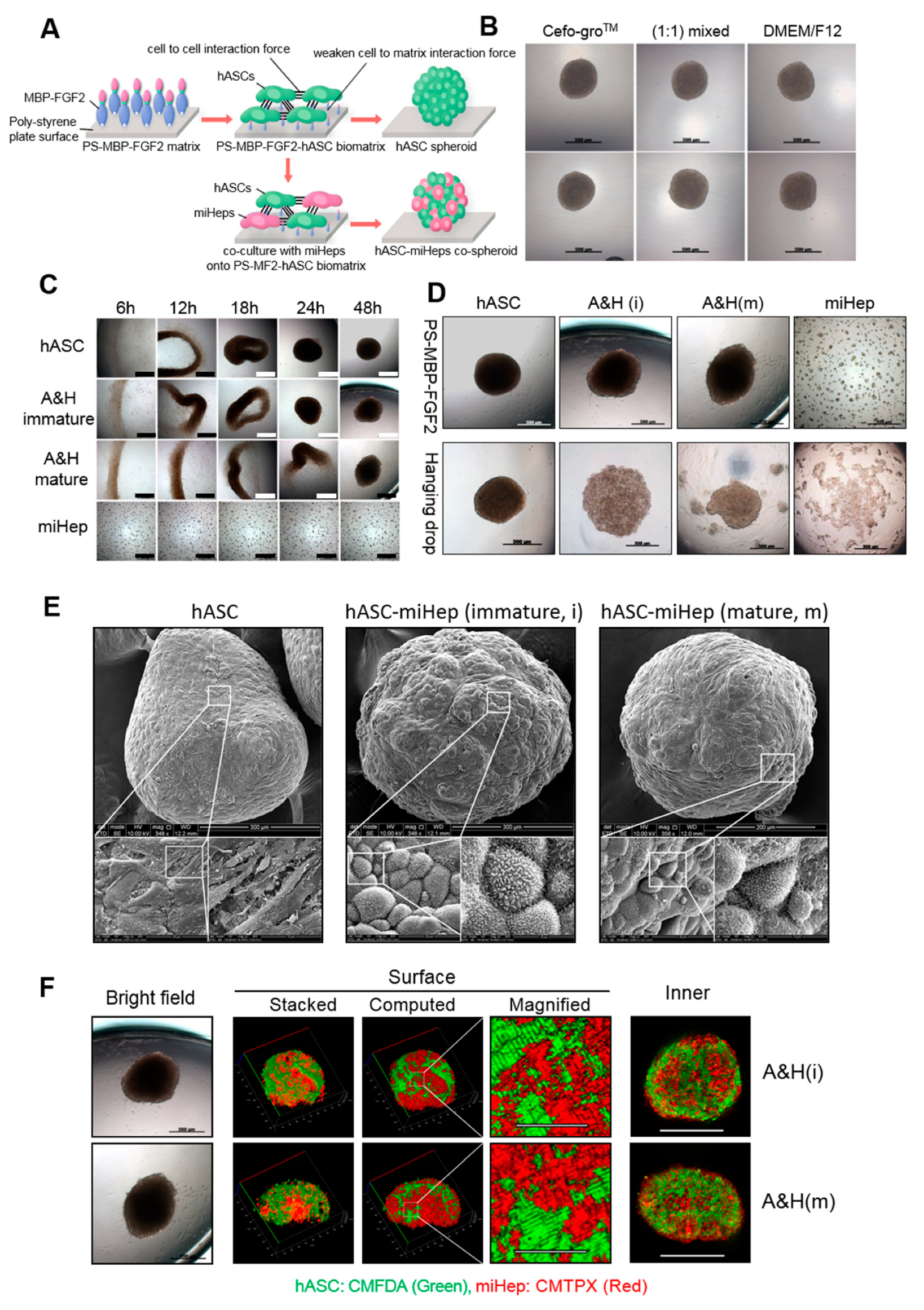
2.2. hASC-miHep Co-Spheroid Formation and Morphological Characteristics
2.3. Effects of hASC Co-Culture Conditions on the Hepatic Function of hASC-miHep Co-Spheroids
2.4. Effects of the hASC Secretome on the Hepatic Phenotype of hASC-miHep Co-Spheroids
2.5. Effects of the Secretome of Other Stromal Cells on miHep Regulation
2.6. Effects of HIF1α on the Hepatic Phenotype via the hASC Secretome
3. Discussion
4. Materials and Methods
4.1. Cells and Reagents
4.2. Cell Culture
4.3. RNA Interference
4.4. MBP-FGF2 Surface Preparation
4.5. Hepatic Microtissue Formation
4.6. Antibodies
4.7. Immunoblotting Assay
4.8. Semi-Quantitative Reverse Transcription Polymerase Chain Reaction
4.9. RT-PCR
4.10. Immunocytochemistry
4.11. Cell Distribution Analysis
4.12. Scanning Electron Microscopy
4.13. Conditioned Media Assay
4.14. Quantitative Densitometric Analysis
4.15. Growth Factor Analysis
4.16. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ALB | albumin |
| ASGR1 | asialoglycoprotein receptor 1 |
| CMFDA | 5-chloromethylfluorescein diacetate |
| CMTPX | chloromethyl 6-(4(5)-amino2-carboxyphenyl)-1,2,2,4,8,10,10,11-octamethyl-1,2,10,11-tetra hydrodipyrido[3,2-b: 20, 30-i] xanthylium |
| CYP | cytochrome P450 |
| DIV | days in vitro |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DMSO | dimethyl sulfoxide |
| E-cad | E-cadherin |
| EGF | epidermal growth factor |
| FGF2 | basic fibroblast growth factor |
| hASCs | human adipose-derived stem cells |
| hBMSCs | human bone marrow-derived mesenchymal cells |
| hDFs | human dermal fibroblasts |
| HGF | hepatocyte growth factor |
| HIF1α | hypoxia-inducible factor 1α |
| HNF4α | hepatocyte nuclear factor 4α |
| IGF | insulin growth factor |
| IGFBP-1 | insulin-like growth factor-binding protein-1 |
| miHeps | mouse induced hepatic precursor cells |
| MEFs | mouse embryonic fibroblasts |
| PFA | paraformaldehyde |
| PBS | phosphate-buffered saline |
| PDGF | platelet-derived growth factor |
| PMSF | phenylmethylsulfonyl fluoride |
| PS | polystyrene |
| TGFβ | tumor growth factor β |
| TNFα | tumor necrosis factor α |
| VEGF | vascular endothelial growth factor |
References
- Griffith, L.G.; Swartz, M.A. Capturing complex 3D tissue physiology in vitro. Nat. Rev. Mol. Cell Biol. 2006, 7, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Griffith, L.G.; Naughton, G. Tissue engineering--current challenges and expanding opportunities. Science 2002, 295, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Matsui, T.; Okumura, H.; Nakajima, M.; Nishimura, M.; Naito, S.; Tateno, C.; Yoshizato, K.; Yokoi, T. Expression of human phase II enzymes in chimeric mice with humanized liver. Drug Metab. Dispos. 2005, 33, 1333–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, M.H.; Melvin, M.A.; Shaw, P.; Melvin, W.T.; Burke, M.D. Studies on the maintenance of cytochromes P-450 and b5, monooxygenases and cytochrome reductases in primary cultures of rat hepatocytes. Febs Lett. 1985, 190, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Godoy, P.; Hewitt, N.J.; Albrecht, U.; Andersen, M.E.; Ansari, N.; Bhattacharya, S.; Bode, J.G.; Bolleyn, J.; Borner, C.; Bottger, J.; et al. Recent advances in 2D and 3D in vitro systems using primary hepatocytes, alternative hepatocyte sources and non-parenchymal liver cells and their use in investigating mechanisms of hepatotoxicity, cell signaling and ADME. Arch. Toxicol. 2013, 87, 1315–1530. [Google Scholar] [PubMed] [Green Version]
- Bissell, D.M.; Guzelian, P.S. Phenotypic stability of adult rat hepatocytes in primary monolayer culture. Ann. N. Y. Acad. Sci. 1980, 349, 85–98. [Google Scholar] [CrossRef]
- Sauer, V.; Roy-Chowdhury, N.; Guha, C.; Roy-Chowdhury, J. Induced pluripotent stem cells as a source of hepatocytes. Curr. Pathobiol. Rep. 2014, 2, 11–20. [Google Scholar] [CrossRef]
- Si-Tayeb, K.; Noto, F.K.; Nagaoka, M.; Li, J.; Battle, M.A.; Duris, C.; North, P.E.; Dalton, S.; Duncan, S.A. Highly efficient generation of human hepatocyte-like cells from induced pluripotent stem cells. Hepatology 2010, 51, 297–305. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, S.; Holton, K.L.; Lanza, R. Efficient differentiation of functional hepatocytes from human embryonic stem cells. Stem Cells 2008, 26, 1117–1127. [Google Scholar] [CrossRef] [Green Version]
- Sekiya, S.; Suzuki, A. Direct conversion of mouse fibroblasts to hepatocyte-like cells by defined factors. Nature 2011, 475, 390–393. [Google Scholar] [CrossRef]
- Huang, P.; Zhang, L.; Gao, Y.; He, Z.; Yao, D.; Wu, Z.; Cen, J.; Chen, X.; Liu, C.; Hu, Y.; et al. Direct reprogramming of human fibroblasts to functional and expandable hepatocytes. Cell Stem Cell 2014, 14, 370–384. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Wang, J.; Jia, J.; Song, N.; Xiang, C.; Xu, J.; Hou, Z.; Su, X.; Liu, B.; Jiang, T.; et al. Human hepatocytes with drug metabolic function induced from fibroblasts by lineage reprogramming. Cell Stem Cell 2014, 14, 394–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, R.; Tang, W.; Yuan, Q.; Hui, L.; Wang, X.; Xie, X. Chemical Cocktails Enable Hepatic Reprogramming of Mouse Fibroblasts with a Single Transcription Factor. Stem Cell Rep. 2017, 9, 499–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamori, D.; Akamine, H.; Takayama, K.; Sakurai, F.; Mizuguchi, H. Direct conversion of human fibroblasts into hepatocyte-like cells by ATF5, PROX1, FOXA2, FOXA3, and HNF4A transduction. Sci. Rep. 2017, 7, 16675. [Google Scholar] [CrossRef]
- Lim, H.L.; Hwang, Y.; Kar, M.; Varghese, S. Smart hydrogels as functional biomimetic systems. Biomater. Sci. 2014, 2, 603–618. [Google Scholar] [CrossRef] [PubMed]
- Lutolf, M.P.; Hubbell, J.A. Synthetic biomaterials as instructive extracellular microenvironments for morphogenesis in tissue engineering. Nat. Biotechnol. 2005, 23, 47–55. [Google Scholar] [CrossRef]
- Murphy, W.L.; McDevitt, T.C.; Engler, A.J. Materials as stem cell regulators. Nat. Mater. 2014, 13, 547–557. [Google Scholar] [CrossRef]
- Gauvin, R.; Chen, Y.C.; Lee, J.W.; Soman, P.; Zorlutuna, P.; Nichol, J.W.; Bae, H.; Chen, S.; Khademhosseini, A. Microfabrication of complex porous tissue engineering scaffolds using 3D projection stereolithography. Biomaterials 2012, 33, 3824–3834. [Google Scholar] [CrossRef] [Green Version]
- Sasai, Y. Next-generation regenerative medicine: Organogenesis from stem cells in 3D culture. Cell Stem Cell 2013, 12, 520–530. [Google Scholar] [CrossRef] [Green Version]
- Jakab, K.; Norotte, C.; Marga, F.; Murphy, K.; Vunjak-Novakovic, G.; Forgacs, G. Tissue engineering by self-assembly and bio-printing of living cells. Biofabrication 2010, 2, 022001. [Google Scholar] [CrossRef]
- Thomas, R.J.; Bhandari, R.; Barrett, D.A.; Bennett, A.J.; Fry, J.R.; Powe, D.; Thomson, B.J.; Shakesheff, K.M. The effect of three-dimensional co-culture of hepatocytes and hepatic stellate cells on key hepatocyte functions in vitro. Cells Tissues Organs 2005, 181, 67–79. [Google Scholar] [CrossRef]
- Sharma, R.; Greenhough, S.; Medine, C.N.; Hay, D.C. Three-dimensional culture of human embryonic stem cell derived hepatic endoderm and its role in bioartificial liver construction. J. Biomed. Biotechnol. 2010, 2010, 236147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dash, A.; Inman, W.; Hoffmaster, K.; Sevidal, S.; Kelly, J.; Obach, R.S.; Griffith, L.G.; Tannenbaum, S.R. Liver tissue engineering in the evaluation of drug safety. Expert Opin. Drug Metab. Toxicol. 2009, 5, 1159–1174. [Google Scholar] [CrossRef]
- Godier, A.F.; Marolt, D.; Gerecht, S.; Tajnsek, U.; Martens, T.P.; Vunjak-Novakovic, G. Engineered microenvironments for human stem cells. Birth Defects Res. C Embryo Today 2008, 84, 335–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riccalton-Banks, L.; Liew, C.; Bhandari, R.; Fry, J.; Shakesheff, K. Long-term culture of functional liver tissue: Three-dimensional coculture of primary hepatocytes and stellate cells. Tissue Eng. 2003, 9, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Goulet, F.; Normand, C.; Morin, O. Cellular interactions promote tissue-specific function, biomatrix deposition and junctional communication of primary cultured hepatocytes. Hepatology 1988, 8, 1010–1018. [Google Scholar] [CrossRef]
- Morin, O.; Normand, C. Long-term maintenance of hepatocyte functional activity in co-culture: Requirements for sinusoidal endothelial cells and dexamethasone. J. Cell Physiol. 1986, 129, 103–110. [Google Scholar] [CrossRef]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived stem cells: Isolation, expansion and differentiation. Methods 2008, 45, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Dai, R.; Wang, Z.; Samanipour, R.; Koo, K.I.; Kim, K. Adipose-Derived Stem Cells for Tissue Engineering and Regenerative Medicine Applications. Stem Cells Int. 2016, 2016, 6737345. [Google Scholar] [CrossRef] [Green Version]
- Linero, I.; Chaparro, O. Paracrine effect of mesenchymal stem cells derived from human adipose tissue in bone regeneration. PLoS ONE 2014, 9, e107001. [Google Scholar] [CrossRef] [Green Version]
- Petrenko, Y.; Sykova, E.; Kubinova, S. The therapeutic potential of three-dimensional multipotent mesenchymal stromal cell spheroids. Stem Cell Res. 2017, 8, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, D.; Tripisciano, C.; Weber, V.; Dominici, M.; Kasper, C. Dynamic Cultivation of Mesenchymal Stem Cell Aggregates. Bioengineering (Basel) 2018, 5, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.S.; Vincent, L.G.; Lee, A.R.; Kretchmer, K.C.; Chirasatitsin, S.; Dobke, M.K.; Engler, A.J. The alignment and fusion assembly of adipose-derived stem cells on mechanically patterned matrices. Biomaterials 2012, 33, 6943–6951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Park, I.S.; Park, Y.; Jung, Y.; Kim, S.H.; Kim, S.H. Therapeutic angiogenesis of three-dimensionally cultured adipose-derived stem cells in rat infarcted hearts. Cytotherapy 2013, 15, 542–556. [Google Scholar] [CrossRef] [PubMed]
- Rajangam, T.; Park, M.H.; Kim, S.H. 3D Human Adipose-Derived Stem Cell Clusters as a Model for In Vitro Fibrosis. Tissue Eng. Part C Methods 2016, 22, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.M.; Han, M.; Park, I.S.; Jung, Y.; Kim, S.H.; Kim, S.H. Adhesion and differentiation of adipose-derived stem cells on a substrate with immobilized fibroblast growth factor. Acta Biomater. 2012, 8, 1759–1767. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Kim, Y.; Jeon, H.; Lee, S.B.; Kim, J.S.; Park, S.A.; Kim, W.D.; Yang, H.M.; Kim, S.J.; Jeong, J.; et al. Three-Dimensional Bioprinting of Hepatic Structures with Directly Converted Hepatocyte-Like Cells. Tissue Eng. Part A 2018, 24, 576–583. [Google Scholar] [CrossRef]
- Arterburn, L.M.; Zurlo, J.; Yager, J.D.; Overton, R.M.; Heifetz, A.H. A morphological study of differentiated hepatocytes in vitro. Hepatology 1995, 22, 175–187. [Google Scholar] [CrossRef]
- Amable, P.R.; Teixeira, M.V.; Carias, R.B.; Granjeiro, J.M.; Borojevic, R. Protein synthesis and secretion in human mesenchymal cells derived from bone marrow, adipose tissue and Wharton’s jelly. Stem Cell Res. 2014, 5, 53. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Hoogduijn, M.J.; Baan, C.C.; Korevaar, S.S.; de Kuiper, R.; Yan, L.; Wang, L.; van Besouw, N.M. Adipose Tissue-Derived Mesenchymal Stem Cells Have a Heterogenic Cytokine Secretion Profile. Stem Cells Int. 2017, 2017, 4960831. [Google Scholar] [CrossRef]
- Kilroy, G.E.; Foster, S.J.; Wu, X.; Ruiz, J.; Sherwood, S.; Heifetz, A.; Ludlow, J.W.; Stricker, D.M.; Potiny, S.; Green, P.; et al. Cytokine profile of human adipose-derived stem cells: Expression of angiogenic, hematopoietic, and pro-inflammatory factors. J. Cell Physiol. 2007, 212, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Park, I.S.; Rhie, J.W.; Kim, S.H. A novel three-dimensional adipose-derived stem cell cluster for vascular regeneration in ischemic tissue. Cytotherapy 2014, 16, 508–522. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.C.; Hendriks, D.F.; Moro, S.M.; Ellis, E.; Walsh, J.; Renblom, A.; Fredriksson Puigvert, L.; Dankers, A.C.; Jacobs, F.; Snoeys, J.; et al. Characterization of primary human hepatocyte spheroids as a model system for drug-induced liver injury, liver function and disease. Sci. Rep. 2016, 6, 25187. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, S.N.; Underhill, G.H.; Zaret, K.S.; Fox, I.J. Cell and tissue engineering for liver disease. Sci. Transl. Med. 2014, 6, 245sr2. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Hoshiba, T.; Akaike, T. Effect of carbohydrates attached to polystyrene on hepatocyte morphology on sugar-derivatized polystyrene matrices. J. Biomed. Mater. Res. A 2003, 67, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Hori, Y.; Yamamoto, T.; Urashima, T.; Ohara, Y.; Tanaka, H. 3D spheroid cultures improve the metabolic gene expression profiles of HepaRG cells. Biosci. Rep. 2015, 35, e00208. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Song, B.; Hong, S.; Kim, S.H.; Lee, K. Biomimetic 3D Clusters Using Human Adipose Derived Mesenchymal Stem Cells and Breast Cancer Cells: A Study on Migration and Invasion of Breast Cancer Cells. Mol. Pharm. 2016, 13, 2204–2213. [Google Scholar] [CrossRef]
- Luebke-Wheeler, J.L.; Nedredal, G.; Yee, L.; Amiot, B.P.; Nyberg, S.L. E-cadherin protects primary hepatocyte spheroids from cell death by a caspase-independent mechanism. Cell Transpl. 2009, 18, 1281–1287. [Google Scholar] [CrossRef] [Green Version]
- Simian, M.; Bissell, M.J. Organoids: A historical perspective of thinking in three dimensions. J. Cell Biol. 2017, 216, 31–40. [Google Scholar] [CrossRef]
- Strand, D.W.; Franco, O.E.; Basanta, D.; Anderson, A.R.; Hayward, S.W. Perspectives on tissue interactions in development and disease. Curr. Mol. Med. 2010, 10, 95–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, S.N.; Balis, U.J.; Yarmush, M.L.; Toner, M. Effect of cell-cell interactions in preservation of cellular phenotype: Cocultivation of hepatocytes and nonparenchymal cells. Faseb J. 1999, 13, 1883–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klezovitch, O.; Vasioukhin, V. Cadherin signaling: Keeping cells in touch. F1000Res 2015, 4, 550. [Google Scholar] [CrossRef]
- Baker, B.M.; Chen, C.S. Deconstructing the third dimension: How 3D culture microenvironments alter cellular cues. J. Cell Sci. 2012, 125 (Pt 13), 3015–3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazdic, M.; Arsenijevic, A.; Markovic, B.S.; Volarevic, A.; Dimova, I.; Djonov, V.; Arsenijevic, N.; Stojkovic, M.; Volarevic, V. Mesenchymal Stem Cell-Dependent Modulation of Liver Diseases. Int. J. Biol. Sci. 2017, 13, 1109–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, W.; Rubin, J.P.; Marra, K.G. Adipose-derived stem cells: Implications in tissue regeneration. World J. Stem Cells 2014, 6, 312–321. [Google Scholar] [CrossRef]
- Nath, B.; Szabo, G. Hypoxia and hypoxia inducible factors: Diverse roles in liver diseases. Hepatology 2012, 55, 622–633. [Google Scholar] [CrossRef] [Green Version]
- Garg, A.; Newsome, P.N. Bone marrow mesenchymal stem cells and liver regeneration: Believe the hypoxia! Stem Cell Res. 2013, 4, 108. [Google Scholar]
- Schadde, E.; Tsatsaris, C.; Swiderska-Syn, M.; Breitenstein, S.; Urner, M.; Schimmer, R.; Booy, C.; Z’Graggen, B.R.; Wenger, R.H.; Spahn, D.R.; et al. Hypoxia of the growing liver accelerates regeneration. Surgery 2017, 161, 666–679. [Google Scholar] [CrossRef] [Green Version]
- Forsberg, E.A.; Botusan, I.R.; Wang, J.; Peters, V.; Ansurudeen, I.; Brismar, K.; Catrina, S.B. Carnosine decreases IGFBP1 production in db/db mice through suppression of HIF-1. J. Endocrinol. 2015, 225, 159–167. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Dilution Factors | |
|---|---|---|
| Immunoblotting | Immunohistochemistry | |
| β-actin | 1:2000 | - |
| HNF4α | 1:400 | 1:100 |
| ALB | 1:1000 | 1:500 |
| ASGR1 | 1:500 | 1:200 |
| CYP1a2 | 1:500 | 1:100 |
| E-Cad | - | 1:200 |
| Ki-67 | - | 1:500 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, S.; Oh, S.J.; Choi, D.; Hwang, Y.; Kim, S.-H. Self-Organized Liver Microtissue on a Bio-Functional Surface: The Role of Human Adipose-Derived Stromal Cells in Hepatic Function. Int. J. Mol. Sci. 2020, 21, 4605. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134605
Hong S, Oh SJ, Choi D, Hwang Y, Kim S-H. Self-Organized Liver Microtissue on a Bio-Functional Surface: The Role of Human Adipose-Derived Stromal Cells in Hepatic Function. International Journal of Molecular Sciences. 2020; 21(13):4605. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134605
Chicago/Turabian StyleHong, Seokheon, Seung Ja Oh, Dongho Choi, Yongsung Hwang, and Sang-Heon Kim. 2020. "Self-Organized Liver Microtissue on a Bio-Functional Surface: The Role of Human Adipose-Derived Stromal Cells in Hepatic Function" International Journal of Molecular Sciences 21, no. 13: 4605. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134605