CPPF, A Novel Microtubule Targeting Anticancer Agent, Inhibits the Growth of a Wide Variety of Cancers


Abstract
:1. Introduction
2. Results
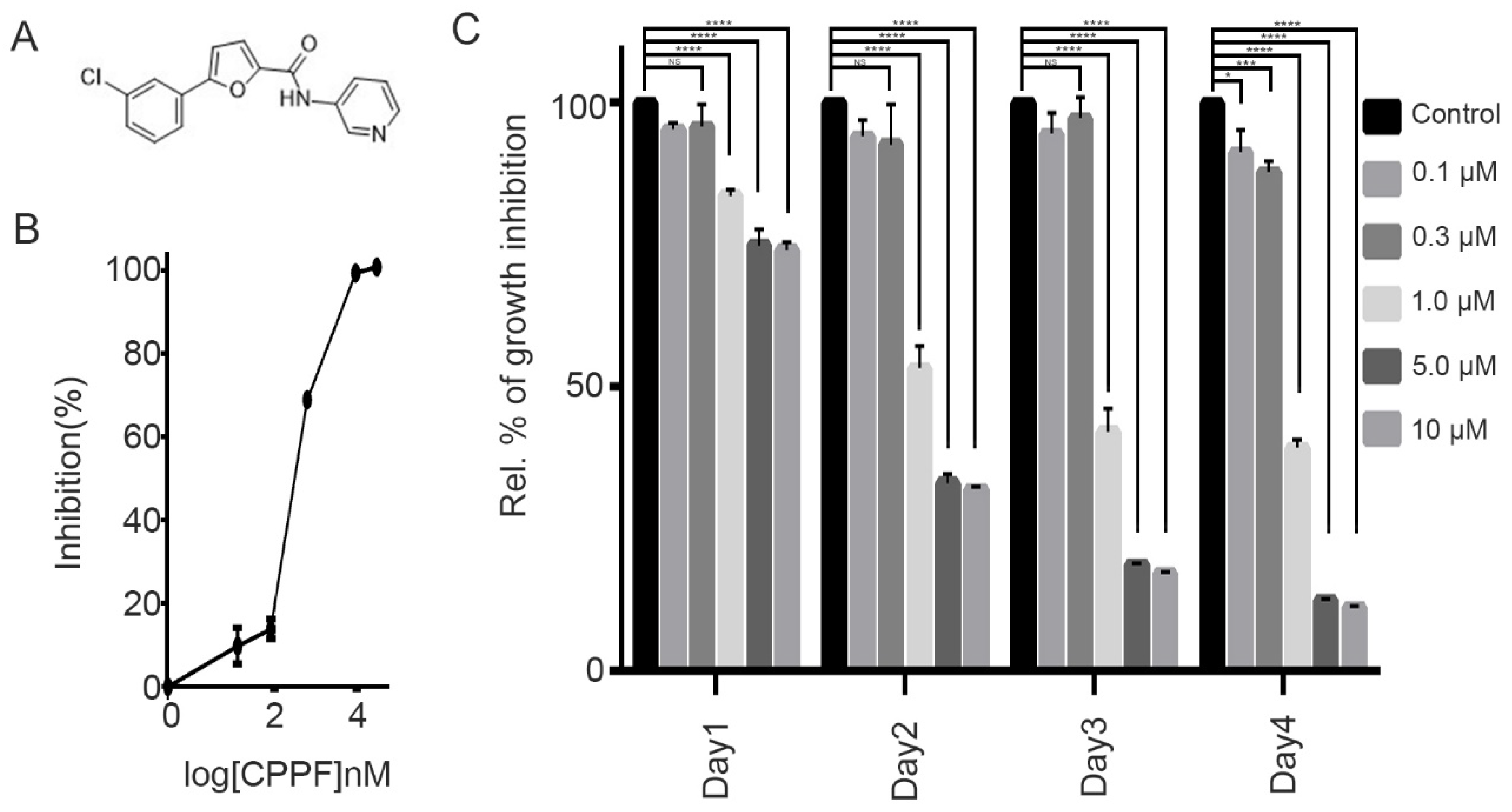
2.1. CPPF Inhibits Cell Proliferation of Various Cancer Cell Lines
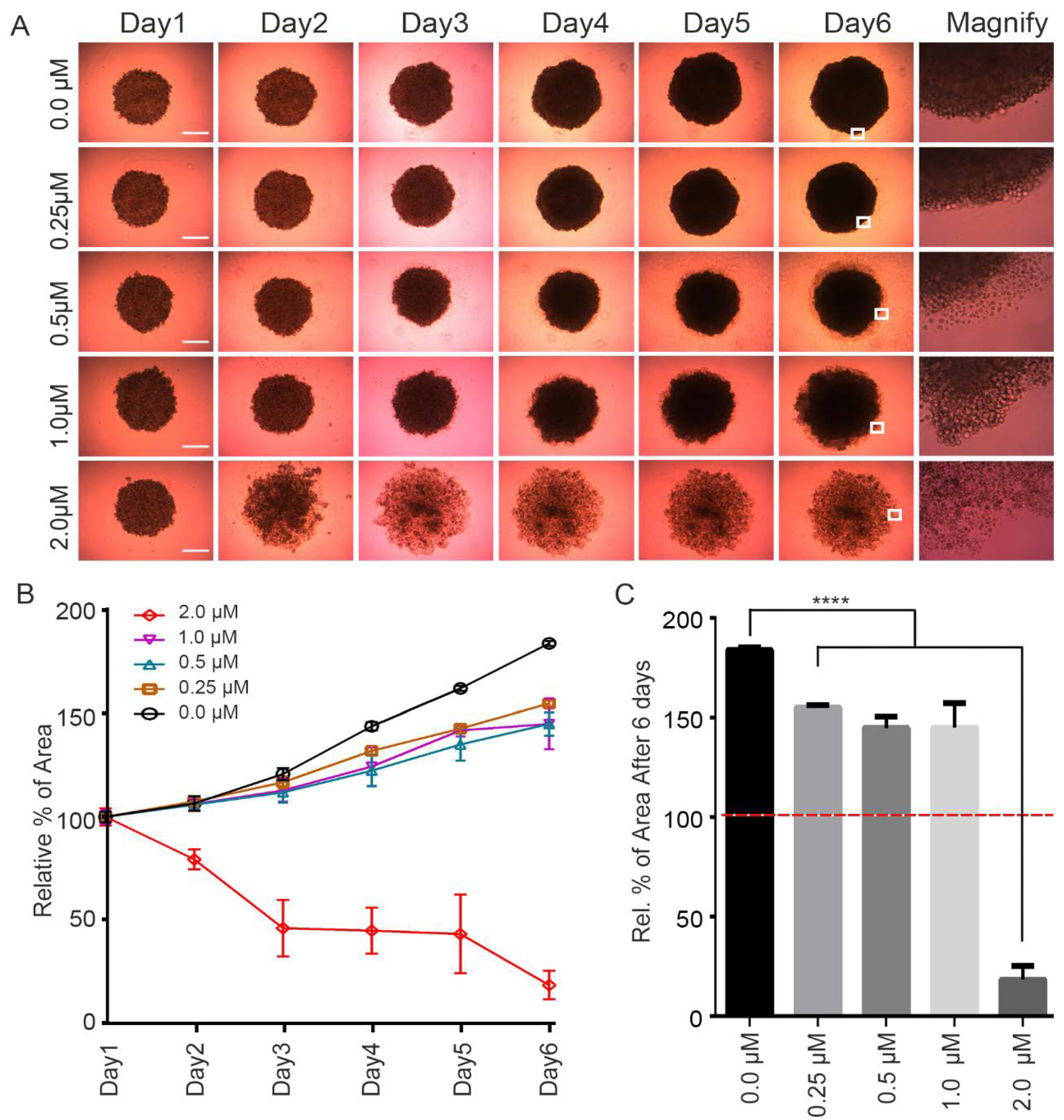
2.2. CPPF Inhibits Growth of 3D Tumor Spheroid Cultures
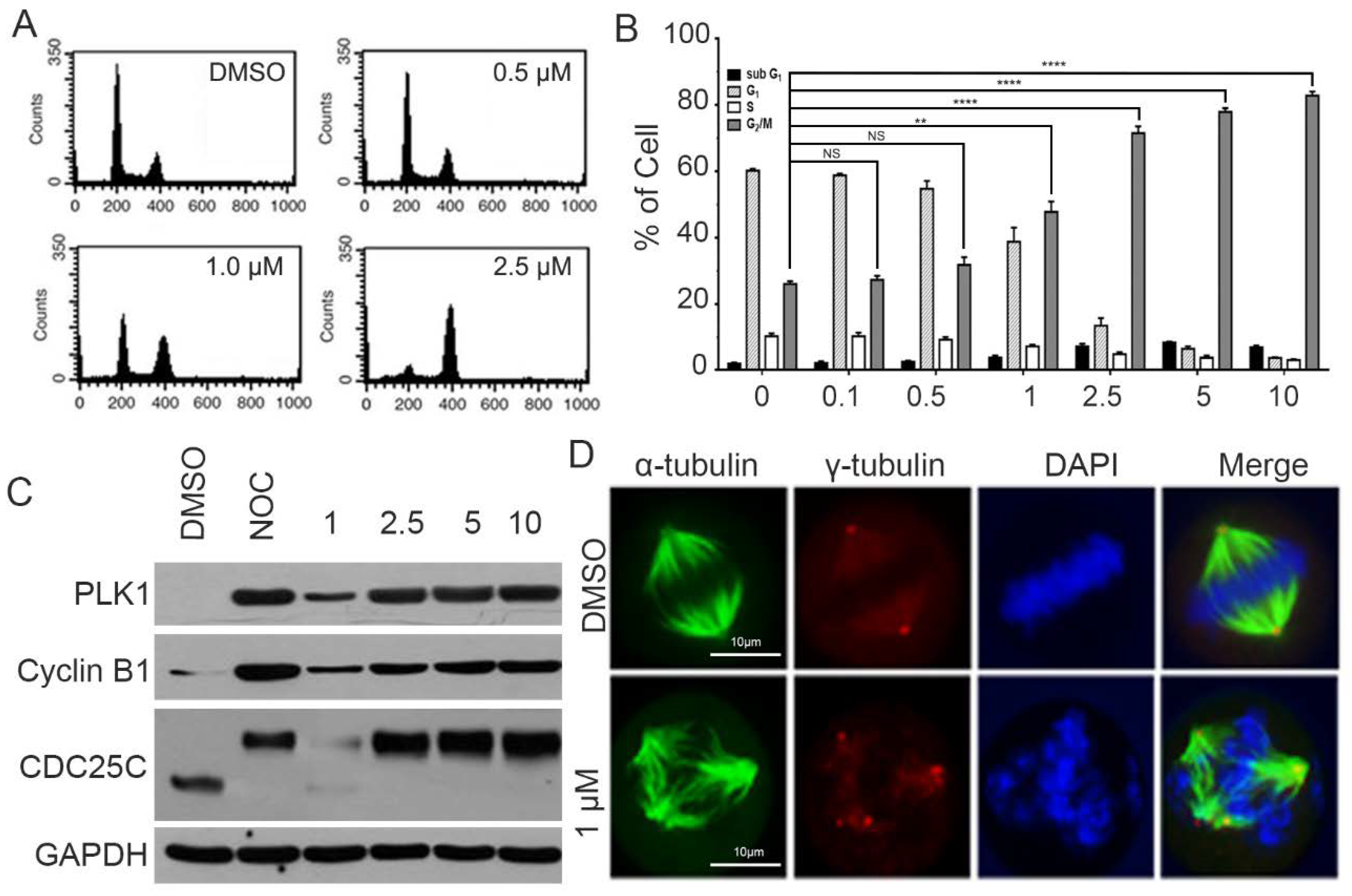
2.3. CPPF Causes Mitotic Arrest
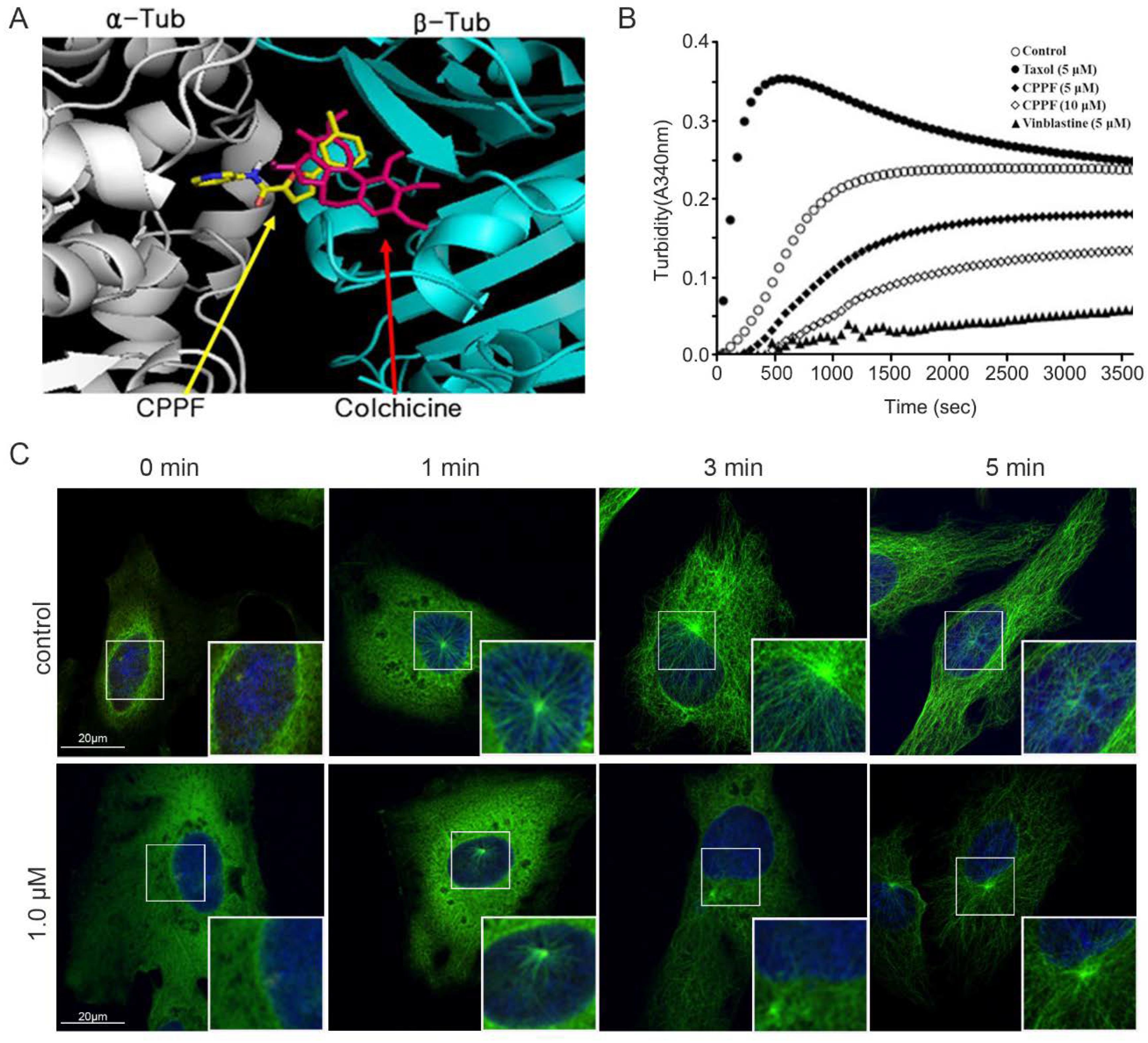
2.4. CPPF Targets Microtubules and Inhibits Tubulin Dynamics
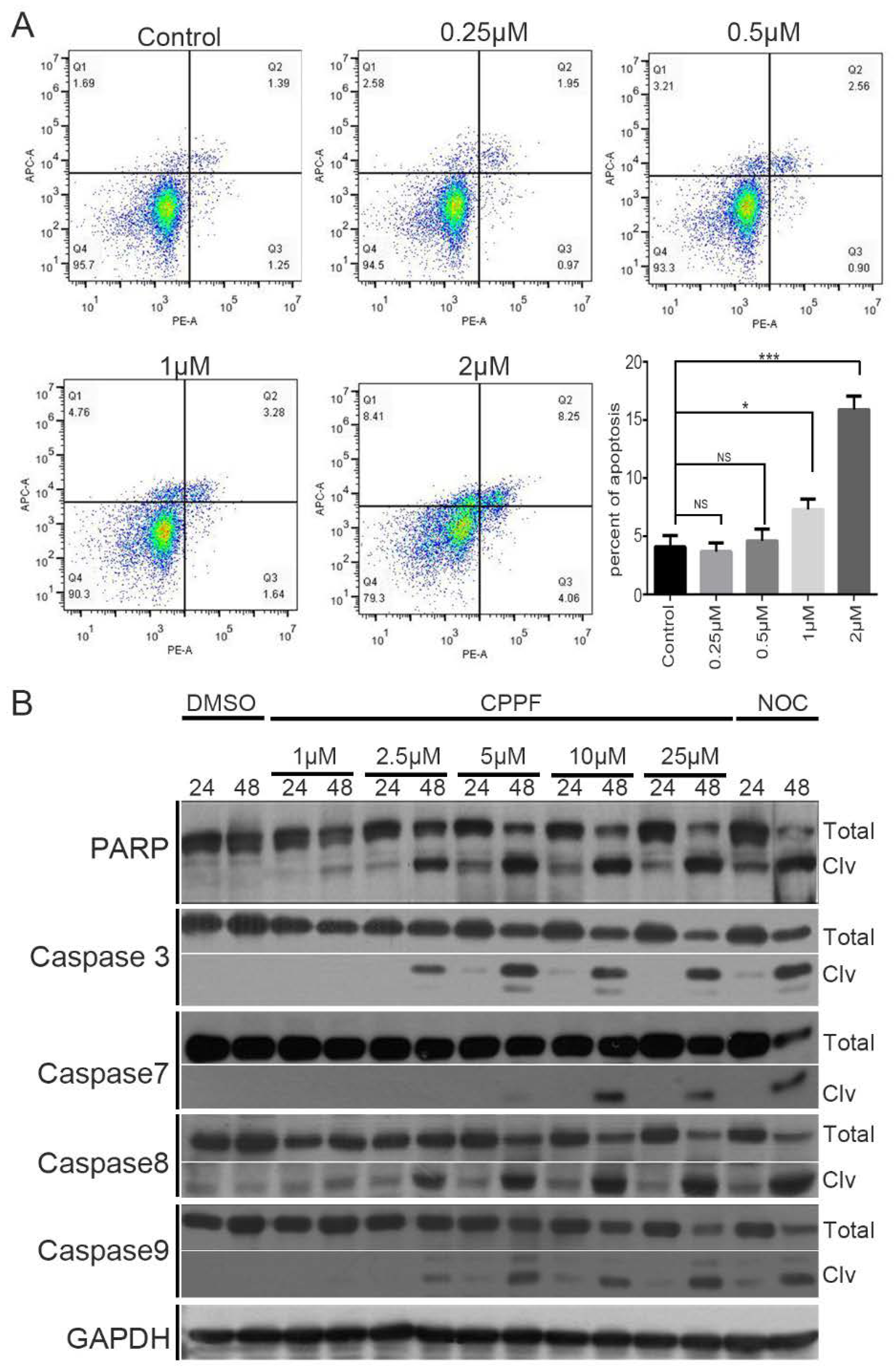
2.5. CPPF Induces Apoptosis
2.6. CPPF Is Effective Against Multidrug Resistance Cell Lines
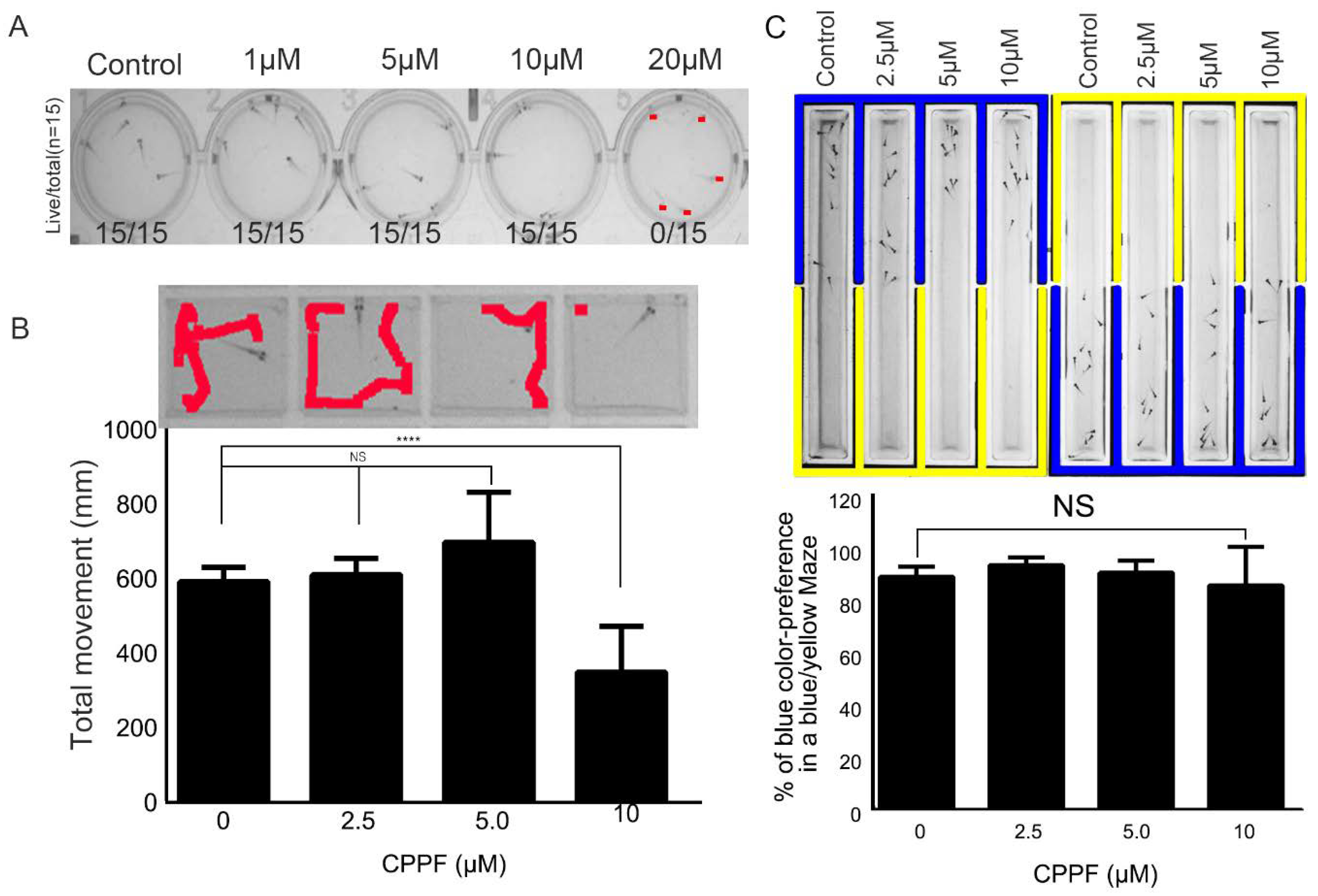
2.7. CPPF Has Low Toxicity As Shown By Zebrafish Testing
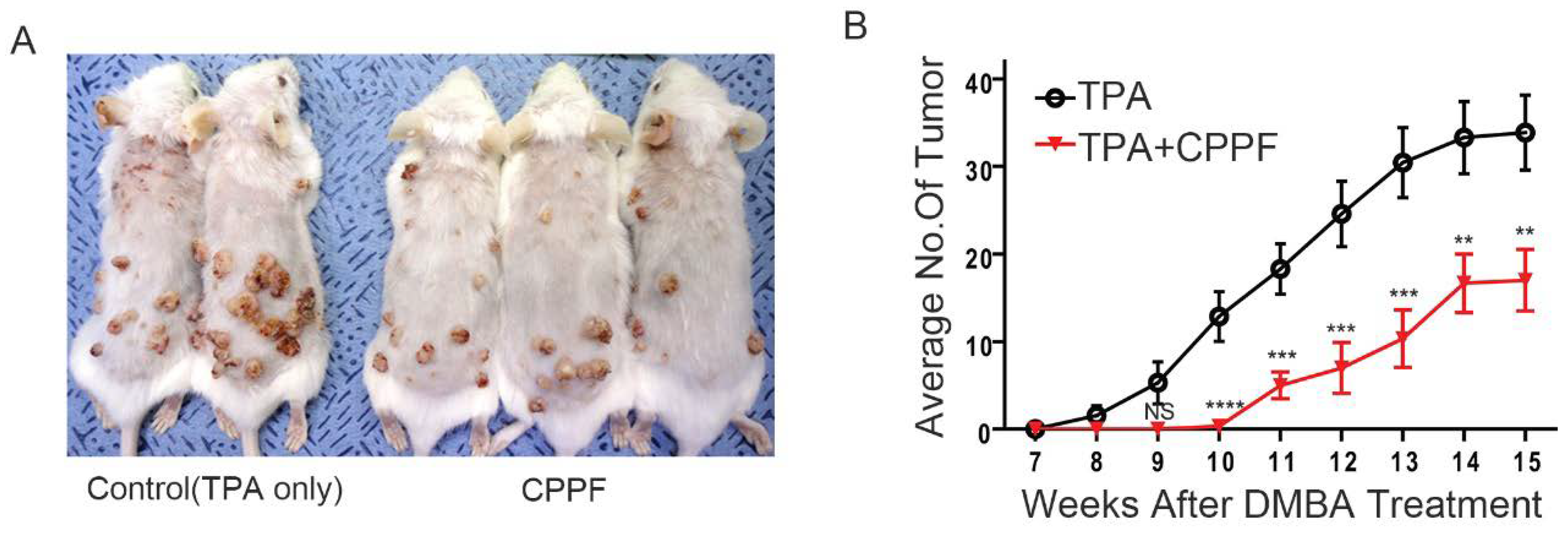
2.8. CPPF Is Effective at Treating and Preventing Cancer in Mice Models
3. Discussion
4. Materials and Methods
4.1. Synthesis of CPPF
4.2. Chemicals and Antibodies
4.3. Cell Lines
4.4. MTT Assay
4.5. Three-Dimensional Culture
4.6. Flow Cytometric Analysis
4.7. Western Blot
4.8. Tubulin Depolymerization Assay
4.9. Computer Modeling Study
4.10. Immunofluorescence Microscopy
4.11. Cold Shock Tubulin Growth Assay
4.12. Zebrafish Toxicity Test
4.13. Zebrafish Behavioral Test
4.14. Carcinogen-Induced Skin Cancer
4.15. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MTA | Microtubule target agent |
| MSAs | Microtubule stabilizer agents |
| MDAs | Microtubule destabilizer agents |
| CPPF | 5-(3-cchlorophenyl)-N-(3-pyridinyl)-2-furamide |
| DMBA | 7,12-dimethylbenz[α] anthracene |
| TPA | Phorbol ester 12-O-tetradecanoylphorbol-13-acetate |
| PLK1 | Polo-like kinase 1 |
| CyclinB1 | G2/mitotic-specific cyclin-B1 |
| CDC25C | M-phase inducer phosphatase 3 |
| PI | Propidium iodide |
| PARP | Poly (ADP-ribose) polymerase |
| ADR | Adriamycin resistance |
| MDR | Multidrug resistance |
| DPF | Day post fertilization |
References
- DeVita, V.T., Jr.; Chu, E. A history of cancer chemotherapy. Cancer Res. 2008, 68, 8643–8653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frei, E., 3rd; Franzino, A.; Shnider, B.I.; Costa, G.; Colsky, J.; Brindley, C.O.; Hosley, H.; Holland, J.F.; Gold, G.L.; Jonsson, U. Clinical studies of vinblastine. Cancer Chemother. Rep. 1961, 12, 125–129. [Google Scholar] [PubMed]
- Manfredi, J.J.; Horwitz, S.B. Taxol: An antimitotic agent with a new mechanism of action. Pharmacol. Ther. 1984, 25, 83–125. [Google Scholar] [CrossRef]
- Giannakakou, P.; Villalba, L.; Li, H.; Poruchynsky, M.; Fojo, T. Combinations of paclitaxel and vinblastine and their effects on tubulin polymerization and cellular cytotoxicity: Characterization of a synergistic schedule. Int. J. Cancer 1998, 75, 57–63. [Google Scholar] [CrossRef]
- Dogterom, M.; Koenderink, G.H. Actin-microtubule crosstalk in cell biology. Nat. Rev. Mol. Cell Biol. 2019, 20, 38–54. [Google Scholar] [CrossRef]
- Gadadhar, S.; Bodakuntla, S.; Natarajan, K.; Janke, C. The tubulin code at a glance. J. Cell Sci. 2017, 130, 1347–1353. [Google Scholar] [CrossRef] [Green Version]
- Brouhard, G.J.; Rice, L.M. Microtubule dynamics: An interplay of biochemistry and mechanics. Nat. Rev. Mol. Cell Biol. 2018, 19, 451–463. [Google Scholar] [CrossRef]
- Jordan, M.A.; Wilson, L. Microtubules as a target for anticancer drugs. Nat. Rev. Cancer 2004, 4, 253–265. [Google Scholar] [CrossRef]
- Steinmetz, M.O.; Prota, A.E. Microtubule-Targeting Agents: Strategies To Hijack the Cytoskeleton. Trends Cell Biol. 2018, 28, 776–792. [Google Scholar] [CrossRef]
- Watanabe, H.; Watanabe, H.; Usui, T.; Kondoh, M.; Osada, H.; Kitahara, T. Synthesis of pironetin and related analogs: Studies on structure-activity relationships as tubulin assembly inhibitors. J. Antibiot. (Tokyo) 2000, 53, 540–545. [Google Scholar] [CrossRef] [Green Version]
- Ravelli, R.B.; Gigant, B.; Curmi, P.A.; Jourdain, I.; Lachkar, S.; Sobel, A.; Knossow, M. Insight into tubulin regulation from a complex with colchicine and a stathmin-like domain. Nature 2004, 428, 198–202. [Google Scholar] [CrossRef]
- Fanale, D.; Bronte, G.; Passiglia, F.; Calo, V.; Castiglia, M.; Di Piazza, F.; Barraco, N.; Cangemi, A.; Catarella, M.T.; Insalaco, L.; et al. Stabilizing versus destabilizing the microtubules: A double-edge sword for an effective cancer treatment option? Anal. Cell Pathol. (Amst) 2015, 2015, 690916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.G.; Horwitz, S.B. Differential mitotic responses to microtubule-stabilizing and -destabilizing drugs. Cancer Res. 2002, 62, 1935–1938. [Google Scholar] [PubMed]
- Ramirez, L.Y.; Huestis, S.E.; Yap, T.Y.; Zyzanski, S.; Drotar, D.; Kodish, E. Potential chemotherapy side effects: What do oncologists tell parents? Pediatr. Blood Cancer 2009, 52, 497–502. [Google Scholar] [CrossRef] [Green Version]
- Gillet, J.P.; Gottesman, M.M. Mechanisms of multidrug resistance in cancer. Methods Mol. Biol. 2010, 596, 47–76. [Google Scholar] [PubMed]
- Guy, G.P., Jr.; Machlin, S.R.; Ekwueme, D.U.; Yabroff, K.R. Prevalence and costs of skin cancer treatment in the U.S., 2002-2006 and 2007-2011. Am. J. Prev. Med. 2015, 48, 183–187. [Google Scholar] [CrossRef]
- Abel, E.L.; Angel, J.M.; Kiguchi, K.; DiGiovanni, J. Multi-stage chemical carcinogenesis in mouse skin: Fundamentals and applications. Nat. Protoc. 2009, 4, 1350–1362. [Google Scholar] [CrossRef]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-dimensional cell culture systems and their applications in drug discovery and cell-based biosensors. Assay. Drug. Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, O.M.; Le Berre, M.; Dimitracopoulos, A.; Bonazzi, D.; Zlotek-Zlotkiewicz, E.; Picone, R.; Duke, T.; Piel, M.; Baum, B. Mitotic rounding alters cell geometry to ensure efficient bipolar spindle formation. Dev. Cell 2013, 25, 270–283. [Google Scholar] [CrossRef] [Green Version]
- Schmit, T.L.; Ahmad, N. Regulation of mitosis via mitotic kinases: New opportunities for cancer management. Mol. Cancer Ther. 2007, 6, 1920–1931. [Google Scholar] [CrossRef] [Green Version]
- Olziersky, A.M.; Labidi-Galy, S.I. Clinical Development of Anti-mitotic Drugs in Cancer. Adv. Exp. Med. Biol. 2017, 1002, 125–152. [Google Scholar] [PubMed]
- Bergen, L.G.; Borisy, G.G. Tubulin-colchicine complex (TC) inhibits microtubule depolymerization by a capping reaction exerted preferentially at the minus end. J. Cell Biochem. 1986, 30, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.L.; Kavallaris, M.; McCarroll, J.A. Microtubules and their role in cellular stress in cancer. Front. Oncol. 2014, 4, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzoor, S.; Bilal, A.; Khan, S.; Ullah, R.; Iftikhar, S.; Emwas, A.H.; Alazmi, M.; Gao, X.; Jawaid, A.; Saleem, R.S.Z.; et al. Identification and characterization of SSE15206, a microtubule depolymerizing agent that overcomes multidrug resistance. Sci. Rep. 2018, 8, 3305. [Google Scholar] [CrossRef] [Green Version]
- Manchado, E.; Guillamot, M.; Malumbres, M. Killing cells by targeting mitosis. Cell Death Differ. 2012, 19, 369–377. [Google Scholar] [CrossRef]
- Kang, M.R.; Lee, K.; Kang, J.S.; Lee, C.W.; Lee, K.H.; Kim, J.H.; Yang, J.W.; Kim, B.G.; Han, G.; Kang, J.S.; et al. KBH-A42, a histone deacetylase inhibitor, inhibits the growth of doxorubicin-resistant leukemia cells expressing P-glycoprotein. Oncol. Rep. 2010, 23, 801–809. [Google Scholar]
- Kim, H.M.; Oh, G.T.; Hong, D.H.; Kim, M.S.; Kang, J.S.; Park, S.M.; Han, S.B. MDR-1 gene expression is a minor factor in determining the multidrug resistance phenotype of MCF7/ADR and KB-V1 cells. FEBS Lett. 1997, 412, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.S.; Hwang, K.S.; Yang, J.Y.; Chae, J.S.; Kim, G.R.; Kan, H.; Jung, M.H.; Lee, H.Y.; Song, J.S.; Ahn, S.; et al. Neurochemical and behavioral analysis by acute exposure to bisphenol A in zebrafish larvae model. Chemosphere 2020, 239, 124751. [Google Scholar] [CrossRef]
- Park, J.S.; Ryu, J.H.; Choi, T.I.; Bae, Y.K.; Lee, S.; Kang, H.J.; Kim, C.H. Innate Color Preference of Zebrafish and Its Use in Behavioral Analyses. Mol. Cells 2016, 39, 750–755. [Google Scholar] [CrossRef] [Green Version]
- Shan, Y.; Wei, Z.; Tao, L.; Wang, S.; Zhang, F.; Shen, C.; Wu, H.; Liu, Z.; Zhu, P.; Wang, A.; et al. Prophylaxis of Diallyl Disulfide on Skin Carcinogenic Model via p21-dependent Nrf2 stabilization. Sci. Rep. 2016, 6, 35676. [Google Scholar] [CrossRef]
- Gupta, S.C.; Sung, B.; Prasad, S.; Webb, L.J.; Aggarwal, B.B. Cancer drug discovery by repurposing: Teaching new tricks to old dogs. Trends Pharmacol. Sci. 2013, 34, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Masimirembwa, C.M.; Bredberg, U.; Andersson, T.B. Metabolic stability for drug discovery and development: Pharmacokinetic and biochemical challenges. Clin. Pharmacokinet 2003, 42, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Szakacs, G.; Paterson, J.K.; Ludwig, J.A.; Booth-Genthe, C.; Gottesman, M.M. Targeting multidrug resistance in cancer. Nat. Rev. Drug. Discov. 2006, 5, 219–234. [Google Scholar] [CrossRef]
- Zoetemelk, M.; Rausch, M.; Colin, D.J.; Dormond, O.; Nowak-Sliwinska, P. Short-term 3D culture systems of various complexity for treatment optimization of colorectal carcinoma. Sci. Rep. 2019, 9, 7103. [Google Scholar] [CrossRef] [PubMed]
- Orth, J.D.; Loewer, A.; Lahav, G.; Mitchison, T.J. Prolonged mitotic arrest triggers partial activation of apoptosis, resulting in DNA damage and p53 induction. Mo.l Biol. Cell 2012, 23, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The Different Mechanisms of Cancer Drug Resistance: A Brief Review. Adv. Pharm. Bull 2017, 7, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, A.L. Zebrafish assays for drug toxicity screening. Expert. Opin. Drug. Metab. Toxicol. 2006, 2, 231–240. [Google Scholar] [CrossRef]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Watson, M.; Holman, D.M.; Maguire-Eisen, M. Ultraviolet Radiation Exposure and Its Impact on Skin Cancer Risk. Semin. Oncol. Nurs. 2016, 32, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Henriksen, T.; Dahlback, A.; Larsen, S.H.; Moan, J. Ultraviolet-radiation and skin cancer. Effect of an ozone layer depletion. Photochem. Photobiol. 1990, 51, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, G.; Nova, J.; Rodriguez-Hernandez, A.E.; Medina, R.D.; Solorzano-Restrepo, C.; Gonzalez, J.; Olmos, M.; Godfrey, K.; Arevalo-Rodriguez, I. Sun protection for preventing basal cell and squamous cell skin cancers. Cochrane Database Syst. Rev. 2016, 7, CD011161. [Google Scholar] [CrossRef] [PubMed]
- Sakchaisri, K.; Kim, S.O.; Hwang, J.; Soung, N.K.; Lee, K.H.; Choi, T.W.; Lee, Y.; Park, C.M.; Thimmegowda, N.R.; Lee, P.Y.; et al. Anticancer activity of a novel small molecule tubulin inhibitor STK899704. PLoS ONE 2017, 12, e0173311. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Saez-Calvo, G.; Olieric, N.; de Asis Balaguer, F.; Barasoain, I.; Lamberth, C.; Diaz, J.F.; Steinmetz, M.O. Quinolin-6-Yloxyacetamides Are Microtubule Destabilizing Agents That Bind to the Colchicine Site of Tubulin. Int. J. Mol. Sci. 2017, 18, 1336. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Shin, Y.; Choe, H.; Hong, S. Computational design and discovery of nanomolar inhibitors of IkappaB kinase beta. J. Am. Chem. Soc. 2015, 137, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Hennings, H.; Glick, A.B.; Lowry, D.T.; Krsmanovic, L.S.; Sly, L.M.; Yuspa, S.H. FVB/N mice: An inbred strain sensitive to the chemical induction of squamous cell carcinomas in the skin. Carcinogenesis 1993, 14, 2353–2358. [Google Scholar] [CrossRef]
- Liu, H.; Hwang, J.; Li, W.; Choi, T.W.; Liu, K.; Huang, Z.; Jang, J.H.; Thimmegowda, N.R.; Lee, K.W.; Ryoo, I.J.; et al. A derivative of chrysin suppresses two-stage skin carcinogenesis by inhibiting mitogen- and stress-activated kinase 1. Cancer Prev. Res. (Phila) 2014, 7, 74–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Lines | CPPF (µM) | Paclitaxel (µM) | Colchicine (µM) |
|---|---|---|---|
| K562 | 0.386 ± 0.067 | 0.008 ± 0.01 | 0.0123 ± 0.002 |
| K562/ADR | 0.326 ± 0.047 (0.84) | 1.229 ± 0.243 (153.6) | 0.3813 ± 0.101 (31) |
| MCF7 | 7.95 ± 0.714 | 0.004 ± 0.002 | 0.012 ± 0.008 |
| MCF7/ADR | 1.91 ± 0.056 (0.24) | 1.573 ± 0.063 (393.2) | 0.446 ± 0.066 (37.1) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, H.J.; Park, C.; Hwang, J.; N.R., T.; Kim, S.-O.; Han, J.; Woo, M.; B, S.; Ryoo, I.-J.; Lee, K.H.; et al. CPPF, A Novel Microtubule Targeting Anticancer Agent, Inhibits the Growth of a Wide Variety of Cancers. Int. J. Mol. Sci. 2020, 21, 4800. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134800
Han HJ, Park C, Hwang J, N.R. T, Kim S-O, Han J, Woo M, B S, Ryoo I-J, Lee KH, et al. CPPF, A Novel Microtubule Targeting Anticancer Agent, Inhibits the Growth of a Wide Variety of Cancers. International Journal of Molecular Sciences. 2020; 21(13):4800. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134800
Chicago/Turabian StyleHan, Ho Jin, Chanmi Park, Joonsung Hwang, Thimmegowda N.R., Sun-Ok Kim, Junyeol Han, Minsik Woo, Shwetha B, In-Ja Ryoo, Kyung Ho Lee, and et al. 2020. "CPPF, A Novel Microtubule Targeting Anticancer Agent, Inhibits the Growth of a Wide Variety of Cancers" International Journal of Molecular Sciences 21, no. 13: 4800. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21134800