Advances in Understanding the Initial Steps of Pruritoceptive Itch: How the Itch Hits the Switch



Abstract
:1. Introduction
2. Types of Itch
3. Itch Mediators, Modulators and their Receptors
3.1. Histamine 1 Receptor
3.2. Serotonin Receptor
3.3. Protease-Activated Receptors
3.4. Neurokinin Receptors
3.5. Bradykinin Receptors
3.6. Calcitonin Gene-Related Peptide Type 1 Receptor
3.7. Mas-related GPR Family Member X1 (MRGPRX1)
3.8. Leukotriene Receptors
3.9. Platelet-Activating Factor Receptor (PAFR)
3.10. Opioid Receptors
3.11. Cannabinoid Receptors
3.12. Interleukin-1 Receptor/Toll-like Receptor Superfamily
3.12.1. Interleukin Receptors
3.12.2. Toll-Like Receptors
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AEA | arachidonoylethanolamide |
| BAM | bovine adrenal medulla |
| BAM8-22 | bovine adrenal medulla peptide (8-22) |
| BLT1, 2 | leukotriene B4 receptor 1 or 2 |
| BDK | bradykinin |
| BDKRB1 | bradykinin receptors 1 or 2 |
| CALCRL | calcitonin gene-related peptide type 1 receptor |
| CB | Cannabinoid |
| CB1 | Cannabinoid receptor 1 or 2 |
| CFA | Complete Freund’s adjuvant |
| CNS | Central nervous system |
| CGPR | Calcitonin gene-related peptide |
| eCB | Endocannabinoids |
| ET-1 | Endothelin-1 |
| GRP | Gastrin-releasing peptide |
| GRPR | Gastrin-releasing peptide receptor |
| GPCR | G protein coupled receptor |
| H1-4R | Histamine receptors 1-4 |
| IL1-14 | Interleukins 1-14 |
| IL1 – 14R | Interleukin 1-14 receptor |
| IP3 | Inositol 1,4,5 triphosphate |
| OPRK | κ- opioid receptor |
| LT | Leukotriene |
| LTB4 | Leukotriene B4 |
| MAPK | Mitogen-activated protein kinase |
| ME | mediator |
| MRGPRX1 | Mas-related GPR family member X1 |
| MO | Modulator |
| MyD88 | Myeloid differentiation primary response 88 |
| OPRM | μ- opioid receptor |
| OSMR | Oncostatin M receptor |
| PAR | Protease-activated receptors |
| PAFR | Platelet-activating factor receptor |
| PIC | Polyinosinic:polycytidylic acid |
| PLCB | Phospholipase CB |
| RAMP1 | Receptor activity-modifying protein |
| SP | Substance P |
| TLR | Toll-like receptor |
| TRIF | TIR-domain-containing adapter-inducing interferon-β |
| TRPA1 | Transient receptor potential cation channel, subfamily A, member 1 |
| TRPV1, 3 | Transient Receptor Potential Vanilloid 1 or 3 |
| TSLP | Thymic stromal lymphopoietin |
| TSLPR | Thymic stromal lymphopoietin receptor |
| 2-AG | 2-Arachidonoylglycerol |
| 5-HTR | 5-Hydroxytryptamine receptor (serotonin receptor) |
References
- Chen, X.-J.; Sun, Y.-G. Central circuit mechanisms of itch. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dalgard, F.J.; Lien, L.; Dalen, I. Itch in the community: Associations with psychosocial factors among adults. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 1215–1219. [Google Scholar] [CrossRef] [PubMed]
- Halvorsen, J.A.; Dalgard, F.; Thoresen, M.; Lien, E. Itch and Mental Distress: A Cross-Sectional Study among Late Adolescents. Acta Derm. Venereol. 2009, 89, 39–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalgard, F.; Svensson, Å.; Holm, J.O.; Sundby, J. Self-Reported Skin Morbidity among Adults: Associations with Quality of Life and General Health in a Norwegian Survey. J. Investig. Dermatol. Symp. Proc. 2004, 9, 120–125. [Google Scholar] [CrossRef] [Green Version]
- Zachariae, R.; Lei, U.; Hædersdal, M.; Zachariae, C. Itch Severity and Quality of Life in Patients with Pruritus: Preliminary Validity of a Danish Adaptation of the Itch Severity Scale. Acta Derm. Venereol. 2012, 92, 508–514. [Google Scholar] [CrossRef] [Green Version]
- Yosipovitch, G.; Papoiu, A.D.P. What causes itch in atopic dermatitis? Curr. Allergy Asthma Rep. 2008, 8, 306–311. [Google Scholar] [CrossRef]
- Yosipovitch, G.; Rosen, J.D.; Hashimoto, T. Itch: From mechanism to (novel) therapeutic approaches. J. Allergy Clin. Immunol. 2018, 142, 1375–1390. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Xian, D.; Yang, L.; Xiong, X.; Lai, R.; Zhong, J. Pruritus: Progress toward Pathogenesis and Treatment. BioMed Res. Int. 2018, 2018, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Andersen, H.; Elberling, J.; Arendt-Nielsen, L. Human Surrogate Models of Histaminergic and Non-histaminergic Itch. Acta Derm. Venereol. 2014, 95, 771–777. [Google Scholar] [CrossRef] [Green Version]
- Patel, T.; Yosipovitch, G. Therapy of pruritus. Expert Opin. Pharmacother. 2010, 11, 1673–1682. [Google Scholar] [CrossRef]
- Twycross, R.; Greaves, M.W.; Handwerker, H.; Jones, E.A.; Libretto, S.E.; Szepietowski, J.C.; Zylicz, Z. Itch: Sratching more than the surface. QJM: Int. J. Med. 2003, 96, 7–26. [Google Scholar] [CrossRef]
- Yosipovitch, G.; Greaves, M.W.; Schmelz, M. Itch. Lancet 2003, 361, 690–694. [Google Scholar] [CrossRef]
- Garibyan, L.; Rheingold, C.G.; Lerner, E.A. Understanding the pathophysiology of itch. Dermatol. Ther. 2013, 26, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Oaklander, A.L. Neuropathic itch. Semin. Cutan. Med. Surg. 2011, 30, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misery, L.; Brenaut, E.; Le Garrec, R.; Abasq, C.; Genestet, S.; Marcorelles, P.; Zagnoli, F. Neuropathic pruritus. Nat. Rev. Neurol. 2014, 10, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Yosipovitch, G.; Samuel, L.S. Neuropathic and psychogenic itch. Dermatol. Ther. 2008, 21, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Tivoli, Y.A.; Rubenstein, R.M. Pruritus: An updated look at an old problem. J. Clin. Aesthet. Dermatol. 2009, 2, 30–36. [Google Scholar]
- Yosipovitch, G. Assessment of itch: More to be learned and improvements to be made. J. Investig. Dermatol. 2003, 121. [Google Scholar] [CrossRef] [Green Version]
- Koma, A.; Steinhoff, M.; Ständer, S.; Yosipovitch, G.; Schmelz, M. The neurobiology of itch. Nat. Rev. Neurosci. 2006, 7, 535–547. [Google Scholar] [CrossRef]
- Rinaldi, G. The Itch-Scratch Cycle: A Review of the Mechanisms. Dermatol. Pr. Concept. 2019, 9, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Tang, Z.; Surdenikova, L.; Kim, S.; Patel, K.N.; Kim, A.; Ru, F.; Guan, Y.; Weng, H.-J.; Geng, Y.; et al. Sensory Neuron-Specific GPCR Mrgprs Are Itch Receptors Mediating Chloroquine-Induced Pruritus. Cell 2009, 139, 1353–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulca, S.; Bayramgürler, D.; Demirsoy, E.O.; Yavuz, M.; Şikar Aktürk, A.; Bilen, N.; Kıran, R. Comparison of effects of 5 and 10 mg oral desloratadine and levocetirizine on histamine-induced wheal and flare response in healthy volunteers. J. Dermatol. Treat. 2013, 24, 473–476. [Google Scholar] [CrossRef]
- Yosipovitch, G.; Bernhard, J.D. Clinical practice. Chronic pruritus. N. Engl. J. Med. 2013, 368, 1625–1634. [Google Scholar] [CrossRef] [PubMed]
- Fowler, E.; Yosipovitch, G. Chronic itch management: Therapies beyond those targeting the immune system. Ann. Allergy Asthma Immunol. 2019, 123, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Kim, B.S. Itch: A Paradigm of Neuroimmune Crosstalk. Immunity 2020, 52, 753–766. [Google Scholar] [CrossRef]
- Ikoma, A.; Handwerker, H.; Miyachi, Y.; Schmelz, M. Electrically evoked itch in humans. Pain 2005, 113, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Dong, X. Peripheral and Central Mechanisms of Itch. Neuron 2018, 98, 482–494. [Google Scholar] [CrossRef]
- Harrison, I.P.; Spada, F. Breaking the Itch-Scratch Cycle: Topical Options for the Management of Chronic Cutaneous Itch in Atopic Dermatitis. Medicine 2019, 6, 76. [Google Scholar] [CrossRef] [Green Version]
- Schmelz, M. Itch and pain. Neurosci. Biobehav. Rev. 2010, 34, 171–176. [Google Scholar] [CrossRef]
- Schmelz, M.; Schmidt, R.; Bickel, A.; Handwerker, H.O.; Torebjork, H.E. Specific C-Receptors for Itch in Human Skin. J. Neurosci. 1997, 17, 8003–8008. [Google Scholar] [CrossRef]
- Sun, S.; Dong, X. Trp channels and itch. Semin. Immunopathol. 2015, 38, 293–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringkamp, M.; Schepers, R.J.; Shimada, S.G.; Johanek, L.M.; Hartke, T.V.; Borzan, J.; Shim, B.; LaMotte, R.H.; Meyer, R.A. A role for nociceptive, myelinated nerve fibers in itch sensation. J. Neurosci. 2011, 31, 14841–14849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usoskin, D.; Furlan, A.; Islam, S.; Abdo, H.; Lönnerberg, P.; Lou, D.; Hjerling-Leffler, J.; Haeggström, J.; Kharchenko, O.; Kharchenko, P.V.; et al. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat. Neurosci. 2014, 18, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-G.; Chen, Z.-F. A gastrin-releasing peptide receptor mediates the itch sensation in the spinal cord. Nature 2007, 448, 700–703. [Google Scholar] [CrossRef]
- Liu, X.Y.; Ginosar, Y.; Yazdi, J.; Hincker, A.; Chen, Z.F. Cross-talk between Human Spinal Cord mu-opioid Receptor 1Y Isoform and Gastrin-releasing Peptide Receptor Mediates Opioid-induced Scratching Behavior. Anesthesiology 2019, 131, 381–391. [Google Scholar] [CrossRef]
- Munanairi, A.; Liu, X.-Y.; Barry, D.M.; Yang, Q.; Yin, J.-B.; Jin, H.; Li, H.; Meng, Q.-T.; Peng, J.-H.; Wu, Z.-Y.; et al. Non-canonical Opioid Signaling Inhibits Itch Transmission in the Spinal Cord of Mice. Cell Rep. 2018, 23, 866–877. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-Y.; Liu, Z.-C.; Sun, Y.-G.; Ross, M.; Kim, S.; Tsai, F.-F.; Li, Q.-F.; Jeffry, J.; Kim, J.-Y.; Loh, H.H.; et al. Unidirectional Cross-Activation of GRPR by MOR1D Uncouples Itch and Analgesia Induced by Opioids. Cell 2011, 147, 447–458. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Dong, X. Itch Mechanisms and Circuits. Annu. Rev. Biophys. 2014, 43, 331–355. [Google Scholar] [CrossRef] [Green Version]
- Davidson, S.; Giesler, G.J. The multiple pathways for itch and their interactions with pain. Trends Neurosci. 2010, 33, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef]
- De La Fuente, M.; Noble, D.N.; Verma, S.; Nieman, M.T. Mapping Human Protease-activated Receptor 4 (PAR4) Homodimer Interface to Transmembrane Helix 4. J. Boil. Chem. 2012, 287, 10414–10423. [Google Scholar] [CrossRef] [Green Version]
- Arachiche, A.; Mumaw, M.M.; De La Fuente, M.; Nieman, M.T. Protease-activated Receptor 1 (PAR1) and PAR4 Heterodimers Are Required for PAR1-enhanced Cleavage of PAR4 by α-Thrombin. J. Boil. Chem. 2013, 288, 32553–32562. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Hu, H. TRP Channels as Drug Targets to Relieve Itch. Pharmaceutics 2018, 11, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-H.; Park, C.-K.; Chen, G.; Han, Q.; Xie, R.-G.; Liu, T.; Ji, R.-R.; Lee, S.-Y. RETRACTED: A Monoclonal Antibody that Targets a NaV1.7 Channel Voltage Sensor for Pain and Itch Relief. Cell 2014, 157, 1393–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, M.; Ganellin, C.R. Histamine and its receptors. Br. J. Pharmacol. 2006, 147, S127–S135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panula, P.; Chazot, P.L.; Cowart, M.; Gutzmer, R.; Leurs, R.; Liu, W.L.S.; Stark, H.; Thurmond, R.L.; Haas, H.L. International Union of Basic and Clinical Pharmacology. XCVIII. Histamine Receptors. Pharmacol. Rev. 2015, 67, 601–655. [Google Scholar] [CrossRef] [Green Version]
- Timmerman, H.; Leurs, R.; Van Der Goot, H. Histamine Receptors and Their Ligands: Mechanisms and Applications. In Reference Module in Neuroscience and Biobehavioral Psychology; Elsevier BV: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Canonica, G.; Blaiss, M. Antihistaminic, Anti-Inflammatory, and Antiallergic Properties of the Nonsedating Second-Generation Antihistamine Desloratadine: A Review of the Evidence. World Allergy Organ. J. 2011, 4, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passani, B.; Lin, J.-S.; Hancock, A.; Crochet, S.; Blandina, P. The histamine H3 receptor as a novel therapeutic target for cognitive and sleep disorders. Trends Pharmacol. Sci. 2004, 25, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Imamachi, N.; Park, G.H.; Lee, H.; Anderson, D.J.; Simon, M.I.; Basbaum, A.I.; Han, S.-K. TRPV1-expressing primary afferents generate behavioral responses to pruritogens via multiple mechanisms. Proc. Natl. Acad. Sci. USA 2009, 106, 11330–11335. [Google Scholar] [CrossRef] [Green Version]
- Pándy-Szekeres, G.; Munk, C.; Tsonkov, T.; Mordalski, S.; Harpsoe, K.; Hauser, A.S.; Bojarski, A.J.; Gloriam, D.E. GPCRdb in 2018: Adding GPCR structure models and ligands. Nucleic Acids Res. 2018, 46, D440–D446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denham, K.J.; Boutsiouki, P.; Clough, G.F.; Church, M.K. Comparison of the effects of desloratadine and levocetirizine on histamine-induced wheal, flare and itch in human skin. Inflamm. Res. 2003, 52, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Tanizaki, H.; Ikoma, A.; Fukuoka, M.; Miyachi, Y.; Kabashima, K. Effects of Bepotastine and Fexofenadine on Histamine-Induced Flare, Wheal and Itch. Int. Arch. Allergy Immunol. 2012, 158, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.G.; Greaves, M.W. Sensory responses of human skin to synthetic histamine analogues and histamine. Br. J. Clin. Pharmacol. 1980, 9, 461–465. [Google Scholar] [PubMed]
- Ohsawa, Y.; Hirasawa, N. The Role of Histamine H1 and H4 Receptors in Atopic Dermatitis: From Basic Research to Clinical Study. Allergol. Int. 2014, 63, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.K.; McQueen, D.S.; Rees, J.L. Involvement of histamine H4and H1receptors in scratching induced by histamine receptor agonists in BalbC mice. Br. J. Pharmacol. 2004, 142, 374–380. [Google Scholar] [CrossRef] [Green Version]
- Roßbach, K.; Nassenstein, C.; Gschwandtner, M.; Schnell, D.; Sander, K.; Seifert, R.; Stark, H.; Kietzmann, M.; Bäumer, W.; Vlad, G.; et al. Histamine H1, H3 and H4 receptors are involved in pruritus. Neurosciences. 2011, 190, 89–102. [Google Scholar] [CrossRef]
- 5Huang, H.; Li, Y.; Liang, J.; Finkelman, F.D. Molecular Regulation of Histamine Synthesis. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Simone, N.A.; Ngeow, J.Y.F.; Whitehouse, J.; Becerra-Cabal, L.; Putterman, G.J.; LaMotte, R.H. The Magnitude and Duration of Itch Produced by Intracutaneous Injections of Histamine. Somatosens. Res. 1987, 5, 81–92. [Google Scholar] [CrossRef]
- Horsmanheimo, L.; Harvima, I.T.; Harvima, R.J.; Brummer-Korvenkontio, H.; Francois, G.; Reunala, T. Histamine and leukotriene C4 release in cutaneous mosquito-bite reactions. J. Allergy Clin. Immunol. 1996, 98, 408–411. [Google Scholar] [CrossRef]
- Thurmond, R.L.; Kazerouni, K.; Chaplan, S.; Greenspan, A.; Carstens, E.; Akiyama, T. Peripheral Neuronal Mechanism of Itch. Advances in Vagal Afferent Neurobiology 2014, 20143036, 143–192. [Google Scholar]
- Hoyer, D.; Clarke, D.E.; Fozard, J.R.; Hartig, P.R.; Martin, G.R.; Mylecharane, E.J.; Saxena, P.R.; Humphrey, P.P. International Union of Pharmacology classification of receptors for 5-hydroxytryptamine (Serotonin). Pharmacol. Rev. 1994, 46, 157–203. [Google Scholar] [PubMed]
- Hoyer, D. Serotonin receptors nomenclature. In The Serotonin System; Tricklebank, M.D., Daly, E., Eds.; Academic Press: Cambridge, MA, USA, 2019; Chapter Four; pp. 63–93. [Google Scholar]
- Nichols, D.E.; Nichols, C.D. Serotonin Receptors. Chem. Rev. 2008, 108, 1614–1641. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.; Gray, J.; Roth, B.L. The Expanded Biology of Serotonin. Annu. Rev. Med. 2009, 60, 355–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masson, J.; Emerit, M.B.; Hamon, M.; Darmon, M. Serotonergic signaling: Multiple effectors and pleiotropic effects. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2012, 1, 685–713. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Nagasawa, T.; Satoh, M.; Kuraishi, Y. Itch-associated response induced by intradermal serotonin through 5-HT2 receptors in mice. Neurosci. Res. 1999, 35, 77–83. [Google Scholar] [CrossRef]
- Kim, D.K.; Kim, H.J.; Kim, H.; Koh, J.Y.; Kim, K.M.; Noh, M.; Kim, J.J.; Lee, C.H. Involvement of serotonin receptors 5-HT1 and 5-HT2 in 12(S)-HPETE-induced scratching in mice. Eur. J. Pharmacol. 2008, 579, 390–394. [Google Scholar] [CrossRef]
- Morita, T.; McClain, S.P.; Batia, L.M.; Pellegrino, M.; Wilson, S.R.; Kienzler, M.A.; Lyman, K.; Olsen, A.S.B.; Wong, J.F.; Stucky, C.L.; et al. HTR7 Mediates Serotonergic Acute and Chronic Itch. Neuron 2015, 87, 124–138. [Google Scholar] [CrossRef] [Green Version]
- Nojima, H.; Carstens, E. 5-Hydroxytryptamine (5-HT)2 Receptor Involvement in Acute 5-HT-Evoked Scratching but Not in Allergic Pruritus Induced by Dinitrofluorobenzene in Rats. J. Pharmacol. Exp. Ther. 2003, 306, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Wang, X.-L.; Huang, Y.; Chen, L.-H.; Cheng, R.-X.; Zhou, F.-M.; Guo, R.; Li, J.-C.; Liu, T. Peripheral and spinal 5-HT receptors participate in cholestatic itch and antinociception induced by bile duct ligation in rats. Sci. Rep. 2016, 6, 36286. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, T.; Ivanov, M.; Nagamine, M.; Davoodi, A.; Carstens, M.I.; Ikoma, A.; Cevikbas, F.; Kempkes, C.; Buddenkotte, J.; Steinhoff, M.; et al. Involvement of TRPV4 in Serotonin-Evoked Scratching. J. Investig. Dermatol. 2016, 136, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Ständer, S.; Böckenholt, B.; Schürmeyer-Horst, F.; Weishaupt, C.; Heuft, G.; Schneider, T. Treatment of Chronic Pruritus with the Selective Serotonin Re-uptake Inhibitors Paroxetine and Fluvoxamine: Results of an Open-labelled, Two-arm Proof-of-concept Study. Acta Derm. Venereol. 2009, 89, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Kim, K. Neuroimmunological Mechanism of Pruritus in Atopic Dermatitis Focused on the Role of Serotonin. Biomol. Ther. 2012, 20, 506–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonne-Rahm, S.; Rickberg, H.; Elnour, H.; Marin, P.; Azmitia, E.C.; Nordlind, K. Neuroimmune mechanisms in patients with atopic dermatitis during chronic stress. J. Eur. Acad. Dermatol. Venereol. 2007, 22, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.-Q.; Liu, X.-Y.; Jeffry, J.; Karunarathne, A.; Li, J.-L.; Munanairi, A.; Zhou, X.-Y.; Li, H.; Sun, Y.-G.; Wan, L.; et al. Descending control of itch transmission by the serotonergic system via 5-HT1A-facilitated GRP-GRPR signaling. Neurology 2014, 84, 821–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapport, M.M.; Green, A.A.; Page, I.H. Serum vasoconstrictor, serotonin; isolation and characterization. J. Boil. Chem. 1948, 176, 1243–1251. [Google Scholar]
- Walther, D.J.; Peter, J.-U.; Bashammakh, S.; Hörtnagl, H.; Voits, M.; Fink, H.; Bader, M. Synthesis of Serotonin by a Second Tryptophan Hydroxylase Isoform. Science 2003, 299, 76. [Google Scholar] [CrossRef]
- Bardoni, R. Serotonergic Modulation of Nociceptive Circuits in Spinal Cord Dorsal Horn. Curr Neuropharmacol. 2019, 17, 1133–1145. [Google Scholar] [CrossRef]
- Reddy, V.B.; Iuga, A.O.; Shimada, S.G.; LaMotte, R.H.; Lerner, E.A. Cowhage-evoked itch is mediated by a novel cysteine protease: A ligand of protease-activated receptors. J. Neurosci. 2008, 28, 4331–4335. [Google Scholar] [CrossRef] [Green Version]
- Nieman, M.T.; Schmaier, A.H. Interaction of Thrombin with PAR1 and PAR4 at the Thrombin Cleavage Site. Biochemicals 2007, 46, 8603–8610. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Covic, L.; Kuliopulos, A. Protease-Activated Receptors. In Platelets; Elsevier BV: Amsterdam, The Netherlands, 2013; pp. 249–259. [Google Scholar]
- Heuberger, D.M.; Schuepbach, R.A. Protease-activated receptors (PARs): Mechanisms of action and potential therapeutic modulators in PAR-driven inflammatory diseases. Thromb. J. 2019, 17, 4. [Google Scholar] [CrossRef] [Green Version]
- Nieman, M.T. Protease-activated receptors in hemostasis. Blood 2016, 128, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiyama, T.; Lerner, E.A.; Carstens, E. Protease-activated receptors and itch. Handb. Exp. Pharmacol. 2015, 226, 219–235. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.N.; Ramachandran, R.; Yau, M.-K.; Suen, J.; Fairlie, D.P.; Hollenberg, M.D.; Hooper, J.D. Structure, function and pathophysiology of protease activated receptors. Pharmacol. Ther. 2011, 130, 248–282. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.B.; Shimada, S.G.; Sikand, P.; LaMotte, R.H.; Lerner, E.A. Cathepsin S Elicits Itch and Signals via Protease-Activated Receptors. J. Investig. Dermatol. 2010, 130, 1468–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soh, U.J.; Dores, M.; Chen, B.; Trejo, J. Signal transduction by protease-activated receptors. Br. J. Pharmacol. 2010, 160, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.; Bae, K.B.; Kim, M.O.; Yu, D.H.; Kim, H.J.; Yuh, H.S.; Ji, Y.R.; Park, S.J.; Kim, S.; Son, K.-H.; et al. Overexpression of Cathepsin S Induces Chronic Atopic Dermatitis in Mice. J. Investig. Dermatol. 2012, 132, 1169–1176. [Google Scholar] [CrossRef] [Green Version]
- Davidson, S.; Zhang, X.; Yoon, C.H.; Khasabov, S.G.; Simone, N.A.; Giesler, G.J. The itch-producing agents histamine and cowhage activate separate populations of primate spinothalamic tract neurons. J. Neurosci. 2007, 27, 10007–10014. [Google Scholar] [CrossRef]
- Reddy, V.; Lerner, E.A. Plant cysteine proteases that evoke itch activate protease-activated receptors. Br. J. Dermatol. 2010, 163, 532–535. [Google Scholar] [CrossRef]
- Barry, G.D.; Le, G.T.; Fairlie, D.P. Agonists and antagonists of protease activated receptors (PARs). Curr. Med. Chem. 2006, 13, 243–265. [Google Scholar] [CrossRef]
- Fostini, A.; Golpanian, R.; Rosen, J.; Xue, R.-D.; Yosipovitch, G. Beat the bite: Pathophysiology and management of itch in mosquito bites. Itch 2019, 4, 1. [Google Scholar] [CrossRef]
- Lin, H.; Liu, A.P.; Smith, T.; Trejo, J. Cofactoring and dimerization of proteinase-activated receptors. Pharmacol. Rev. 2013, 65, 1198–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Fuente, M.; Arachiche, A.; Nieman, M.T. Thrombin Modulates PAR1-PAR4 Heterodimers. Blood 2011, 118, 2203. [Google Scholar] [CrossRef]
- Han, X.; Hofmann, L.; De La Fuente, M.; Alexander, N.; Palczewski, K.; INVENT INVENT Consortium; Nieman, M.T. PAR4 activation involves extracellular loop-3 and transmembrane residue Thr153. Blood 2020. [Google Scholar] [CrossRef]
- Zhao, P.; Metcalf, M.; Bunnett, N.W. Biased Signaling of Protease-Activated Receptors. Front. Endocrinol. 2014, 5, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Munanairi, A.; Liu, X.-Y.; Zhang, J.; Hu, L.; Hu, M.; Bu, D.; Liu, L.; Xie, Z.; Kim, B.S.; et al. PAR2 Mediates Itch via TRPV3 Signaling in Keratinocytes. J. Investig. Dermatol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ständer, S.; Luger, T.A. NK-1 Antagonists and Itch. Drug Delivery 2015, 226, 237–255. [Google Scholar] [CrossRef]
- Majkowska-Pilip, A.; Halik, P.; Gniazdowska, E. The Significance of NK1 Receptor Ligands and Their Application in Targeted Radionuclide Tumour Therapy. Pharm. 2019, 11, 443. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.-T.; Venkatachalam, K.; Li, H.-S.; Montell, C.; Kurosaki, T.; Patterson, R.L.; Gill, D.L. Assessment of the Role of the Inositol 1,4,5-Trisphosphate Receptor in the Activation of Transient Receptor Potential Channels and Store-operated Ca2+ Entry Channels. J. Boil. Chem. 2001, 276, 18888–18896. [Google Scholar] [CrossRef] [Green Version]
- Douglas, S.D.; Leeman, S.E. Neurokinin-1 receptor: Functional significance in the immune system in reference to selected infections and inflammation. Ann. N. Y. Acad. Sci. 2010, 1217, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Amatya, B.; Elnour, H.; Holst, M.; Theodorsson, E.; Nordlind, K. Expression of tachykinins and their receptors in plaque psoriasis with pruritus. Br. J. Dermatol. 2011, 164, 1023–1029. [Google Scholar] [CrossRef]
- Ständer, S.; Siepmann, D.; Herrgott, I.; Sunderkötter, C.; Luger, T.A. Targeting the Neurokinin Receptor 1 with Aprepitant: A Novel Antipruritic Strategy. PLoS ONE 2010, 5, e10968. [Google Scholar] [CrossRef]
- Ständer, S.; Yosipovitch, G. Substance P and neurokinin 1 receptor are new targets for the treatment of chronic pruritus. Br. J. Dermatol. 2019, 181, 932–938. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Zeng, S.; Wang, X.; Babazada, H.; Li, Z.; Liu, R.; Yu, W. Neurokinin 1 and opioid receptors: Relationships and interactions in nervous system. Transl. Perioper. Pain Med. 2016, 3, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Euler, U.S.V.; Gaddum, J.H. An unidentified depressor substance in certain tissue extracts. J. Physiol. 1931, 72, 74–87. [Google Scholar] [CrossRef]
- Studer, R.O.; Trzeciak, A.; Lergier, W. Isolierung und Aminosäuresequenz von Substanz P aus Pferdedarm. Helv. Chim. Acta 1973, 56, 860–866. [Google Scholar] [CrossRef]
- Severini, C. The Tachykinin Peptide Family. Pharmacol. Rev. 2002, 54, 285–322. [Google Scholar] [CrossRef]
- Haas, S.; Capellino, S.; Phan, N.Q.; Böhm, M.; Luger, T.A.; Straub, R.H.; Ständer, S. Low density of sympathetic nerve fibers relative to substance P-positive nerve fibers in lesional skin of chronic pruritus and prurigo nodularis. J. Dermatol. Sci. 2010, 58, 193–197. [Google Scholar] [CrossRef]
- Azimi, E.; Reddy, V.B.; Shade, K.-T.C.; Anthony, R.M.; Talbot, S.; Pereira, P.J.S.; Lerner, E.A. Dual action of neurokinin-1 antagonists on Mas-related GPCRs. JCI Insight 2016, 1, e89362. [Google Scholar] [CrossRef]
- Katsube, N.; Maruyama, M.; Andoh, T.; Kuraishi, Y. Involvement of Leukotriene B4 in Substance P-Induced Itch-Associated Response in Mice. J. Investig. Dermatol. 2001, 117, 1621–1626. [Google Scholar] [CrossRef] [Green Version]
- Hosogi, M.; Schmelz, M.; Miyachi, Y.; Ikoma, A. Bradykinin is a potent pruritogen in atopic dermatitis: A switch from pain to itch. Pain 2006, 126, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Raimondi, F.; Kadji, F.M.N.; Singh, G.; Kishi, T.; Uwamizu, A.; Ono, Y.; Shinjo, Y.; Ishida, S.; Arang, N.; et al. Illuminating G-Protein-Coupling Selectivity of GPCRs. Cell 2019, 177, 1933–1947.e25. [Google Scholar] [CrossRef]
- Calixto, J.B.; Cabrini, D.A.; Ferreira, J.; Campos, M.M. Kinins in pain and inflammation. Pain 2000, 87, 1–5. [Google Scholar] [CrossRef]
- Liang, J.; He, Y.; Ji, W. Bradykinin-evoked scratching responses in complete Freund’s adjuvant-inflamed skin through activation of B1 receptor. Exp. Biol. Med. 2012, 237, 318–326. [Google Scholar] [CrossRef]
- Hayashi, I.; Majima, M. Reduction of sodium deoxycholic acid-induced scratching behaviour by bradykinin B2receptor antagonists. Br. J. Pharmacol. 1999, 126, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Lang, E.; Novák, A.; Reeh, P.W.; Handwerker, H.O. Chemosensitivity of fine afferents from rat skin in vitro. J. Neurophysiol. 1990, 63, 887–901. [Google Scholar] [CrossRef] [PubMed]
- Koppert, W.; Martus, P.; Reeh, P.W. Interactions of histamine and bradykinin on polymodal C-fibres in isolated rat skin. Eur. J. Pain 2001, 5, 97–106. [Google Scholar] [CrossRef]
- Averbeck, B.; Reeh, P.W. Interactions of inflammatory mediators stimulating release of calcitonin gene-related peptide, substance P and prostaglandin E2 from isolated rat skin. Neuropharmacol. 2001, 40, 416–423. [Google Scholar] [CrossRef]
- Golias, C.; Charalabopoulos, A.; Stagikas, D.; Charalabopoulos, K.; Batistatou, A. The kinin system—Bradykinin: Biological effects and clinical implications. Multiple role of the kinin system—Bradykinin. Hippokratia 2007, 11, 124–128. [Google Scholar]
- Tetta, C.; Maffei, S.; Cisterna, B.; Fonsato, V.; Triolo, G.; Paolo, G.; Camussi, G.; Chiara, M.; Gatti, E. The Evolution of Biocompatibility: From Microinflammation to Microvesiscles. In Progress in Hemodialysis—From Emergent Biotechnology to Clinical Practice; Intech Open: London, UK, 2011. [Google Scholar]
- Liu, Y.; Liu, J.; Li, M.; Dai, S.; Liang, J.; Ji, W. The effect of kinin B1 receptor on chronic itching sensitization. Mol. Pain 2015, 11, 70–75. [Google Scholar] [CrossRef] [Green Version]
- Cormia, F.E.; Dougherty, J.W. Proteolytic activity in development of pain and itching. Cutaneous reactions to bradykinin and kallikrein. J. Investig. Dermatol. 1960, 35, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Poyner, D.R.; Sexton, P.M.; Marshall, I.; Smith, D.M.; Quirion, R.; Born, W.; Muff, R.; Fischer, J.A.; Foord, S.M. International Union of Pharmacology. XXXII. The mammalian calcitonin gene-related peptides, adrenomedullin, amylin, and calcitonin receptors. Pharmacol. Rev. 2002, 54, 233–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booe, J.M.; Warner, M.L.; Roehrkasse, A.M.; Hay, D.L.; Pioszak, A.A. Probing the Mechanism of Receptor Activity–Modifying Protein Modulation of GPCR Ligand Selectivity through Rational Design of Potent Adrenomedullin and Calcitonin Gene-Related Peptide Antagonists. Mol. Pharmacol. 2018, 93, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Kuwasako, K.; Cao, Y.-N.; Chu, C.-P.; Iwatsubo, S.; Eto, T.; Kitamura, K. Functions of the Cytoplasmic Tails of the Human Receptor Activity-modifying Protein Components of Calcitonin Gene-related Peptide and Adrenomedullin Receptors. J. Boil. Chem. 2006, 281, 7205–7213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Southan, C.; Sharman, J.L.; Benson, H.E.; Faccenda, E.; Pawson, A.J.; Alexander, S.P.; Buneman, O.P.; Davenport, A.P.; McGrath, J.C.; Peters, J.A.; et al. The IUPHAR/BPS Guide to PHARMACOLOGY in 2016: Towards curated quantitative interactions between 1300 protein targets and 6000 ligands. Nucleic Acids Res. 2015, 44, D1054–D1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottrell, G.S. CGRP Receptor Signalling Pathways. Drug Delivery 2018, 255, 37–64. [Google Scholar] [CrossRef]
- Rogoz, K.; Andersen, H.H.; Lagerström, M.C.; Kullander, K. Multimodal Use of Calcitonin Gene-Related Peptide and Substance P in Itch and Acute Pain Uncovered by the Elimination of Vesicular Glutamate Transporter 2 from Transient Receptor Potential Cation Channel Subfamily V Member 1 Neurons. J. Neurosci. 2014, 34, 14055–14068. [Google Scholar] [CrossRef] [Green Version]
- McCoy, E.S.; Taylor-Blake, B.; Street, S.E.; Pribisko, A.L.; Zheng, J.; Zylka, M.J. Peptidergic CGRPα primary sensory neurons encode heat and itch and tonically suppress sensitivity to cold. Neuron 2013, 78, 138–151. [Google Scholar] [CrossRef] [Green Version]
- Russell, F.A.; King, R.; Smillie, S.J.; Kodji, X.; Brain, S.D. Calcitonin gene-related peptide: Physiology and pathophysiology. Physiol. Rev. 2014, 94, 1099–1142. [Google Scholar] [CrossRef] [Green Version]
- Kee, Z.; Kodji, X.; Brain, S.D. The Role of Calcitonin Gene Related Peptide (CGRP) in Neurogenic Vasodilation and Its Cardioprotective Effects. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Amara, S.; Jonas, V.; Rosenfeld, M.G.; Ong, E.S.; Evans, R.M. Alternative RNA processing in calcitonin gene expression generates mRNAs encoding different polypeptide products. Nature 1982, 298, 240–244. [Google Scholar] [CrossRef]
- Schlereth, T.; Schukraft, J.; Krämer-Best, H.H.; Geber, C.; Ackermann, T.; Birklein, F. Interaction of calcitonin gene related peptide (CGRP) and substance P (SP) in human skin. Neuropeptides 2016, 59, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Vaalasti, A.; Suomalainen, H.; Rechardt, L. Calcitonin gene-related peptide immunoreactivity in prurigo nodularis: A comparative study with neurodermatitis circumscripta. Br. J. Dermatol. 1989, 120, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, D.M.; Rasmussen, S.G.F.; Kobilka, B.K. The structure and function of G-protein-coupled receptors. Nature 2009, 459, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Jackson, T.R.; Blair, L.A.C.; Marshall, J.; Goedert, M.; Hanley, M.R. The mas oncogene encodes an angiotensin receptor. Nature 1988, 335, 437–440. [Google Scholar] [CrossRef]
- Dong, X.; Han, S.-K.; Zylka, M.J.; Simon, M.I.; Anderson, D.J. A Diverse Family of GPCRs Expressed in Specific Subsets of Nociceptive Sensory Neurons. Cell 2001, 106, 619–632. [Google Scholar] [CrossRef] [Green Version]
- Bader, M.; Alenina, N.; Andrade-Navarro, M.A.; Santos, R.A. Mas and Its Related G protein-coupled Receptors, Mrgprs. Pharmacol. Rev. 2014, 66, 1080–1105. [Google Scholar] [CrossRef]
- Zhang, L.; Taylor, N.; Xie, Y.; Ford, R.; Johnson, J.; Paulsen, J.E.; Bates, B. Cloning and expression of MRG receptors in macaque, mouse, and human. Mol. Brain Res. 2005, 133, 187–197. [Google Scholar] [CrossRef]
- Han, L.; Ma, C.; Liu, Q.; Weng, H.-J.; Cui, Y.; Tang, Z.; Kim, Y.; Nie, H.; Qu, L.; Patel, K.N.; et al. A subpopulation of nociceptors specifically linked to itch. Nat. Neurosci. 2012, 16, 174–182. [Google Scholar] [CrossRef]
- Han, S.-K.; Dong, X.; Hwang, J.-I.; Zylka, M.J.; Anderson, D.J.; Simon, M.I. Orphan G protein-coupled receptors MrgA1 and MrgC11 are distinctively activated by RF-amide-related peptides through the G q/11 pathway. Proc. Natl. Acad. Sci USA 2002, 99, 14740–14745. [Google Scholar] [CrossRef] [Green Version]
- Mc Lembo, P.; Grazzini, E.; Groblewski, T.; O’Donnell, D.; Roy, M.-O.; Zhang, J.; Hoffert, C.; Cao, J.; Schmidt, R.; Pelletier, M.; et al. Proenkephalin A gene products activate a new family of sensory neuron–specific GPCRs. Nat. Neurosci. 2002, 5, 201–209. [Google Scholar] [CrossRef]
- Prchalová, E.; Hin, N.; Thomas, A.G.; Veeravalli, V.; Ng, J.; Alt, J.; Rais, R.; Rojas, C.; Li, Z.; Hihara, H.; et al. Discovery of Benzamidine- and 1-Aminoisoquinoline-Based Human MAS-Related G-Protein-Coupled Receptor X1 (MRGPRX1) Agonists. J. Med. Chem. 2019, 62, 8631–8641. [Google Scholar] [CrossRef]
- Wilson, S.R.; Gerhold, K.A.; Bifolck-Fisher, A.; Liu, Q.; Patel, K.N.; Dong, X.; Bautista, D.M. TRPA1 is required for histamine-independent, Mas-related G protein-coupled receptor-mediated itch. Nat. Neurosci. 2011, 14, 595–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghahowa, S.E.; Obianwu, H.O.; Isah, A.O.; Arhewoh, I.M. Chloroquine-induced Pruritus. Indian J. Pharm. Sci. 2010, 72, 283–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osifo, N.G. The antipruritic effects of chlorpheniramine, cyproheptadine and sulphapyridine monitored with limb activity meters on chloroquine induced pruritus among patients with malaria. Afr. J. Med. Med. Sci. 1995, 24, 67–73. [Google Scholar] [PubMed]
- Abila, B.; Ezeamuzie, I.C.; Igbigbi, P.S.; Ambakederemo, A.W.; Asomugha, L. Effects of two antihistamines on chloroquine and histamine induced weal and flare in healthy African volunteers. Afr. J. Med. Med. Sci. 1994, 23, 139–142. [Google Scholar]
- Sanjel, B.; Maeng, H.J.; Shim, W.-S. BAM8-22 and its receptor MRGPRX1 may attribute to cholestatic pruritus. Sci. Rep. 2019, 9, 10888–10913. [Google Scholar] [CrossRef] [Green Version]
- Swain, M.G.; MacArthur, L.; Vergalla, J.; Jones, E.A. Adrenal secretion of BAM-22P, a potent opioid peptide, is enhanced in rats with acute cholestasis. Am. J. Physiol. Liver Physiol. 1994, 266, G201–G205. [Google Scholar] [CrossRef]
- Chen, H.; Ikeda, S.R. Modulation of Ion Channels and Synaptic Transmission by a Human Sensory Neuron-Specific G-Protein-Coupled Receptor, SNSR4/mrgX1, Heterologously Expressed in Cultured Rat Neurons. J. Neurosci. 2004, 24, 5044–5053. [Google Scholar] [CrossRef]
- Sikand, P.; Dong, X.; LaMotte, R.H. BAM8–22 Peptide Produces Itch and Nociceptive Sensations in Humans Independent of Histamine Release. J. Neurosc. 2011, 31, 7563–7567. [Google Scholar] [CrossRef]
- Liu, Q.; Weng, H.-J.; Patel, K.N.; Tang, Z.; Bai, H.; Steinhoff, M.; Dong, X. The Distinct Roles of Two GPCRs, MrgprC11 and PAR2, in Itch and Hyperalgesia. Sci. Signal. 2011, 4, ra45. [Google Scholar] [CrossRef] [Green Version]
- Toda, A.; Yokomizo, T.; Shimizu, T. Leukotriene B4 receptors. Prostaglandins Other Lipid Mediat. 2002, 68, 575–585. [Google Scholar] [CrossRef]
- Andoh, T.; Saito, A.; Kuraishi, Y. Leukotriene B4 Mediates Sphingosylphosphorylcholine-Induced Itch-Associated Responses in Mouse Skin. J. Investig. Derma. 2009, 129, 2854–2860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andoh, T.; Haza, S.; Saito, A.; Kuraishi, Y. Involvement of leukotriene B4 in spontaneous itch-related behaviour in NC mice with atopic dermatitis-like skin lesions. Exp. Dermatol. 2011, 20, 894–898. [Google Scholar] [CrossRef]
- Andoh, T.; Kuraishi, Y. Intradermal leukotriene B4, but not prostaglandin E2, induces itch-associated responses in mice. Eur. J. Pharmacol. 1998, 353, 93–96. [Google Scholar] [CrossRef]
- Tsukumo, Y.; Harada, D.; Manabe, H. Pharmacological characterization of itch-associated response induced by repeated application of oxazolone in mice. J. Pharmacol. Sci. 2010, 113, 255–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camp, R.; Jones, R.R.; Brain, S.D.; Woollard, P.; Greaves, M. Production of Intraepidermal Microabscesses by Topical Application of Leukotriene B4. J. Investig. Dermatol. 1984, 82, 202–204. [Google Scholar] [CrossRef] [Green Version]
- Soter, N.; Lewis, R.A.; Corey, E.J.; Austen, K.F. Local Effects of Synthetic Leukotrienes (LTC4, LTD4, and LTB4) in Human Skin. J. Investig. Dermatol. 1983, 80, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Gijón, M.A.; Zarini, S.; Murphy, R.C. Biosynthesis of eicosanoids and transcellular metabolism of leukotrienes in murine bone marrow cells. J. Lipid Res. 2006, 48, 716–725. [Google Scholar] [CrossRef] [Green Version]
- Harizi, H.; Corcuff, J.-B.; Gualde, N. Arachidonic-acid-derived eicosanoids: Roles in biology and immunopathology. Trends Mol. Med. 2008, 14, 461–469. [Google Scholar] [CrossRef]
- Yokomizo, T.; Izumi, T.; Shimizu, T. Leukotriene B4: Metabolism and Signal Transduction. Arch. Biochem. Biophys. 2001, 385, 231–241. [Google Scholar] [CrossRef]
- Yost, C.C.; Weyrich, A.S.; Zimmerman, G.A. The platelet activating factor (PAF) signaling cascade in systemic inflammatory responses. Biochimicals 2010, 92, 692–697. [Google Scholar] [CrossRef] [Green Version]
- Fjellner, B.; Hägermark, O. Experimental pruritus evoked by platelet activating factor (PAF-acether) in human skin. Acta Derm. Venereol. 1985, 65, 409–412. [Google Scholar]
- Shigenobu, K.; Mori, T.; Kamata, K.; Kasuya, Y. Platelet-activating factor: Lack of direct action on guinea pig myocardium and possible transmitter release from cardiac sympathetic nerve endings at high concentrations. Can. J. Physiol. Pharmacol. 1989, 67, 669–674. [Google Scholar] [CrossRef]
- Petersen, L.J.; Church, M.K.; Skov, P.S. Platelet-activating factor induces histamine release from human skin mast cells in vivo, which is reduced by local nerve blockade. J. Allergy Clin. Immunol. 1997, 99, 640–647. [Google Scholar] [CrossRef]
- Prescott, S.M.; Zimmerman, G.A.; Stafforini, D.M.; McIntyre, T.M. Platelet-Activating Factor and Related Lipid Mediators. Annu. Rev. Biochem. 2000, 69, 419–445. [Google Scholar] [CrossRef]
- Inflammation: Basic Principles and Clinical Correlates. Ann. Int. Med. 1988, 109, 519. [CrossRef] [Green Version]
- Abeck, D.; Andersson, T.; Grosshans, E.; Jablonska, S.; Kragballe, K.; Vahlquist, A.; Schmidt, T.; Dupuy, P.; Ring, J. Topical application of a platelet-activating factor (PAF) antagonist in atopic dermatitis. Acta Derm. Venereol. 1997, 77, 449–451. [Google Scholar]
- Greisenegger, E.K.; Zimprich, A.; Stingl, G.; Kopp, T.; Zimprich, F. Analysis of the prodynorphin promoter polymorphism in atopic dermatitis and disease-related pruritus. Clin. Exp. Dermatol. 2009, 34, 728–730. [Google Scholar] [CrossRef]
- Melo, H.; Basso, L.; Iftinca, M.; Macnaughton, W.K.; Hollenberg, M.D.; McKay, D.M.; Altier, C. Itch induced by peripheral mu opioid receptors is dependent on TRPV1-expressing neurons and alleviated by channel activation. Sci. Rep. 2018, 8, 15551. [Google Scholar] [CrossRef]
- Goodman, A.J.; Le Bourdonnec, B.; Dolle, R.E. Mu Opioid Receptor Antagonists: Recent Developments. ChemMedChem 2007, 2, 1552–1570. [Google Scholar] [CrossRef]
- Yuan, C.-S.; Foss, J.; O’Connor, M.; Osinski, J.; Roizen, M.F.; Moss, J. Efficacy of orally administered methylnaltrexone in decreasing subjective effects after intravenous morphine. Drug Alcohol Depend. 1998, 52, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Avidor-Reiss, T.; Nevo, I.; Saya, D.; Bayewitch, M.; Vogel, Z. Opiate-induced adenylyl cyclase superactivation is isozyme-specific. J. Biol. Chem. 1997, 272, 5040–5047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, A.; Szepietowski, J.C. Opioid-induced pruritus: An update. Clin. Exp. Dermatol. 2010, 35, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Taneda, K.; Tominaga, M.; Negi, O.; Tengara, S.; Kamo, A.; Ogawa, H.; Takamori, K. Evaluation of epidermal nerve density and opioid receptor levels in psoriatic itch. Br. J. Dermatol. 2011, 165, 277–284. [Google Scholar] [CrossRef]
- Tominaga, M.; Ogawa, H.; Takamori, K. Possible Roles of Epidermal Opioid Systems in Pruritus of Atopic Dermatitis. J. Investig. Dermatol. 2007, 127, 2228–2235. [Google Scholar] [CrossRef] [Green Version]
- Kupczyk, P.; Reich, A.; Holysz, M.; Gajda, M.; Wysokińska, E.; Kobuszewska, A.; Nevozhay, D.; Nowakowska, B.; Strzadała, L.; Jagodziński, P.P.; et al. Opioid Receptors in Psoriatic Skin: Relationship with Itch. Acta Derm. Venereol. 2017, 97, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Holzer, P. Non-analgesic effects of opioids: Management of opioid-induced constipation by peripheral opioid receptor antagonists: Prevention or withdrawal? Curr. Pharm. Des. 2012, 18, 6010–6020. [Google Scholar] [CrossRef]
- Phan, N.Q.; Siepmann, D.; Gralow, I.; Ständer, S. Adjuvant topical therapy with a cannabinoid receptor agonist in facial postherpetic neuralgia. J. der Dtsch. Dermatol. Ges. 2009, 8, 88–91. [Google Scholar] [CrossRef]
- Phan, N.; Lotts, T.; Antal, A.; Bernhard, J.; Ständer, S. Systemic Kappa Opioid Receptor Agonists in the Treatment of Chronic Pruritus: A Literature Review. Acta Derm. Venereol. 2012, 92, 555–560. [Google Scholar] [CrossRef]
- Day, R.; Lazure, C.; Basak, A.; Boudreault, A.; Limperis, P.; Dong, W.; Lindberg, I. Prodynorphin processing by proprotein convertase 2. Cleavage at single basic residues and enhanced processing in the presence of carboxypeptidase activity. J. Biol. Chem. 1998, 273, 829–836. [Google Scholar] [CrossRef] [Green Version]
- Kardon, A.P.; Polgár, E.; Hachisuka, J.; Snyder, L.M.; Cameron, D.; Savage, S.; Cai, X.; Karnup, S.; Fan, C.R.; Hemenway, G.M.; et al. Dynorphin acts as a neuromodulator to inhibit itch in the dorsal horn of the spinal cord. Neuron 2014, 82, 573–586. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, A. Opioid peptides endorphins in pituitary and brain. Science 1976, 193, 1081–1086. [Google Scholar] [CrossRef]
- Ishikawa, T.; Hashimoto, T.; Munetsugu, T.; Yokozeki, H.; Satoh, T. Increased β-endorphin and autotaxin in patients with prurigo. J. Cutan. Immunol. Allergy 2019, 2, 94–101. [Google Scholar] [CrossRef]
- Mackie, K. Cannabinoid Receptors: Where They are and What They do. J. Neuroendocr. 2008, 20, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef] [PubMed]
- Stander, S.; Reinhardt, H.W.; Luger, T.A. Topical cannabinoid agonists. An effective new possibility for treating chronic pruritus. Hautarzt 2006, 57, 801–807. [Google Scholar]
- Avila, C.; Massick, S.; Kaffenberger, B.H.; Kwatra, S.G.; Bechtel, M. Cannabinoids for the treatment of chronic pruritus: A review. J. Am. Acad. Dermatol. 2020, 82, 1205–1212. [Google Scholar] [CrossRef]
- Pogatzki-Zahn, E.; Marziniak, M.; Schneider, G.; Luger, T.A.; Ständer, S. Chronic pruritus: Targets, mechanisms and future therapies. Drug News Perspect. 2008, 21, 541–551. [Google Scholar] [PubMed]
- Dvorak, M.; Watkinson, A.; McGlone, F.; Rukwied, R. Histamine induced responses are attenuated by a cannabinoid receptor agonist in human skin. Inflamm. Res. 2003, 52, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Binzen, U.; Greffrath, W.; Hennessy, S.; Bausen, M.; Saaler-Reinhardt, S.; Treede, R.-D. Co-expression of the voltage-gated potassium channel Kv1.4 with transient receptor potential channels (TRPV1 and TRPV2) and the cannabinoid receptor CB1 in rat dorsal root ganglion neurons. Neurosciences 2006, 142, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Duo, L.; Hu, L.; Tian, N.; Cheng, G.; Wang, H.; Lin, Z.; Wang, Y.; Yang, Y. TRPV1 gain-of-function mutation impairs pain and itch sensations in mice. Mol. Pain 2018, 14. [Google Scholar] [CrossRef] [PubMed]
- Moiseenkova-Bell, V.Y.; Stanciu, L.A.; Serysheva, I.I.; Tobe, B.J.; Wensel, T.G. Structure of TRPV1 channel revealed by electron cryomicroscopy. Proc. Natl. Acad. Sci. USA 2008, 105, 7451–7455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, H.; Hiura, A. Four Possible Itching Pathways Related to the TRPV1 Channel, Histamine, PAR-2 and Serotonin. Malays. J. Med. Sci. 2013, 20, 5–12. [Google Scholar]
- Wang, J.; Ueda, N. Biology of endocannabinoid synthesis system. Prostaglandins Other Lipid Mediat. 2009, 89, 112–119. [Google Scholar] [CrossRef]
- Di Marzo, V. Endocannabinoids: Synthesis and degradation. Rev. Physiol. Biochem. Pharmacol. 2006, 160, 1–24. [Google Scholar] [CrossRef]
- Lu, H.-C.; Mackie, K. An Introduction to the Endogenous Cannabinoid System. Boil. Psychiatry 2015, 79, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Grotenhermen, F. Cannabinoids. Curr. Drug. Targets CNS Neurol. Disord. 2005, 4, 507–530. [Google Scholar] [CrossRef] [PubMed]
- Pacher, P.; Bátkai, S.; Kunos, G. The Endocannabinoid System as an Emerging Target of Pharmacotherapy. Pharmacol. Rev. 2006, 58, 389–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zygmunt, P.; Petersson, J.; Andersson, D.A.; Chuang, H.-H.; Sørgård, M.; Di Marzo, V.; Julius, D.; Högestätt, E.D. Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 1999, 400, 452–457. [Google Scholar] [CrossRef]
- Muller, C.; Morales, P.; Reggio, P.H. Cannabinoid Ligands Targeting TRP Channels. Front. Mol. Neurosci. 2019, 11, 487. [Google Scholar] [CrossRef]
- Carstens, E.; Akiyama, T. Itch: Mechanisms and Treatment; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2014. [Google Scholar]
- Asao, H. Interleukin-2. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Malek, T.R.; Castro, I. Interleukin-2 Receptor Signaling: At the Interface between Tolerance and Immunity. Immunity 2010, 33, 153–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, B.H.; Willerford, D.M. Biology of the Interleukin-2 Receptor. In Advances in Immunology; Elsevier BV: Berlin/Heidelberg, Germany, 1998; Volume 70, pp. 1–81. [Google Scholar]
- Gaffen, S.L. Signaling Domains of the Interleukin 2 Receptor. Cytokine 2001, 14, 63–77. [Google Scholar] [CrossRef]
- Azim, A.A.A.; Farag, A.S.; Hassan, D.A.E.-M.; Abdu, S.M.I.; Lashin, S.M.A.-E.; Abdelaziz, N.M. Role of Interleukin-2 in Uremic Pruritus Among Attendants of AL-Zahraa Hospital Dialysis Unit. Indian J. Dermatol. 2015, 60, 211. [Google Scholar] [PubMed]
- Darsow, U.; Scharein, E.; Bromm, B.; Ring, J. Skin testing of the pruritogenic activity of histamine and cytokines (interleukin-2 and tumour necrosis factor-α) at the dermal-epidermal junction. Br. J. Dermatol. 1997, 137, 415–417. [Google Scholar] [CrossRef] [PubMed]
- Boulay, J.-L.; Paul, W.E. Hematopoietin sub-family classification based on size, gene organization and sequence homology. Curr. Boil. 1993, 3, 573–581. [Google Scholar] [CrossRef]
- Théze, J. Interleukin 2. In Encyclopedia of Immunology; Elsevier BV: Amsterdam, The Netherlands, 1998; pp. 1436–1438. [Google Scholar]
- Luckheeram, R.V.; Zhou, R.; Verma, A.D.; Xia, B. CD4+T Cells: Differentiation and Functions. Clin. Dev. Immunol. 2012, 2012, 1–12. [Google Scholar] [CrossRef] [Green Version]
- IL-2. Available online: http://chemocare.com/chemotherapy/drug-info/il-2.aspx (accessed on 9 July 2020).
- Mueller, T.D.; Zhang, J.-L.; Sebald, W.; Duschl, A. Structure, binding, and antagonists in the IL-4/IL-13 receptor system. Biochimica et Biophysica Acta (BBA)—Mol. Cell. Res. 2002, 1592, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Heeb, L.E.M.; Egholm, C.; Boyman, O. Evolution and function of interleukin-4 receptor signaling in adaptive immunity and neutrophils. Genes Immun. 2020, 1–7. [Google Scholar] [CrossRef] [Green Version]
- LaPorte, S.L.; Juo, Z.S.; Vaclavikova, J.; Colf, L.A.; Qi, X.; Heller, N.M.; Keegan, A.D.; Garcia, K.C. Molecular and structural basis of cytokine receptor pleiotropy in the interleukin-4/13 system. Cell 2008, 132, 259–272. [Google Scholar] [CrossRef] [Green Version]
- Weidemann, T.; Höfinger, S.; Auer, M. Signaling of IL-4R, a Typical Class I Cytokine Receptor. In Handbook of Cell Signaling; Elsevier BV: Amsterdam, The Netherlands, 2010; pp. 323–328. [Google Scholar]
- Suzuki, A.; Leland, P.; Joshi, B.H.; Puri, R. Targeting of IL-4 and IL-13 receptors for cancer therapy. Cytokine 2015, 75, 79–88. [Google Scholar] [CrossRef]
- Junttila, I.S. Tuning the Cytokine Responses: An Update on Interleukin (IL)-4 and IL-13 Receptor Complexes. Front. Immunol. 2018, 9, 888. [Google Scholar] [CrossRef]
- Campion, M.; Smith, L.; Gatault, S.; Metais, C.; Buddenkotte, J.; Steinhoff, M. Interleukin-4 and interleukin-13 evoke scratching behaviour in mice. Exp. Dermatol. 2019, 28, 1501–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oetjen, L.K.; Mack, M.R.; Feng, J.; Whelan, T.M.; Niu, H.; Guo, C.J.; Chen, S.; Trier, A.M.; Xu, A.Z.; Tripathi, S.V.; et al. Sensory Neurons Co-opt Classical Immune Signaling Pathways to Mediate Chronic Itch. Cell 2017, 171, 217–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, S.; Humbert, M.; Barkans, J.; Corrigan, C.J.; Pfister, R.; Menz, G.; Larché, M.; Robinson, D.S.; Durham, S.R.; Kay, A.B. Expression of IL-4 and IL-5 mRNA and protein product by CD4+ and CD8+ T cells, eosinophils, and mast cells in bronchial biopsies obtained from atopic and nonatopic (intrinsic) asthmatics. J. Immunol. 1997, 158, 3539–3544. [Google Scholar] [PubMed]
- Beck, L.A.; Thaci, D.; Hamilton, J.D.; Graham, N.M.; Bieber, T.; Rocklin, R.; Ming, J.E.; Ren, H.; Kao, R.; Simpson, E.; et al. Dupilumab Treatment in Adults with Moderate-to-Severe Atopic Dermatitis. N. Engl. J. Med. 2014, 371, 130–139. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.L.; Bieber, T.; Guttman-Yassky, E.; Beck, L.A.; Blauvelt, A.; Cork, M.J.; Silverberg, J.I.; Deleuran, M.; Kataoka, Y.; Lacour, J.P.; et al. Two Phase 3 Trials of Dupilumab versus Placebo in Atopic Dermatitis. N. Engl. J. Med. 2016, 375, 2335–2348. [Google Scholar] [CrossRef]
- Thaçi, D.; Simpson, E.; Beck, L.; Bieber, T.; Blauvelt, A.; Papp, K.; Soong, W.; Worm, M.; Szepietowski, J.C.; Sofen, H.; et al. Efficacy and safety of dupilumab in adults with moderate-to-severe atopic dermatitis inadequately controlled by topical treatments: A randomised, placebo-controlled, dose-ranging phase 2b trial. Lancet 2016, 387, 40–52. [Google Scholar] [CrossRef]
- Lupardus, P.J.; Birnbaum, M.E.; Garcia, K.C. Molecular basis for shared cytokine recognition revealed in the structure of an unusually high affinity complex between IL-13 and IL-13Ralpha2. Structure 2010, 18, 332–342. [Google Scholar] [CrossRef] [Green Version]
- Hershey, G.K.K. IL-13 receptors and signaling pathways: An evolving web. J. Allergy Clin. Immunol. 2003, 111, 677–690. [Google Scholar] [CrossRef]
- Oh, M.-H.; Oh, S.Y.; Lu, J.; Lou, H.; Myers, A.C.; Zhu, Z.; Zheng, T. TRPA1-dependent pruritus in IL-13-induced chronic atopic dermatitis. J. Immunol. 2013, 191, 5371–5382. [Google Scholar] [CrossRef] [Green Version]
- Chandramohan, V.; Sampson, J.H.; Pastan, I.H.; Bigner, D.D. Chapter 10-Immunotoxin Therapy for Brain Tumors. In Translational Immunotherapy of Brain Tumors; Sampson, J.H., Ed.; Academic Press: San Diego, CA, USA, 2017; pp. 227–260. [Google Scholar]
- Minty, A.; Chalon, P.; Derocq, J.-M.; Dumont, X.; Guillemot, J.-C.; Kaghad, M.; Labit, C.; Leplatois, P.; Liauzun, P.; Miloux, B.; et al. lnterleukin-13 is a new human lymphokine regulating inflammatory and immune responses. Nature 1993, 362, 248–250. [Google Scholar] [CrossRef]
- Bao, K.; Reinhardt, R.L. The differential expression of IL-4 and IL-13 and its impact on type-2 immunity. Cytokine 2015, 75, 25–37. [Google Scholar] [CrossRef]
- Kelly-Welch, A.; Hanson, E.M.; Keegan, A.D. Interleukin-13 (IL-13) Pathway. Science’s STKE 2005, 2005, cm8. [Google Scholar] [CrossRef] [PubMed]
- Punnonen, J.; Carballido, J.M.; Aversa, G.; De Vries, J.E. Interleukin 13 and its Receptor. In Encyclopedia of Immunology; Elsevier BV: Amsterdam, The Netherlands, 1998; pp. 1489–1492. [Google Scholar]
- Zhang, Q.; Putheti, P.; Zhou, Q.; Liu, Q.; Gao, W. Structures and biological functions of IL-31 and IL-31 receptors. Cytokine Growth Factor Rev. 2008, 19, 347–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelissen, C.; Lüscher-Firzlaff, J.; Baron, J.M.; Lüscher, B. Signaling by IL-31 and functional consequences. Eur. J. Cell Boil. 2012, 91, 552–566. [Google Scholar] [CrossRef]
- Dillon, S.R.; Sprecher, C.; Hammond, A.; Bilsborough, J.; Rosenfeld-Franklin, M.; Presnell, S.R.; Haugen, H.S.; Maurer, M.; Harder, B.; Johnston, J.; et al. Interleukin 31, a cytokine produced by activated T cells, induces dermatitis in mice. Nat. Immunol. 2004, 5, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, C.; Marquardt, Y.; Czaja, K.; Wenzel, J.; Frank, J.; Lüscher-Firzlaff, J.; Lüscher, B.; Baron, J.M.; Wenzel, J. IL-31 regulates differentiation and filaggrin expression in human organotypic skin models. J. Allergy Clin. Immunol. 2012, 129, 426–433.e8. [Google Scholar] [CrossRef]
- Neis, M.; Peters, B.; Dreuw, A.; Wenzel, J.; Bieber, T.; Mauch, C.; Krieg, T.; Stanzel, S.; Heinrich, P.C.; Merk, H.F. Enhanced expression levels of IL-31 correlate with IL-4 and IL-13 in atopic and allergic contact dermatitis. J. Allergy Clin. Immunol. 2006, 118, 930–937. [Google Scholar] [CrossRef]
- Cevikbas, F.; Wang, X.; Akiyama, T.; Kempkes, C.; Savinko, T.; Antal, A.; Kukova, G.; Buhl, T.; Ikoma, A.; Buddenkotte, J.; et al. A sensory neuron-expressed IL-31 receptor mediates T helper cell-dependent itch: Involvement of TRPV1 and TRPA1. J. Allergy Clin. Immunol. 2013, 133, 448–460. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, B.F.; Patsinakidis, N.; Raap, U. Role of the Pruritic Cytokine IL-31 in Autoimmune Skin Diseases. Front. Immunol. 2019, 10, 1383. [Google Scholar] [CrossRef]
- Sonkoly, E.; Müller, A.; Lauerma, A.I.; Pivarcsi, A.; Soto, H.; Kemény, L.; Alenius, H.; Dieu-Nosjean, M.-C.; Meller, S.; Rieker, J.; et al. IL-31: A new link between T cells and pruritus in atopic skin inflammation. J. Allergy Clin. Immunol. 2006, 117, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Takaoka, A.; Arai, I.; Sugimoto, M.; Honma, Y.; Futaki, N.; Nakamura, A.; Nakaike, S. Involvement of IL-31 on scratching behavior in NC/Nga mice with atopic-like dermatitis. Exp. Dermatol. 2006, 15, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Yamamura, K.; Kido-Nakahara, M.; Nakahara, T.; Fukui, Y. Emerging role of interleukin-31 and interleukin-31 receptor in pruritus in atopic dermatitis. Allergy 2017, 73, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Lai, T.; Wu, N.; Li, W.; Chen, M.; Yi, Z.; Huang, D.; Jing, Z.; Lu, Y.; Lv, Q.; Li, N.; et al. Interleukin-31 expression and relation to disease severity in human asthma. Sci. Rep. 2016, 6, 22835. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Ozaki, K.; Baumann, H.; Levin, S.D.; Puel, A.; Farr, A.G.; Ziegler, S.F.; Leonard, W.J.; Lodish, H.F. Cloning of a receptor subunit required for signaling by thymic stromal lymphopoietin. Nat. Immunol. 2000, 1, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, S.F.; Roan, F.; Bell, B.D.; Stoklasek, T.A.; Kitajima, M.; Han, H. Chapter 4—The Biology of Thymic Stromal Lymphopoietin (TSLP). In Advances in Pharmacology; Webb, D.R., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 66, pp. 129–155. [Google Scholar]
- Ziegler, S.F. The role of thymic stromal lymphopoietin (TSLP) in allergic disorders. Curr. Opin. Immunol. 2010, 22, 795–799. [Google Scholar] [CrossRef] [Green Version]
- He, R.; Geha, R.S. Thymic stromal lymphopoietin. Ann. N. Y. Acad. Sci. 2010, 1183, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Wohlmann, A.; Sebastian, K.; Borowski, A.; Krause, S.; Friedrich, K. Signal transduction by the atopy-associated human thymic stromal lymphopoietin (TSLP) receptor depends on Janus kinase function. Boil. Chem. 2010, 391, 181–186. [Google Scholar] [CrossRef]
- Rochman, Y.; Kashyap, M.; Robinson, G.W.; Sakamoto, K.; Gomez-Rodriguez, J.; Wagner, K.-U.; Leonard, W.J. Thymic stromal lymphopoietin-mediated STAT5 phosphorylation via kinases JAK1 and JAK2 reveals a key difference from IL-7-induced signaling. Proc. Natl. Acad. Sci. USA 2010, 107, 19455–19460. [Google Scholar] [CrossRef] [Green Version]
- Arima, K.; Watanabe, N.; Hanabuchi, S.; Chang, M.; Sun, S.-C.; Liu, Y.-J. Distinct Signal Codes Generate Dendritic Cell Functional Plasticity. Sci. Signal. 2010, 3, ra4. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Roan, F.; Ziegler, S.F. The atopic march: Current insights into skin barrier dysfunction and epithelial cell-derived cytokines. Immunol. Rev. 2017, 278, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.R.; The, L.; Batia, L.M.; Beattie, K.; Katibah, G.E.; McClain, S.P.; Pellegrino, M.; Estandian, D.M.; Bautista, D.M. The epithelial cell-derived atopic dermatitis cytokine TSLP activates neurons to induce itch. Cell 2013, 155, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Varricchi, G.; Pecoraro, A.; Marone, G.; Criscuolo, G.; Spadaro, G.; Genovese, A.; Marone, G. Thymic Stromal Lymphopoietin Isoforms, Inflammatory Disorders, and Cancer. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Fornasa, G.; Tsilingiri, K.; Caprioli, F.; Botti, F.; Mapelli, M.; Meller, S.; Kislat, A.; Homey, B.; Di Sabatino, A.; Sonzogni, A.M.; et al. Dichotomy of short and long thymic stromal lymphopoietin isoforms in inflammatory disorders of the bowel and skin. J. Allergy Clin. Immunol. 2015, 136, 413–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quentmeier, H.; Drexler, H.G.; Fleckenstein, D.; Zaborski, M.; Armstrong, A.; Sims, J.; Lyman, S.D. Cloning of human thymic stromal lymphopoietin (TSLP) and signaling mechanisms leading to proliferation. Leuk. 2001, 15, 1286–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjerkan, L.; Schreurs, O.; Engen, S.A.; Jahnsen, F.L.; Baekkevold, E.S.; Blix, I.J.; Schenck, K. The short form of TSLP is constitutively translated in human keratinocytes and has characteristics of an antimicrobial peptide. Mucosal Immunol. 2014, 8, 49–56. [Google Scholar] [CrossRef]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A.; et al. Human epithelial cells trigger dendritic cell-mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673–680. [Google Scholar] [CrossRef]
- Kashyap, M.; Rochman, Y.; Spolski, R.; Samsel, L.; Leonard, W.J. Thymic stromal lymphopoietin is produced by dendritic cells. J. Immunol. 2011, 187, 1207–1211. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, N.; Hanabuchi, S.; Soumelis, V.; Yuan, W.; Ho, S.; Malefyt, R.D.W.; Liu, Y.-J. Human thymic stromal lymphopoietin promotes dendritic cell-mediated CD4+ T cell homeostatic expansion. Nat. Immunol. 2004, 5, 426–434. [Google Scholar] [CrossRef]
- Muñoz-Carrillo, J.L.; Contreras-Cordero, J.F.; Gutiérrez-Coronado, O.; Villalobos-Gutiérrez, P.T.; Ramos-Gracia, L.G.; Hernández-Reyes, V.E. Cytokine Profiling Plays a Crucial Role in Activating Immune System to Clear Infectious Pathogens. In Immune Response Activation and Immunomodulation; IntechOpen: London, UK, 2018. [Google Scholar]
- Misery, L. TSLP, the key of pruritus in atopic dermatitis. Med. Sci. 2014, 30, 142–144. [Google Scholar]
- Kumagai, A.; Kubo, T.; Kawata, K.; Kamekura, R.; Yamashita, K.; Jitsukawa, S.; Nagaya, T.; Sumikawa, Y.; Himi, T.; Yamashita, T.; et al. Keratinocytes in atopic dermatitis express abundant DeltaNp73 regulating thymic stromal lymphopoietin production via NF-kappaB. J. Dermatol. Sci. 2017, 88, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Storan, E.R.; O’Gorman, S.M.; McDonald, I.D.; Steinhoff, M. Role of Cytokines and Chemokines in Itch. Drug Delivery 2015, 226, 163–176. [Google Scholar] [CrossRef]
- Bryant, C.E.; Orr, S.J.; Ferguson, B.; Symmons, M.F.; Boyle, J.P.; Monie, T.P. International Union of Basic and Clinical Pharmacology. XCVI. Pattern Recognition Receptors in Health and Disease. Pharmacol. Rev. 2015, 67, 462–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef] [PubMed]
- Bryant, A.H.; Menzies, G.E.; Scott, L.M.; Spencer-Harty, S.; Davies, L.B.; Smith, R.A.; Jones, R.; Thornton, C.A. Human gestation-associated tissues express functional cytosolic nucleic acid sensing pattern recognition receptors. Clin. Exp. Immunol. 2017, 189, 36–46. [Google Scholar] [CrossRef] [Green Version]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Park, B.S.; Song, D.H.; Kim, H.M.; Choi, B.-S.; Lee, H.; Lee, J.-O. The structural basis of lipopolysaccharide recognition by the TLR4–MD-2 complex. Nature 2009, 458, 1191–1195. [Google Scholar] [CrossRef] [PubMed]
- Shimazu, R.; Akashi, S.; Ogata, H.; Nagai, Y.; Fukudome, K.; Miyake, K.; Kimoto, M. MD-2, a Molecule that Confers Lipopolysaccharide Responsiveness on Toll-like Receptor 4. J. Exp. Med. 1999, 189, 1777–1782. [Google Scholar] [CrossRef] [PubMed]
- Taves, S.; Ji, R.-R. Itch Control by Toll-Like Receptors. Drug Delivery 2015, 226, 135–150. [Google Scholar] [CrossRef]
- Uematsu, S.; Akira, S. Toll-Like Receptors (TLRs) and Their Ligands. Drug Delivery 2008, 183, 1–20. [Google Scholar] [CrossRef]
- De Bouteiller, O.; Merck, E.; Hasan, U.; Hubac, S.; Benguigui, B.; Trinchieri, G.; Bates, E.E.M.; Caux, C. Recognition of Double-stranded RNA by Human Toll-like Receptor 3 and Downstream Receptor Signaling Requires Multimerization and an Acidic pH. J. Boil. Chem. 2005, 280, 38133–38145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Berta, T.; Xu, Z.-Z.; Park, C.-K.; Zhang, L.; Lu, N.; Liu, Q.; Liu, Y.; Gao, Y.-J.; Liu, Y.-C.; et al. TLR3 deficiency impairs spinal cord synaptic transmission, central sensitization, and pruritus in mice. J. Clin. Investig. 2012, 122, 2195–2207. [Google Scholar] [CrossRef]
- Szöllősi, A.G.; McDonald, I.; Szabó, I.L.; Meng, J.; Bogaard, E.V.D.; Steinhoff, M. TLR3 in Chronic Human Itch: A Keratinocyte-Associated Mechanism of Peripheral Itch Sensitization. J. Investig. Dermatol. 2019, 139, 2393–2396. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.H.; Zhang, Y.; Jiang, L.Q.; Wang, S.; Lei, C.Q.; Sun, M.S.; Shu, H.B.; Liu, Y. WDFY1 mediates TLR3/4 signaling by recruiting TRIF. EMBO Rep. 2015, 16, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Vaure, C.; Liu, Y.; Vaure, C. A Comparative Review of Toll-Like Receptor 4 Expression and Functionality in Different Animal Species. Front. Immunol. 2014, 5, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peri, F.; Calabrese, V. Toll-like Receptor 4 (TLR4) Modulation by Synthetic and Natural Compounds: An Update. J. Med. Chem. 2013, 57, 3612–3622. [Google Scholar] [CrossRef]
- Diogenes, A.; Ferraz, C.; Akopian, A.; Henry, M.; Hargreaves, K. LPS Sensitizes TRPV1 via Activation of TLR4 in Trigeminal Sensory Neurons. J. Dent. Res. 2011, 90, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Min, H.; Lee, H.; Lim, H.; Jang, Y.H.; Chung, S.J.; Lee, C.J.; Lee, S.J. TLR4 enhances histamine-mediated pruritus by potentiating TRPV1 activity. Mol. Brain 2014, 7, 59. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Han, Q.; Chen, G.; Huang, Y.; Zhao, L.-X.; Berta, T.; Gao, Y.-J.; Ji, R.-R. Toll-like receptor 4 contributes to chronic itch, alloknesis, and spinal astrocyte activation in male mice. Pain 2016, 157, 806–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-Specific Recognition of Single-Stranded RNA via Toll-like Receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef] [Green Version]
- Roßbach, K.; Bäumer, W. PCR detects bands consistent with the expression of receptors associated with pruritus in canine dorsal root ganglia. Veter—Dermatol. 2013, 25. [Google Scholar] [CrossRef]
- Liu, T.; Xu, Z.-Z.; Park, C.-K.; Berta, T.; Ji, R.-R. Toll-like receptor 7 mediates pruritus. Nat. Neurosci. 2010, 13, 1460–1462. [Google Scholar] [CrossRef] [PubMed]
- Larson, P.; Kucaba, T.A.; Xiong, Z.; Olin, M.; Griffith, T.S.; Ferguson, D.M. Design and Synthesis of N1-Modified Imidazoquinoline Agonists for Selective Activation of Toll-like Receptors 7 and 8. ACS Med. Chem. Lett. 2017, 8, 1148–1152. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-J.; Park, G.H.; Kim, N.; Lee, J.; Min, H.; Wall, E.; Lee, C.J.; Simon, M.I.; Lee, S.J.; Han, S.-K. Analysis of cellular and behavioral responses to imiquimod reveals a unique itch pathway in transient receptor potential vanilloid 1 (TRPV1)-expressing neurons. Proc. Natl. Acad. Sci. USA 2011, 108, 3371–3376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flock, T.; Hauser, A.S.; Lund, N.; Gloriam, D.E.; Balaji, S.; Babu, M.M. Selectivity determinants of GPCR–G-protein binding. Nature 2017, 545, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Gschwandtner, M.; Purwar, R.; Wittmann, M.; Bäumer, W.; Kietzmann, M.; Werfel, T.; Gutzmer, R. Histamine Upregulates Keratinocyte MMP-9 Production via the Histamine H1 Receptor. J. Investig. Dermatol. 2008, 128, 2783–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Yamagata, K.; Nakayamada, S.; Sawamukai, N.; Yamaoka, K.; Sakata, K.; Nakano, K.; Tanaka, Y. Histamine inhibits differentiation of skin fibroblasts into myofibroblasts. Biochem. Biophys. Res. Commun. 2015, 463, 434–439. [Google Scholar] [CrossRef]
- Migalovich-Sheikhet, H.; Friedman, S.; Mankuta, D.; Levi-Schaffer, F. Novel Identified Receptors on Mast Cells. Front. Immunol. 2012, 3, 238. [Google Scholar] [CrossRef] [Green Version]
- Gangwar, R.S.; Landolina, N.; Arpinati, L.; Levi-Schaffer, F. Mast cell and eosinophil surface receptors as targets for anti-allergic therapy. Pharmacol. Ther. 2017, 170, 37–63. [Google Scholar] [CrossRef]
- Zschiebsch, K.; Fischer, C.; Wilken-Schmitz, A.; Geisslinger, G.; Channon, K.M.; Watschinger, K.; Tegeder, I. Mast cell tetrahydrobiopterin contributes to itch in mice. J. Cell. Mol. Med. 2018, 23, 985–1000. [Google Scholar] [CrossRef]
- Matsuda, N.; Jesmin, S.; Takahashi, Y.; Hatta, E.; Kobayashi, M.; Matsuyama, K.; Kawakami, N.; Sakuma, I.; Gando, S.; Fukui, H.; et al. Histamine H1 and H2 Receptor Gene and Protein Levels Are Differentially Expressed in the Hearts of Rodents and Humans. J. Pharmacol. Exp. Ther. 2004, 309, 786–795. [Google Scholar] [CrossRef]
- Booth, R.G.; Moniri, N.H.; Bakker, R.A.; Choksi, N.Y.; Nix, W.B.; Timmerman, H.; Leurs, R. A novel phenylaminotetralin radioligand reveals a subpopulation of histamine H(1) receptors. J. Pharmacol. Exp. Ther. 2002, 302, 328–336. [Google Scholar] [CrossRef]
- Heinrich, T.; Böttcher, H.; Gericke, R.; Bartoszyk, G.D.; Anzali, S.; Seyfried, C.A.; Greiner, H.E.; Van Amsterdam, C. Synthesis and Structure—Activity Relationship in a Class of Indolebutylpiperazines as Dual 5-HT1AReceptor Agonists and Serotonin Reuptake Inhibitors. J. Med. Chem. 2004, 47, 4684–4692. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Wortsman, J.; Tobin, D.J. The cutaneous serotoninergic/melatoninergic system: Securing a place under the sun. FASEB J. 2005, 19, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Nordlind, K.; Azmitia, E.C.; Slominski, A.T. The skin as a mirror of the soul: Exploring the possible roles of serotonin. Exp. Dermatol. 2008, 17, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Ahern, G.P. 5-HT and the immune system. Curr. Opin. Pharmacol. 2011, 11, 29–33. [Google Scholar] [CrossRef]
- Shajib, M.S.; Khan, W.I. The role of serotonin and its receptors in activation of immune responses and inflammation. Acta Physiol. 2014, 213, 561–574. [Google Scholar] [CrossRef]
- Sleight, A.J.; Stam, N.J.; Mutel, V.; Vanderheyden, P.M. Radiolabelling of the human 5-HT2A receptor with an agonist, a partial agonist and an antagonist: Effects on apparent agonist affinities. Biochem. Pharmacol. 1996, 51, 71–76. [Google Scholar] [CrossRef]
- Bard, J.A.; Zgombick, J.; Adham, N.; Vaysse, P.; Branchek, T.A.; Weinshank, R.L. Cloning of a novel human serotonin receptor (5-HT7) positively linked to adenylate cyclase. J. Boil. Chem. 1993, 268, 23422–23426. [Google Scholar]
- McKelvey, K.; Jackson, C.J.; Xue, M. Activated protein C: A regulator of human skin epidermal keratinocyte function. World J. Boil. Chem. 2014, 5, 169–179. [Google Scholar]
- Duitman, J.; Ruela-De-Sousa, R.R.; Shi, K.; De Boer, O.J.; Borensztajn, K.S.; Florquin, S.; Peppelenbosch, M.P.; Spek, C.A. Protease Activated Receptor-1 Deficiency Diminishes Bleomycin-Induced Skin Fibrosis. Mol. Med. 2014, 20, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Regard, J.B.; Sato, I.T.; Coughlin, S.R. Anatomical Profiling of G Protein-Coupled Receptor Expression. Cell 2008, 135, 561–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rattenholl, A.; Steinhoff, M. Proteinase-activated receptor-2 in the skin: Receptor expression, activation and function during health and disease. Drug News Perspect. 2008, 21, 369. [Google Scholar] [CrossRef]
- Kuckleburg, C.J.; Newman, P.J. Neutrophil proteinase 3 acts on protease-activated receptor-2 to enhance vascular endothelial cell barrier function. Arter. Thromb. Vasc. Boil. 2012, 33, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Mumaw, M.M. A Novel Approach To Antiplatelet Therapy: Targeting Protease Activated Receptor 4. Ph.D. Thesis, Case Western Reserve University, Cleveland, Ohio, 2015. [Google Scholar]
- Liu, J.Y.; Hu, J.H.; Zhu, Q.G.; Li, F.Q.; Sun, H.-J. Substance P receptor expression in human skin keratinocytes and fibroblasts. Br. J. Dermatol. 2006, 155, 657–662. [Google Scholar] [CrossRef]
- Staniek, V.; Misery, L.; Péguet-Navarro, J.; Abello, J.; Doutremepuich, J.-D.; Claudy, A.; Schmitt, D. Binding and in vitro modulation of human epidermal Langerhans cell functions by substance P. Arch. Dermatol. Res. 1997, 289, 285–291. [Google Scholar] [CrossRef]
- Cunin, P.; Caillon, A.; Corvaisier, M.; Garo, E.; Scotet, M.; Blanchard, S.; Delneste, Y.; Jeannin, P. The tachykinins substance P and hemokinin-1 favor the generation of human memory Th17 cells by inducing IL-1beta, IL-23, and TNF-like 1A expression by monocytes. J. Immunol. 2011, 186, 4175–4182. [Google Scholar] [CrossRef] [Green Version]
- Ho, W.-Z.; Lai, J.P.; Zhu, X.H.; Uvaydova, M.; Douglas, S.D. Human monocytes and macrophages express substance P and neurokinin-1 receptor. J. Immunol. 1997, 159, 5654–5660. [Google Scholar]
- Bellucci, F.; Carini, F.; Catalani, C.; Cucchi, P.; Lecci, A.; Meini, S.; Patacchini, R.; Quartara, L.; Ricci, R.; Tramontana, M.; et al. Pharmacological profile of the novel mammalian tachykinin, hemokinin 1. Br. J. Pharmacol. 2002, 135, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Wu, L.H.; Oxender, D.L.; Chung, F.Z. The unpredicted high affinities of a large number of naturally occurring tachykinins for chimeric NK1/NK3 receptors suggest a role for an inhibitory domain in determining receptor specificity. J. Biol. Chem. 1996, 271, 20250–20257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarau, H.M.; Griswold, D.E.; Potts, W.; Foley, J.J.; Schmidt, D.B.; Webb, E.F.; Martin, L.D.; Brawner, M.E.; Elshourbagy, N.A.; Medhurst, A.D.; et al. Nonpeptide tachykinin receptor antagonists: I. Pharmacological and pharmacokinetic characterization of SB 223412, a novel, potent and selective neurokinin-3 receptor antagonist. J. Pharmacol. Exp. Ther. 1997, 281, 1303–1311. [Google Scholar] [PubMed]
- Matus, C.E.; Ehrenfeld, P.; Pavicic, F.; González, C.B.; Concha, M.; Bhoola, K.D.; Burgos, R.A.; Figueroa, C. Activation of the human keratinocyte B1 bradykinin receptor induces expression and secretion of metalloproteases 2 and 9 by transactivation of epidermal growth factor receptor. Exp. Dermatol. 2016, 25, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.K. Chapter 7—Cell-Based Alzheimer’s Disease Biomarkers. In Biomarkers in Alzheimer’s Disease; Khan, T.K., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 221–256. [Google Scholar]
- Prat, A.; Weinrib, L.; Becher, B.; Poirier, J.; Duquette, P.; Couture, R.; Antel, J.P. Bradykinin B1 receptor expression and function on T lymphocytes in active multiple sclerosis. Neurology 1999, 53, 2087. [Google Scholar] [CrossRef] [PubMed]
- Hess, J.F.; Derrick, A.W.; MacNeil, T.; Borkowski, J.A. The agonist selectivity of a mouse B1 bradykinin receptor differs from human and rabbit B1 receptors. Immunopharmacology 1996, 33, 1–8. [Google Scholar] [CrossRef]
- Hess, J.F.; Borkowski, J.A.; MacNeil, T.; Stonesifer, G.Y.; Fraher, J.; Strader, C.D.; Ransom, R.W. Differential pharmacology of cloned human and mouse B2 bradykinin receptors. Mol. Pharmacol. 1994, 45, 1–8. [Google Scholar] [PubMed]
- Jones, C.; Phillips, E.; Davis, C.; Arbuckle, J.; Yaqoob, M.; Burgess, G.M.; Docherty, R.J.; Webb, M.; Bevan, S.; McIntyre, P. Molecular characterisation of cloned bradykinin B1 receptors from rat and human. Eur. J. Pharmacol. 1999, 374, 423–433. [Google Scholar] [CrossRef]
- Hou, Q.; Barr, T.; Gee, L.; Vickers, J.; Wymer, J.; Borsani, E.; Rodella, L.F.; Getsios, S.; Burdo, T.; Eisenberg, E.; et al. Keratinocyte expression of calcitonin gene-related peptide β: Implications for neuropathic and inflammatory pain mechanisms. Pain 2011, 152, 2036–2051. [Google Scholar] [CrossRef] [Green Version]
- Roggenkamp, D.; Köpnick, S.; Stäb, F.; Wenck, H.; Schmelz, M.; Neufang, G. Epidermal Nerve Fibers Modulate Keratinocyte Growth via Neuropeptide Signaling in an Innervated Skin Model. J. Investig. Dermatol. 2013, 133, 1620–1628. [Google Scholar] [CrossRef] [Green Version]
- Bernardo, H.; Vanina, T.-V.; Paola, M.; Jerônimo, F.; Silvana, G.; Lydia, F. The action of CGRP and SP on cultured skin fibroblasts. Op. L. Sci. 2014, 9, 717–726. [Google Scholar]
- Upton, P.D.; Austin, C.; Taylor, G.M.; Nandha, K.A.; Clark, A.J.; Ghatei, M.A.; Bloom, S.R.; Smith, D.M. Expression of adrenomedullin (ADM) and its binding sites in the rat uterus: Increased number of binding sites and ADM messenger ribonucleic acid in 20-day pregnant rats compared with nonpregnant rats. Endocrinology 1997, 138, 2508–2514. [Google Scholar] [CrossRef]
- Aiyar, N.; Disa, J.; Pullen, M.; Nambi, P. Receptor activity modifying proteins interaction with human and porcine calcitonin receptor-like receptor (CRLR) in HEK-293 cells. Mol. Cell. Biochem. 2001, 224, 123–133. [Google Scholar] [CrossRef] [PubMed]
- McLatchie, L.M.; Fraser, N.J.; Main, M.J.; Wise, A.; Brown, J.; Thompson, N.; Solari, R.; Lee, M.G.; Foord, S.M. RAMPs regulate the transport and ligand specificity of the calcitonin-receptor-like receptor. Nature 1998, 393, 333–339. [Google Scholar] [CrossRef]
- Hong, Y.; Hay, D.L.; Quirion, R.; Poyner, D.R. The pharmacology of adrenomedullin 2/intermedin. Br. J. Pharmacol. 2012, 166, 110–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nodin, C.; Vauquelin, G.; Von Mentzer, B. Cys2, 7EtalphaCGRP is a potent agonist for CGRP1 receptors in SK-N-MC cells. Biochem. Pharmacol. 2005, 69, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Southern, C.; Cook, J.M.; Neetoo-Isseljee, Z.; Taylor, D.L.; Kettleborough, C.A.; Merritt, A.; Bassoni, D.L.; Raab, W.J.; Quinn, E.; Wehrman, T.S.; et al. Screening β-arrestin recruitment for the identification of natural ligands for orphan G-protein-coupled receptors. J Biomol Screen. 2013, 18, 599–609. [Google Scholar] [CrossRef] [Green Version]
- Yokomizo, T. Two distinct leukotriene B4 receptors, BLT1 and BLT2. J. Biochem. 2014, 157, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Bäck, M.; Bu, D.-X.; Bränström, R.; Sheikine, Y.; Yan, Z.-Q.; Hansson, G.K. Leukotriene B4 signaling through NF- B-dependent BLT1 receptors on vascular smooth muscle cells in atherosclerosis and intimal hyperplasia. Proc. Natl. Acad. Sci. USA 2005, 102, 17501–17506. [Google Scholar] [CrossRef] [Green Version]
- Toda, A.; Terawaki, K.; Yamazaki, S.; Saeki, K.; Shimizu, T.; Yokomizo, T. Attenuated Th1 induction by dendritic cells from mice deficient in the leukotriene B4 receptor 1. Biochemical 2010, 92, 682–691. [Google Scholar] [CrossRef]
- Huang, W.-W.; Garcia-Zepeda, E.A.; Sauty, A.; Oettgen, H.C.; Rothenberg, M.E.; Luster, A.D. Molecular and Biological Characterization of the Murine Leukotriene B4 Receptor Expressed on Eosinophils. J. Exp. Med. 1998, 188, 1063–1074. [Google Scholar] [CrossRef] [Green Version]
- Runarsson, G.; Liu, A.; Mahshid, Y.; Feltenmark, S.; Pettersson, A.; Klein, E.; Björkholm, M.; Claesson, H.-E. Leukotriene B4 plays a pivotal role in CD40-dependent activation of chronic B lymphocytic leukemia cells. Blood 2005, 105, 1274–1279. [Google Scholar] [CrossRef]
- Yokomizo, T.; Izumi, T.; Chang, K.; Takuwa, Y.; Shimizu, T. A G-protein-coupled receptor for leukotriene B4 that mediates chemotaxis. Nature 1997, 387, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Yokomizo, T.; Kato, K.; Terawaki, K.; Izumi, T.; Shimizu, T. A second leukotriene B(4) receptor, BLT2. A new therapeutic target in inflammation and immunological disorders. J. Exp. Med. 2000, 192, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Man, M.Q.; Hatano, Y.; Lee, S.H.; Man, M.; Chang, S.; Feingold, K.R.; Leung, D.Y.; Holleran, W.; Uchida, Y.; Elias, P.M. Characterization of a hapten-induced, murine model with multiple features of atopic dermatitis: Structural, immunologic, and biochemical changes following single versus multiple oxazolone challenges. J. Investig. Dermatol. 2008, 128, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeki, K.; Yokomizo, T. Identification, signaling, and functions of LTB 4 receptors. Semin. Immunol. 2017, 33, 30–36. [Google Scholar] [CrossRef]
- Ishii, Y.; Saeki, K.; Liu, M.; Sasaki, F.; Koga, T.; Kitajima, K.; Meno, C.; Okuno, T.; Yokomizo, T. Leukotriene B 4 receptor type 2 (BLT2) enhances skin barrier function by regulating tight junction proteins. FASEB J. 2015, 30, 933–947. [Google Scholar] [CrossRef] [Green Version]
- Michel, L.; Denizot, Y.; Thomas, Y.; Jean-Louis, F.; Heslan, M.; Benveniste, J.; Dubertret, L. Production of Paf-Acether by Human Epidermal Cells. J. Investig. Dermatol. 1990, 95, 576–581. [Google Scholar] [CrossRef] [Green Version]
- Ye, R.D.; Prossnitz, E.R.; Zou, A.; Cochrane, C.G. Characterization of a human cDNA that encodes a functional receptor for platelet activating factor. Biochem. Biophys. Res. Commun. 1991, 180, 105–111. [Google Scholar] [CrossRef]
- Nakamura, M.; Honda, Z.; Izumi, T.; Sakanaka, C.; Mutoh, H.; Minami, M.; Bito, H.; Seyama, Y.; Matsumoto, T.; Noma, M. Molecular cloning and expression of platelet-activating factor receptor from human leukocytes. J. Boil. Chem. 1991, 266, 20400–20405. [Google Scholar]
- Aoki, Y.; Nakamura, M.; Kodama, H.; Matsumoto, T.; Shimizu, T.; Noma, M. A radioreceptor binding assay for platelet-activating factor (PAF) using membranes from CHO cells expressing human PAF receptor. J. Immunol. Methods 1995, 186, 225–231. [Google Scholar] [CrossRef]
- Bigliardi, P.L.; Bigliardi-Qi, M.; Buechner, S.; Rufli, T. Expression of mu-opiate receptor in human epidermis and keratinocytes. J. Inv. Dermatol. 1998, 111, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Ständer, S.; Gunzer, M.; Metze, D.; Luger, T. Steinhoff M. Localization of mu-opioid receptor 1A on sensory nerve fibers in human skin. Regul. Pept. 2002, 110, 75–83. [Google Scholar] [CrossRef]
- Chuang, T.; Killam, K.; Chuang, L.; Kung, H.; Sheng, W.; Chao, C.; Yu, L.; Chuang, R. Mu Opioid Receptor Gene Expression in Immune Cells. Biochem. Biophys. Res. Commun. 1995, 216, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Toll, L.; Berzetei-Gurske, I.P.; Polgar, W.E.; Brandt, S.R.; Adapa, I.D.; Rodriguez, L.; Schwartz, R.W.; Haggart, D.; O’Brien, A.; White, A.; et al. Standard binding and functional assays related to medications development division testing for potential cocaine and opiate narcotic treatment medications. NIDA Res. Monogr. 1998, 178, 440–466. [Google Scholar]
- Raynor, K.; Kong, H.; Chen, Y.; Yasuda, K.; Yu, L.; Bell, G.I.; Reisine, T. Pharmacological characterization of the cloned kappa-, delta-, and mu-opioid receptors. Mol. Pharmacol. 1994, 45, 330–334. [Google Scholar]
- Ganesh, A.; Maxwell, L.G. Pathophysiology and management of opioid-induced pruritus. Drugs 2007, 67, 2323–2333. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Liu, H.-W.; Fu, X.-B.; Sheng, Z.-Y.; Li, J.-F. Coexistence and upregulation of three types of opioid receptors, mu, delta and kappa, in human hypertrophic scars. Br. J. Dermatol. 2008, 158, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Salemi, S.; Aeschlimann, A.; Reisch, N.; Jüngel, A.; Gay, R.E.; Heppner, F.L.; Michel, B.A.; Gay, S.; Sprott, H. Detection of kappa and delta opioid receptors in skin—Outside the nervous system. Biochem. Biophys. Res. Commun. 2005, 338, 1012–1017. [Google Scholar] [CrossRef]
- Gavériaux, C.; Peluso, J.; Simonin, F.; Laforet, J.; Kieffer, B. Identification of kappa- and delta-opioid receptor transcripts in immune cells. FEBS Lett. 1995, 369, 272–276. [Google Scholar]
- Zhu, J.; Chen, C.; Xue, J.C.; Kunapuli, S.; Deriel, J.K.; Liu-Chen, L.Y. Cloning of a human kappa opioid receptor from the brain. Life Sci. 1995, 56, 201–207. [Google Scholar] [CrossRef]
- Yasuda, K.; Raynor, K.; Kong, H.; Breder, C.D.; Takeda, J.; Reisine, T.; Bell, G.I. Cloning and functional comparison of kappa and delta opioid receptors from mouse brain. Proc. Natl. Acad. Sci. USA 1993, 90, 6736–6740. [Google Scholar] [CrossRef] [Green Version]
- Wikström, B.; Gellert, R.; Ladefoged, S.D.; Danda, Y.; Akai, M.; Ide, K.; Ogasawara, M.; Kawashima, Y.; Ueno, K.; Mori, A.; et al. κ-Opioid System in Uremic Pruritus: Multicenter, Randomized, Double-Blind, Placebo-Controlled Clinical Studies. J. Am. Soc. Nephrol. 2005, 16, 3742–3747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bort, A.; Alvarado-Vazquez, P.A.; Moracho-Vilrriales, C.; Virga, K.G.; Gumina, G.; Romero-Sandoval, E.A.; Asbill, S. Effects of JWH015 in cytokine secretion in primary human keratinocytes and fibroblasts and its suitability for topical/transdermal delivery. Mol. Pain 2017, 13. [Google Scholar] [CrossRef] [Green Version]
- Herkenham, M.; Lynn, A.B.; Little, M.D.; Johnson, M.R.; Melvin, L.S.; De Costa, B.R.; Rice, K.C. Cannabinoid receptor localization in brain. Proc. Natl. Acad. Sci. USA 1990, 87, 1932–1936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechoulam, R.; Ben-Shabat, S.; Hanuš, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R.; et al. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 1995, 50, 83–90. [Google Scholar] [CrossRef]
- Showalter, V.M.; Compton, D.R.; Martin, B.R.; Abood, M.E. Evaluation of binding in a transfected cell line expressing a peripheral cannabinoid receptor (CB2): Identification of cannabinoid receptor subtype selective ligands. J. Pharmacol. Exp. Ther. 1996, 278, 989–999. [Google Scholar] [PubMed]
- Schlosburg, J.E.; O’Neal, S.T.; Conrad, D.H.; Lichtman, A.H. CB1 receptors mediate rimonabant-induced pruritic responses in mice: Investigation of locus of action. Psychopharmacology 2011, 216, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Arenas-Ramirez, N.; Woytschak, J.; Boyman, O. Interleukin-2: Biology, Design and Application. Trends Immunol. 2015, 36, 763–777. [Google Scholar] [CrossRef] [Green Version]
- Junghans, V.; Jung, T.; Neumann, C. Human keratinocytes constitutively express IL-4 receptor molecules and respond to IL-4 with an increase in B7/BB1 expression. Ex. Dermato. 1996, 5, 316–324. [Google Scholar] [CrossRef]
- McCormick, S.; Heller, N. Commentary: IL-4 and IL-13 receptors and signaling. Cytokine 2015, 75, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Graber, P.; Gretener, D.; Herren, S.; Aubry, J.P.; Elson, G.; Poudrier, J.; Lecoanet-Henchoz, S.; Alouani, S.; Losberger, C.; Bonnefoy, J.Y.; et al. The distribution of IL-13 receptor alpha1 expression on B cells, T cells and monocytes and its regulation by IL-13 and IL-4. Eur. J. Immunol. 1998, 28, 4286–4298. [Google Scholar] [CrossRef]
- Kasraie, S.; Niebuhr, M.; Baumert, K.; Werfel, T. Functional effects of interleukin 31 in human primary keratinocytes. Allergy 2011, 66, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Jawa, R.S.; Chattopadhyay, S.; Tracy, E.; Wang, Y.; Huntoon, K.; Dayton, M.T.; Baumann, H. Regulated Expression of the IL-31 Receptor in Bronchial and Alveolar Epithelial Cells, Pulmonary Fibroblasts, and Pulmonary Macrophages. J. Interf. Cytokine Res. 2008, 28, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Ghilardi, N.; Li, J.; Hongo, J.-A.; Yi, S.; Gurney, A.; De Sauvage, F.J. A Novel Type I Cytokine Receptor Is Expressed on Monocytes, Signals Proliferation, and Activates STAT-3 and STAT-5. J. Boil. Chem. 2002, 277, 16831–16836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosley, B.; De Imus, C.; Friend, D.; Boiani, N.; Thoma, B.; Park, L.S.; Cosman, D. Dual oncostatin M (OSM) receptors. Cloning and characterization of an alternative signaling subunit conferring OSM-specific receptor activation. J. Biol. Chem. 1996, 271, 32635–32643. [Google Scholar] [CrossRef] [Green Version]
- Leyva-Castillo, J.M.; Hener, P.; Jiang, H.; Li, M. TSLP Produced by Keratinocytes Promotes Allergen Sensitization through Skin and Thereby Triggers Atopic March in Mice. J. Investig. Dermatol. 2013, 133, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Fitzgerald, K.A.; O’Neill, L.A.; Gearing, A.J.; Callard, R.E. TSLP. In The Cytokine FactsBook and Webfacts; Elsevier BV: Amsterdam, The Netherlands, 2001; pp. 490–492. [Google Scholar]
- Lebre, M.C.; Van Der Aar, A.M.; Van Baarsen, L.; Van Capel, T.M.; Schuitemaker, J.H.; Kapsenberg, M.L.; De Jong, E.C. Human Keratinocytes Express Functional Toll-Like Receptor 3, 4, 5, and 9. J. Investig. Dermatol. 2007, 127, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.S. Toll-like receptors in skin. Adv. Dermatol. 2008, 24, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Lafon, M.; Mégret, F.; Lafage, M.; Prehaud, C. The Innate Immune Facet of Brain: Human Neurons Express TLR-3 and Sense Viral dsRNA. J. Mol. Neurosci. 2006, 29, 185–194. [Google Scholar] [CrossRef]
- Medzhitov, R.; Preston-Hurlburt, P.; Janeway, C.A. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature 1997, 388, 394–397. [Google Scholar] [CrossRef]
- Tatematsu, M.; Yoshida, R.; Morioka, Y.; Ishii, N.; Funami, K.; Watanabe, A.; Saeki, K.; Seya, T.; Matsumoto, M. Raftlin Controls Lipopolysaccharide-Induced TLR4 Internalization and TICAM-1 Signaling in a Cell Type-Specific Manner. J. Immunol. 2016, 196, 3865–3876. [Google Scholar] [CrossRef] [Green Version]
- Sibaud, V.; Lebœuf, N.R.; Roche, H.; Belum, V.R.; Gladieff, L.; Deslandres, M.; Montastruc, M.; Eche, A.; Vigarios, E.; Dalenc, F.; et al. Dermatological adverse events with taxane chemotherapy. Eur. J. Dermatol. EJD 2016, 26, 427–443. [Google Scholar] [CrossRef] [PubMed]
- Pendergraft, W.F.; Means, T.K. Chapter 18—Toll-Like Receptors, Systemic Lupus Erythematosus. In Systemic Lupus Erythematosus; Tsokos, G.C., Ed.; Academic Press: Boston, MA, USA; Cambridge, MA, USA, 2016; pp. 143–151. [Google Scholar]
- Feng, J.; Hu, H. A novel player in the field: Merkel disc in touch, itch and pain. Exp. Dermatol. 2019, 28, 1412–1415. [Google Scholar] [CrossRef] [Green Version]
- Sakai, K.; Akiyama, T. Disinhibition of Touch-Evoked Itch in a Mouse Model of Psoriasis. J. Investig. Dermatol. 2019, 139, 1407–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Luo, J.; Yang, P.; Du, J.; Kim, B.S.; Hu, H. Piezo2 channel—Merkel cell signaling modulates the conversion of touch to itch. Science 2018, 360, 530–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acton, D.; Ren, X.; Di Costanzo, S.; Dalet, A.; Bourane, S.; Bertocchi, I.; Eva, C.; Goulding, M. Spinal Neuropeptide Y1 Receptor-Expressing Neurons Form an Essential Excitatory Pathway for Mechanical Itch. Cell Rep. 2019, 28, 625–639.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, H.; Fatima, M.; Li, A.; Lee, H.; Cai, W.; Horwitz, L.; Hor, C.C.; Zaher, N.; Cin, M.; Slade, H.; et al. Identification of a Spinal Circuit for Mechanical and Persistent Spontaneous Itch. Neuron 2019, 103, 1135–1149. e6. [Google Scholar] [CrossRef]
- Xu, Z.-Z.; Kim, Y.H.; Bang, S.; Zhang, Y.; Berta, T.; Wang, F.; Oh, S.B.; Ji, R.-R. Inhibition of mechanical allodynia in neuropathic pain by TLR5-mediated A-fiber blockade. Nat. Med. 2015, 21, 1326–1331. [Google Scholar] [CrossRef]
- Cheng, R.-X.; Feng, Y.; Liu, D.; Wang, Z.-H.; Zhang, J.-T.; Chen, L.-H.; Su, C.-J.; Wang, B.; Huang, Y.; Ji, R.-R.; et al. The role of Nav1.7 and methylglyoxal-mediated activation of TRPA1 in itch and hypoalgesia in a murine model of type 1 diabetes. Theranostics 2019, 9, 4287–4307. [Google Scholar] [CrossRef]




{kind=link}
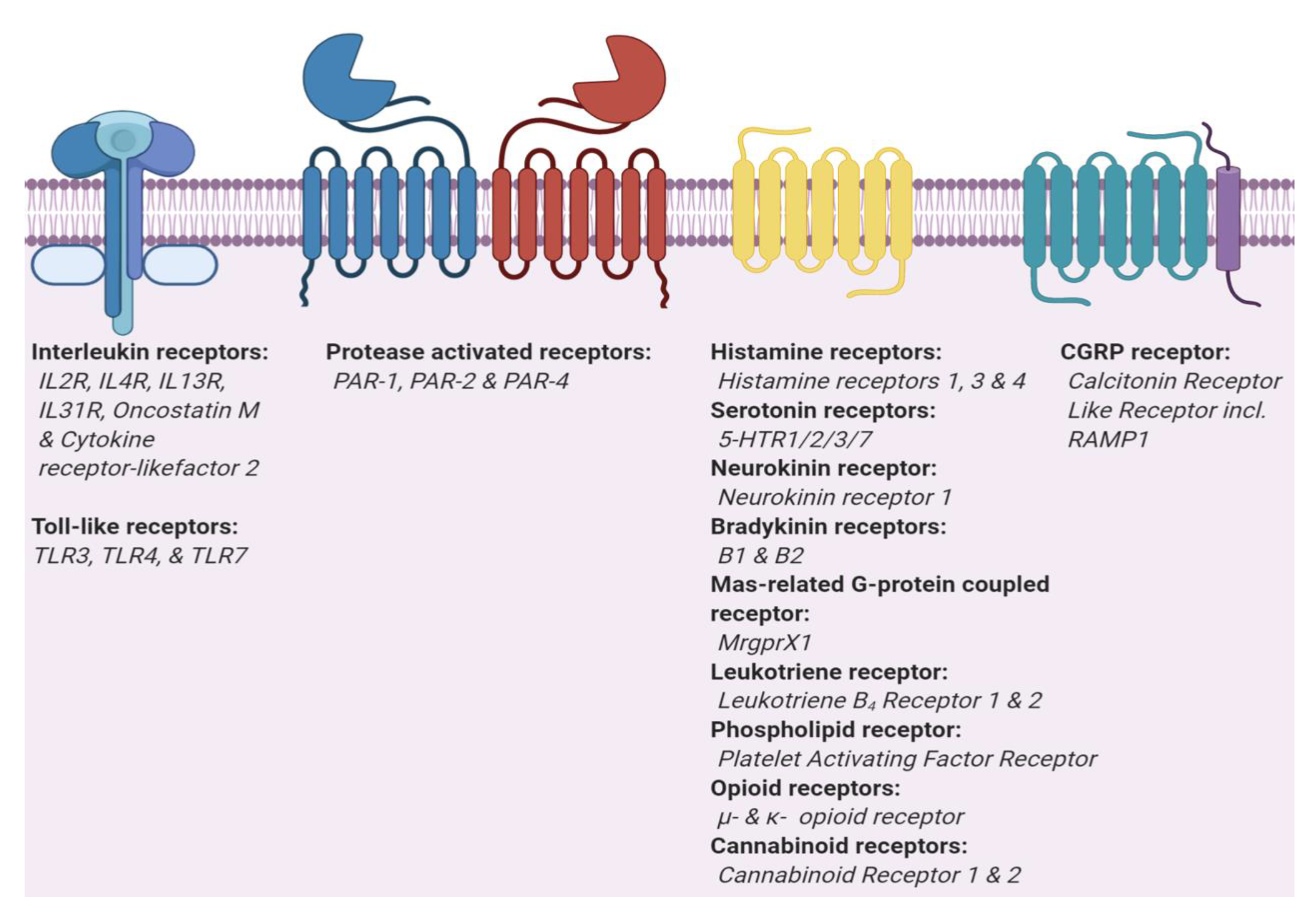{kind=link}
{kind=link}
| ME | ME | ME | ME | MO | MO | MO | MO | MO | ME | ME | ME | ME | MO | MO | |
| endogenous pruritogens | Thrombin | Trypsin | Thrombin | Bradykinin, kallidin | Leukotriene B4 | Leukotriene B4 | Bradykinin, kallidin | IL-4 | IL-13 | IL-31 | IL-2 | Serotonin | BAM8-22 | Calcitonin gene-related peptide | TSLP |
| receptor | PAR1 | PAR2 | PAR4 | BDKRB1 | BLT1 | BLT2 | BDKRB2 | IL4R | IL13R | IL31R | IL2R | 5-HTR7 | MRGPRX1 | CGRP | TSLPR |
| receptor signaling | Gi/Go, Gq/G11, G12/G13 | Gi/Go, G12/G13 | Gi/Go, Gq/G11 | Gi/Go | Gs, Gi/Go, Gq/G11 | JAK1/2/3 | JAK1 | JAK1/2 | JAK1/3 | Gs, Gq/G11, G12/G13 | Gs | JAK1/2 | |||
| ion channel | TRPA1, TRPV1 | - | TRPA1 | ||||||||||||
| ME | MO | MO | MO | MO | ME | ME | MO | MO | MO | MO | MO | ||||
| endogenous pruritogens | endorphins | dynorphins and endorphines | endocanna-binoids | endocanna-binoids | platelet-activating factor | substance P | Histamine | Oncostatin M | ds RNA | LPS | ss RNA | Serotonin | |||
| receptor | OPRM | OPRK | CB1 | CB2 | PAFR | NK1R | H1R | OSMR | TLR3 | TLR4 | TLR7 | 5-HTR2 | |||
| receptor signaling | Gi/Go | Gi/Go, G12/G13 | Gs, Gi/Go, Gq/G11 | Gs, Gq/G11 | Gi/Go, Gq/G11 | JAK1/2 | TRIF | MyD88/TRIF | MyD88 | Gi/Go, Gq/G11 | |||||
| ion channel | TRPV1 | TRPV4 | |||||||||||||
| Receptors | Expression | G-Protein Binding [289] | Endogenous Pruritogens | Exogenous Puritogens | |||
|---|---|---|---|---|---|---|---|
| Gs | Gi/Go | Gq/G11 | G12/G13 | ||||
| 5|18 | 14|18 | 10|18 | 6|18 | ||||
| Histamine receptor | GPCR | Monoamine | |||||
| H1R | Keratinocytes [290], dermal fibroblasts [291], Granulocytes, mast cells [292,293,294] smooth muscles, endothelial cells, CNS, and cardiovascular system [295] | Histamine [296] | vilazodone [297] | ||||
| Serotonin receptor | GPCRs | Monoamine | |||||
| 5-HTR2 | Keratinocytes [298], dermal fibroblasts [299], dermal langerhans cells [74] T cells, B cells, granulocytes, dendritic cells, macrophages, and monocytes [300,301] | 5-hydroxytryptamine [302] | alpha-methylserotonin [67] | ||||
| 5-HTR7 | Keratinocytes [298], dermal fibroblasts [299], dermal Langerhans cells [74], T cells, B cells, granulocytes, macrophages, monocytes, and dendritic cells [301] | 5-hydroxytryptamine [303] | LP44 [69] | ||||
| Protease-activated receptors | GPCRs | Proteases | |||||
| PAR1 | Keratinocytes [304], dermal fibroblast [305], platelets, and vascular endothelial cells [306] | Thrombin [83] | Hexapeptide derived from thedered peptide sequence e.g., TFLLR [85] bromelain, ficin, papain, mucunain, and trypsin from animals [91] | ||||
| PAR2 | Keratinocytes [98], dermal fibroblast [307], endothelial cells in pancreas, liver, kidney, GI tract, and colon but not in the brain or skeletal muscle [306,308] | Trypsin [83] | Hexapeptide derived from thedered peptide sequence e.g., SLIGRL [85] bromelain, ficin, papain, mucunain, and trypsin from animals [91] | ||||
| PAR4 | Platelets, lung, thyroid, testis, small intestin, and pancreas [306,309] | Thrombin [83] | Hexapeptide derived from thedered peptide sequence e.g., AYPGKF [85] bromelain, ficin, papain, mucunain, and trypsin from animals [91] | ||||
| Neurokinin receptors | GPCR | Peptides | |||||
| NK1R | Keratinocytes, dermal fibroblast [310], dermal Langerhans cells [311], T cells, macrophages, and monocytes [312,313] | substance P, neurokinin A, neurokinin B, neuropeptide-γ, neuropeptide K [314,315,316] | N/A | ||||
| Bradykinin receptors | GPCRs | Peptides | |||||
| BDKRB1 | Keratinocytes [317], dermal fibroblasts [318], Granulocytes, and T lymphocytes [319] | bradykinin, kallidin, T-kinin, [des-Arg10] kallidin [320,321,322] | diphenylcyclopropenone [123] | ||||
| BDKRB2 | N/A | bradykinin, kallidin, T-kinin, [des-Arg10] kallidin [320,321,322] | sodium deoxycholic acid [117] | ||||
| Calcitonin receptor-like receptor | GPCR receptors | Peptides | |||||
| CGRP | Keratinocytes [323,324], dermal fibroblasts [325], Lung, uterus, and placenta [326] | adrenomedullin, adrenomedullin 2/intermedin, α-CGRP, β-CGRP [327,328,329] | [Cys(Et)2,7]α-CGRP [330] | ||||
| Mas-related GPR family | GPCR | Peptide | |||||
| MRGPRX1 | Small dorsal root, and trigeminal sensory neurons [144] | BAM8-22 [331] | chloroquine [21] | ||||
| Leukotriene receptors | GPCRs | Leukotrienes | |||||
| BLT1 | Keratinocytes [332], Granulocytes, B-lymphocytes, leukocytes, endothelial cells, and dendritic cells [333,334,335,336] | 20-hydroxy-LTB4, 12R-HETE, Leukotriene B4 [337,338] | oxazolone [339] | ||||
| BLT2 | Keratinocytes [340,341], spleen, liver, ovary, leukocytes, and atherosclerotic lesions [333,338] | 20-hydroxy-LTB4, 12R-HETE, Leukotriene B4 [337,338] | oxazolone [339] | ||||
| Phospholipid receptor | GPCR | Phospholipid | |||||
| PAFR | Keratinocytes [342], leukocytes, and granulocytes [343,344] | platelet-activating factor, methylcarbamyl PAF [345] | N/A | ||||
| Opoid receptors | GPCRs | Opioids | |||||
| OPRM | Keratinocytes [346], dermal fibroblasts [347], CNS, T/B lymphocytes, CD4+, monocytes, macrophages, and neutrophils [348] | β-endorphin, enkephalin, dynorphin [349,350] | morphine [351] | ||||
| OPRK | Keratinocytes [352], dermal fibroblasts [353], CNS, and immune cells [353,354,355] | dynorphin, β-endorphin, enkephalin, neoendorphin [349,356] | nor-binaltorphimine [357] | ||||
| Cannabinoid receptors | GPCRs | Cannabinoids | |||||
| CB1 | Keratinocytes [358], CNS, and peripheral neurons [359] | anandamide, 2-arachidonoylglycerol [360,361] | Rimonabant [362] | ||||
| CB2 | Keratinocytes [358], spleen, tonsils, bone marrow, and peripheral blood leukocytes [189] | anandamide, 2-arachidonoylglycerol [360,361] | N/A | ||||
| Interleukin receptors | non-GPCRs | Interleukins | |||||
| IL2R | CD8 + T cells, and natural killer cells [363] | JAK1/3 | IL-2 | Aldesleukin [215] | |||
| IL4R | Keratinocytes [364], activated T-cells, and hematopoietic immune cells [365] | JAK1/2/3 | IL-4 | N/A | |||
| IL13R | B-cells, T-cells, basophils, eosinophils, mast cells and endothelial cells of heart, liver, skeletal muscle and ovary [366] | JAK1 | IL-13 | N/A | |||
| IL31R | CD14-, CD56- positive blood cells, macrophages, keratinocytes, dorsal root ganglia neurons, lung epithelial cells [241,367,368,369] | JAK1/2 | IL-31 | N/A | |||
| OSMR | keratinocytes, neural cells, fibroblast, and epithelial cells [367,370] | JAK1/2 | Oncostatin M | N/A | |||
| TSLPR | keratinocytes [371], B-cells, T-cells, and macrophage cell lines [372] | JAK1/2 | Thymic stromal lymphopoietin (TSLP) | N/A | |||
| Toll-like receptors | non-GPCRs | various | |||||
| TLR3 | keratinocytes [373], dermal langerhans cells [374], dendritic cells, astrocytes, glia and neurons; especially in placenta and pancreas [375] | TRIF | ds RNA | Polyinosinic:polycytidylic acid [105] | |||
| TLR4 | Keratinocytes [373], monocytes, macrophages, dendritic cells, and T-cells [376,377] | MyD88/TRIF | LPS | paclitaxel [378] | |||
| TLR7 | Plasmacytoid dendritic cells, and B cells [379] | MyD88 | ss RNA | Imiquimod [288] | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kahremany, S.; Hofmann, L.; Gruzman, A.; Cohen, G. Advances in Understanding the Initial Steps of Pruritoceptive Itch: How the Itch Hits the Switch. Int. J. Mol. Sci. 2020, 21, 4883. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144883
Kahremany S, Hofmann L, Gruzman A, Cohen G. Advances in Understanding the Initial Steps of Pruritoceptive Itch: How the Itch Hits the Switch. International Journal of Molecular Sciences. 2020; 21(14):4883. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144883
Chicago/Turabian StyleKahremany, Shirin, Lukas Hofmann, Arie Gruzman, and Guy Cohen. 2020. "Advances in Understanding the Initial Steps of Pruritoceptive Itch: How the Itch Hits the Switch" International Journal of Molecular Sciences 21, no. 14: 4883. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144883