The Potential of Insects as Alternative Sources of Chitin: An Overview on the Chemical Method of Extraction from Various Sources

 ,
,
Abstract
:1. Introduction
2. Result and Discussion
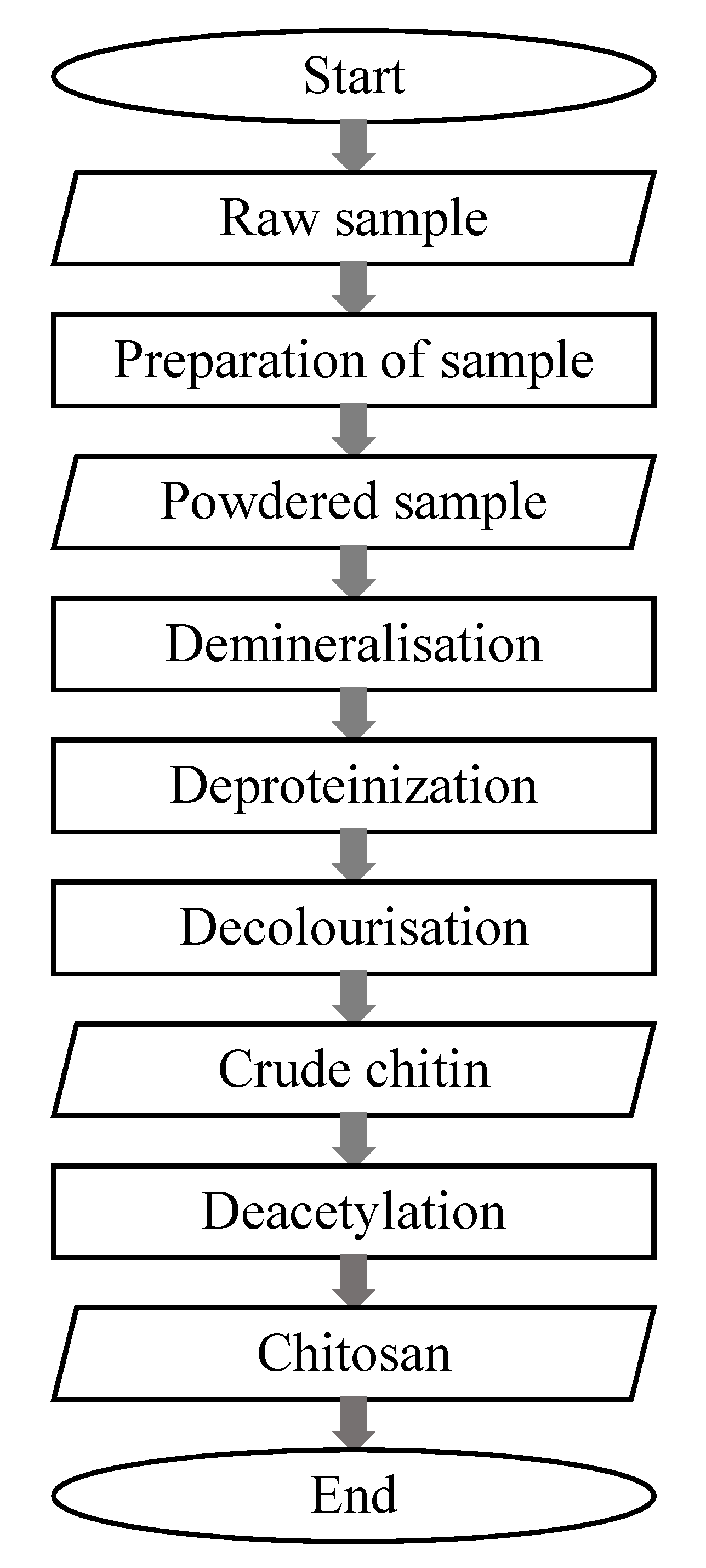
2.1. Commercial Preparation of Chitin
2.2. Chemical Extraction of Chitin and Chitosan from Insects as an Alternative to Commercial Chitin from Crustaceans
2.3. Chemical Extraction of Chitin and Chitosan from Crustaceans
2.4. Chemical Extraction of Chitin and Chitosan from Other Resources
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CaCO3 | calcium carbonate |
| HCl | hydrochloric acid |
| NaOH | sodium hydroxide |
| FTIR | Fourier-transform infrared |
| 13CP/MAS-NMR | 13C cross-polarization magic-angle-spinning nuclear magnetic resonance |
| DDA | degree of deacetylation |
| H NMR | hydrogen nuclear magnetic resonance |
| IR | infrared |
| XRD | X-ray diffraction |
| SEM | scanning electron microscope |
| TGA | thermogravimetric analysis |
| DTGmax | maximal decomposition rate |
References
- Brine, C.J.; Austin, P.R. Chitin variability with species and method of preparation. Comp. Biochem. Physiol. 1981, 69, 283–286. [Google Scholar] [CrossRef]
- Braconnot, H. Sur la nature des champignons. Ann. Chim. Phys. 1811, 79, 265–304. [Google Scholar]
- Hoffman, A. How LSD Originated. J. Phychoactive Drugs 1979, 11, 53–60. [Google Scholar] [CrossRef]
- Hagenbach, D.A.; Werthmuller, L.; Grof, S. Mystic Chemist: The Life of Albert Hoffman and His Discovery of LSD; Synergetic Press: Santa Fe, NM, USA, 2013. [Google Scholar]
- Sashiwa, H.; Aiba, S.-i. Chemically modified chitin and chitosan as biomaterials. Prog. Polym. Sci. 2004, 29, 887–908. [Google Scholar] [CrossRef]
- Pilai, K.S.; Paul, W.; Sharma, C.P. Chitin and chitosan polymers: Chemistry, solublity and fiber formation. Prog. Polym. Sci. 2009, 34, 641–678. [Google Scholar] [CrossRef]
- Robert, G.A. Chitin Chemistry; Macmillan Press Ltd.: London, UK, 1992. [Google Scholar]
- Muzzarelli, R.A. Chitin nanostructure in living organisms. In Chitin: Formation and Diagenesis; Gupta, N., Ed.; Springer: Dordrecht, The Netherland, 2011; pp. 1–34. [Google Scholar]
- Domard, A.; Rinaudo, M. Preparation and characterization of fully deacetylated chitosan. Int. J. Biol. Macromol. 1983, 5, 49–52. [Google Scholar] [CrossRef]
- Zhang, M.; Haga, A.; Sekiguchi, H.; Hirano, S. Structure of insect chitin isolated from beetle larva cuticle and silkworm (Bombyx mori) pupa exuvia. Int. J. Biol. Macromol. 2000, 40, 99–105. [Google Scholar] [CrossRef]
- Liu, S.; Sun, J.; Yu, L.; Zhang, C.; Bi, J.; Zhu, F.; Qu, M.; Jiang, C.; Yang, Q. Extraction and characterization of chitin from the beetle Holotrichia parallela motschulsky. Molecules 2012, 17, 4604–4611. [Google Scholar] [CrossRef] [PubMed]
- No, H.K.; Lee, M.Y. Isolation of chitin from crab shells waste. J. Korean Soc. Food Nutr. 1995, 24, 105–113. [Google Scholar]
- Tolaimate, A.; Desbrieres, J.; Rhazi, M.; Alagui, A. Contribution to the preparation of chitins and chitosans with controlled physico-chemical properties. Polymer 2003, 44, 7939–7952. [Google Scholar] [CrossRef]
- Azuma, K.; Ifuku, S.; Osaki, T.; Okamoto, Y.; Minami, S. Preparation and biomedical applications of chitin and chitosan nanofibers. J. Biomed. Nanotechnol. 2014, 10, 2891–2920. [Google Scholar] [CrossRef] [PubMed]
- Andrew, C.A.; Wan, A.C.; Tai, B.C. Chitin—A promising biomaterial for tissue engineering and stem cell technologies. Biotechnol. Adv. 2013, 31, 1776–1785. [Google Scholar]
- Jayakumar, R.; Prabaharan, M.; Nair, S.V.; Tamura, H. Novel chitin and chitosan nanofibers in biomedical applications. Biotechnol. Adv. 2010, 28, 142–150. [Google Scholar] [CrossRef]
- No, H.K.; Meyers, S.P.; Lee, K.S. Isolation and characterization of chitin from crayfish shell waste. J. Agric. Food Chem. 1989, 37, 575–579. [Google Scholar] [CrossRef]
- Shah, P.; Jogani, V.; Mishra, P.; Mishra, A.K.; Bagchi, T.; Misra, A. Modulation of gancioclovir intestinal absorption in presence of absorption enhancers. J. Pharm. Sci. 2007, 96, 2710–2722. [Google Scholar] [CrossRef]
- Chen, C.S.; Liau, W.Y.; Tsai, G.J. Antibacterial effects of N-sulfonated and N-sulfobenzoyl chitosan and application to oyster preservation. J. Food Prot. 1998, 64, 1124–1128. [Google Scholar] [CrossRef]
- Mezzana, P. Clinical efficacy of a new chitin nanofibrils based gel in wound healing. Acta Chir. Plast. 2009, 50, 81–84. [Google Scholar]
- Morganti, P. Chitin-nanofibrils in skin treatment. J. Appl. Cosmetol. 2009, 27, 251–270. [Google Scholar]
- Francis, R.J.; Prestwich, G.D.; Hunt, G. System and Method of Delivering a Hyaluronic Acid Composition and a Copper Composition for Treatment of Dermatologic Conditions. US Patent 2015,081,304 A1, 4 June 2015. [Google Scholar]
- Majeti, N.V.; Kumar, R. A review of chitin and chitosan applications. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar]
- Puvvada, Y.S.; Vankayalapati, S.; Sukhavasi, S. Extraction of chitin from chitosan from exoskeleton of shrimp for application in the pharmaceutical industry. Int. Curr. Pharm. J. 2012, 1, 258–263. [Google Scholar] [CrossRef] [Green Version]
- Schipper, N.G.M.; Varum, K.M.; Artursson, P. Chitosans as absorption enhancers for poorly absorbable drugs: 1. Influence of molecular weight and degree of deacetylation on drug transport across human intestinal epithelial (Caco-2) cells. Pharm. Res. 1996, 13, 1686–1692. [Google Scholar] [CrossRef] [PubMed]
- Schipper, N.G.M.; Olsson, S.; Hoogstraate, J.A.; deBoer, A.G.; Varum, K.M.; Artursson, P. Chitosan as absorption enhancers for poorly absorbable drugs: 2. Mechanism of absorption enhancement. Pharm. Res. 1997, 14, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Trapani, A.; Lopedota, A.; Franco, M.; Cioffi, N.; Ieva, E.; Garcia-Fuentes, M.; Alonso, M.J. A comparative study of chitosan and chitosan/cyclodextrin nanoparticles as potential carriers for the oral delivery of small peptides. Eur. J. Pharm. Biopharmacy 2010, 75, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Kuria, K. Controlled fractionation of the polysaccharide chitin. Prog. Polym. Sci. 2001, 26, 1921–1971. [Google Scholar] [CrossRef]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–623. [Google Scholar] [CrossRef]
- McGavin, G.C. Insects, Spider and Other Terrestrial Arthropods; Dorling Kindersley Limited: London, UK, 2000. [Google Scholar]
- Richards, L. Integument of Arthropods; University of Minnesota Press: Minneapolis, MN, USA, 1951. [Google Scholar]
- Pavlichko, J.P. Polymer interactions to enhance the function of hyaluronic acid. Drug Cosmet. Ind. 1990, 147, 12–24. [Google Scholar]
- De Smedt, S.C.; Demeester, J.; Hennink, W.E. Cationic polymer based gene delivery systems. Pharmacol. Res. 2000, 17, 113–126. [Google Scholar] [CrossRef]
- Fan, Y.; Saito, T.; Isogai, A. Individual chitin nano-whiskers prepared from partially deacetylated alpha-chitin by fibril surface cationization. Carbohydr. Polym. 2010, 79, 1046–1051. [Google Scholar] [CrossRef]
- Chandrkrachang, S. The applications of chitin in agriculture in Thailand. Adv. Chitin Sci. 2002, 5, 458–462. [Google Scholar]
- Nahar, S.J.; Kazuhiko, S.; Haque, S.M. Effect of polysaccharides including elicitors on organogenesis in protocorm-like body (PLB) of Cymbidium insigne in vitro. J. Agric. Sci. Technol. 2012, 2, 1029–1033. [Google Scholar]
- Ohta, K.; Tanguchi, A.; Konishi, N.; Hosoki, T. Chitosan treatment affects plant growth and flower quality in Eustoma grandiflorum. HortScience 1999, 34, 233–234. [Google Scholar] [CrossRef] [Green Version]
- Knorr, D. Functional properties of chitin and chitosan. J. Food Sci. 1982, 47, 593–595. [Google Scholar] [CrossRef]
- Tzoumaki, M.V.; Moschakis, T.; Kiosseoglou, V.; Biliaderis, C.G. Oil-in-water emulsions stabilized by chitin nanocrystal particles. Food Hydrocoll. 2011, 25, 1521–1529. [Google Scholar] [CrossRef]
- Tzoumaki, M.V.; Moschakis, T.; Scholten, E.; Biliaderis, C.G. In vitro lipid digestion of chitin nanocrystal stabilized o/w emulsions. Food Funct. 2013, 4, 121–129. [Google Scholar] [CrossRef]
- Cho, Y.-W.; Cho, Y.-N.; Chung, S.-H.; Yoo, G.; Ko, S.W. Water-soluble chitin as a wound healing accelerator. Biomaterials 1999, 20, 2139–2145. [Google Scholar] [CrossRef]
- Madhumathi, K.; Kumar, P.T.S.; Abhilash, S.; Sreeja, V.; Tamura, H.; Manzoor, K.; Nair, S.V.; Jayakumar, R. Development of novel chitin/nanosilver composite scaffolds for wound dressing applications. J. Mater. Sci. Mater. Med. 2010, 21, 807–813. [Google Scholar] [CrossRef]
- Zia, K.M.; Zuber, M.; Bhatti, I.A.; Barikani, M.; Sheikh, M.A. Evaluation of biocompatibility and mechanical behavior of polyurethane elastomers based on chitin/1,4-butane diol blends. Int. J. Biol. Macromol. 2009, 44, 18–22. [Google Scholar] [CrossRef]
- Park, B.K.; Kim, M.-M. Applications of chitin and its derivatives in biological medicine. Int. J. Mol. Sci. 2010, 11, 5152–5164. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, M.A.; Rodriguez, A.T.; Alfonso, L.; Peniche, C. Chitin and its derivatives as biopolymers with potential agricultural applications. Biotechnol. Apl. 2010, 27, 270–276. [Google Scholar]
- Illum, L. Chitosan and its use as a pharmaceutical excipient. Pharm. Res. 1998, 15, 1326–1331. [Google Scholar] [CrossRef]
- Yoshinori, K.; Hiraku, O.; Yoshiharu, M. Application of chitin and chitosan derivatives in the pharmaceutical field. Curr. Pharm. Biotechnol. 2003, 4, 303–309. [Google Scholar]
- Morganti, P.; Morganti, G.; Morganti, A. Transforming nanostructured chitin from crustacean waste into beneficial health products: A must for our society. Nanotechnol. Sci. Appl. 2011, 4, 123–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morganti, P.; Fabrizi, G.; Palombo, M.; Ruocco, E.; Cardillo, A.; Morganti, G. Chitin-nanofibrils: A new active cosmetic carrier. J. Appl. Cosmetol. 2008, 26, 113–128. [Google Scholar]
- Jeon, Y.-J.; Shahidi, F.; Kim, S.-K. Preparation of chitin and chitosan oligomers and their applications in physiological functional foods. Food Rev. Int. 2012, 16, 159–176. [Google Scholar] [CrossRef]
- Manni, L.; Ghorbel-Bellaaj, O.; Jellouli, K.; Younes, I.; Nasri, M. Extraction and characterization of chitin, chitosan and protein hydrolysates prepared from shrimp waste by treatment with crude protease from Bacillus cereus SV1. Appl. Biochem. Biotechnol. 2010, 162, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Dhillon, G.S. Recent trends in biological extraction of chitin from marine shell wastes: A review. Crit. Rev. Biotechnol. 2015, 35, 44–61. [Google Scholar] [CrossRef]
- Synowiecki, J.; Al-Khateeb, N.A.A.Q. The recovery of protein hydrolysate during enzymatic isolation of chitin from shrimp Crangon crangon processing discards. Food Chem. 2000, 68, 147–152. [Google Scholar] [CrossRef]
- Younes, I.; Hajji, S.; Franchet, V.; Rinaudo, M.; Jellouli, K.; Nasri, M. Chirin extraction from shrimp shell using enzymatic treatment. Antitumor, antioxidant and antimocrobial activities of chitosan. Int. J. Biol. Macromol. 2014, 69, 489–498. [Google Scholar] [CrossRef]
- Majtan, J.; Bilikova, K.; Markovic, O.; Grof, J.; Kogan, G.; Simuth, J. Isolation and characterization of chitin from bumblebee (Bombus terrestris). Int. J. Biol. Macromol. 2007, 40, 237–241. [Google Scholar] [CrossRef]
- Chang, K.L.; Tsai, G. Response surface optimization and kinetics of isolating chitin from pink shrimp (Solenocera melantho) shell waste. J. Agric. Food Chem. 1997, 45, 1900–1904. [Google Scholar] [CrossRef]
- Tokatli, K.; Demirdoven, A. Optimization of chitin and chitosan production from shrimp wastes and characterization. J. Food Process. Preserv. 2017, 1–13. [Google Scholar] [CrossRef]
- Duong, N.T.H.; Nghia, N.D. Kinetics and optimization of the deproteinization by pepsin in chitin extraction from white shrimp shell. J. Chitin Chitosan Sci. 2014, 2, 1–8. [Google Scholar] [CrossRef]
- Guo, N.; Sun, J.; Zhang, Z.; Mao, X. Recovery of chitin and protein from shrimp head waste by endogenous enzyme autolysis and fermentation. J. Ocean Univ. China 2019, 18, 719–726. [Google Scholar] [CrossRef]
- Garcia, Y.H.; Troncoso-Rojas, R.; Tiznado-Hernandez, M.E.; Baez-Flores, M.E.; Carvajal-Mill, E.; Rason-Chu, A.; Lizardi-Mendoza, J.; Martinez-Robinson, K.G. Enzymatic treatments as alternative to produce chitin fragments of low molecular weight from Alternaria alternata. J. Appl. Polym. Sci. 2019, 136, 47339. [Google Scholar] [CrossRef]
- Hackman, R.H. Studies on chitin I. Enzymatic Degradation of chitin and chitin esters. Aust. J. Biol. Sci. 1954, 7, 168–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younes, I.; Ghorbel-Bellaaj, O.; Nasri, R.; Chaabouni, M.; Rinaudo, M.; Moncef, N. Chitin and chitosan preparation from shrimp shells using optimized enzymatic deproteinization. Process Biochem. 2012, 47, 2032–2039. [Google Scholar] [CrossRef]
- Arbia, W.; Arbia, L.; Adour, L.; Amrane, A. Chitin etraction from crustacean shells using biological methods- a review. Food Technol. Biotecnol. 2013, 51, 12–25. [Google Scholar]
- Machalowski, T.; Wysokowski, M.; Tsurkan, M.V.; Galli, R.; Schimpf, C.; Rafaja, D.; Brendler, E.; Viehweger, C.; Zoltowska-Aksamitowska, S.; Petrenko, I.; et al. Spider chitin: An ultrafast microwave-assisted method for chitin isolation from Caribena versicolor spider molt cuticle. Molecules 2019, 24, 3736. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Benjakul, S.; Prodpran, T. Ultrasound-assisted extraction of chitosan from squid pen: Molecular characterization and fat binding capacity. J. Food Sci. 2019, 84, 224–234. [Google Scholar] [CrossRef]
- Kardas, I.; Struszczyk, M.H.; Kucharska, M.; van den Broek, L.A.M.; van Dam, J.E.G.; Ciechanska, D. Chitin and chitosan as functional biopolymers for industrial applications. Eur. Polysacch. Netw. Excell. 2012, 329–373. [Google Scholar] [CrossRef]
- No, H.K.; Meyers, S.P. Preparation and characterization of chitin and chitosan—A review. J. Aquat. Food Prod. Technol. 1995, 4, 27–52. [Google Scholar] [CrossRef]
- Hajji, S.; Younes, I.; Ghorbel-Bellaj, O.; Hajji, R.; Rinaudo, M.; Nasri, M.; Jellouli, K. Structural differences between chitin and chitosan extracted from three different marine sources. Int. J. Biol. Macromol. 2014, 65, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Younes, I.; Ghorbel-Bellaaj, O.; Chaabouni, M.; Rinaudo, M.; Souard, F.; Vanhaverbeke, C.; Jellouli, K.; Nasri, M. Use of a fractional factorial design to study the effects of experimental factors on the chitin deacetylation. Int. J. Biol. Macromol. 2014, 70, 385–390. [Google Scholar] [CrossRef]
- Jalal, A.F.; Risheed, C.M.; Ibrahim, B.M. Optimization of chitin extraction from chicken feet. J. Anal. Bioanal. Tech. 2012, 3, 1–5. [Google Scholar] [CrossRef]
- Peniston, Q.P.; Johnson, E.L. Method for Treating an Aqueous Medium with Chitosan and Derivatives of Chitin to Remove Impurities. U.S. Patent 3,533,940, 13 October 1970. [Google Scholar]
- Peniston, Q.P.; Gortner, R.A.; Sandstorm, W.M. Process for Depolymerization of Chitosan. U.S. Patent 3,922,260, 25 November 1975. [Google Scholar]
- Peniston, Q.P.; Johnson, E.L. Method of Recovering Chitosan and Other by-Products from Shellfish Waste and the Like. US Patent 3,862,122, 21 January 1975. [Google Scholar]
- Peniston, Q.P.; Johnson, L.E. Process for Demineralization of Crustacea Shells. US Patent 4,066,735, 3 January 1978. [Google Scholar]
- Peniston, Q.P.; Johnson, E.L. Process for the Manufacture of Chitosan. US Patent 4,195,175, 25 March 1980. [Google Scholar]
- Chaput, C.; Chenite, A. Injectable in Situ Self-Performing Mineral-Polymer Hybrid Composition and Uses Thereof. US Patent 8,747,899, 10 June 2014. [Google Scholar]
- McCarthy, S.; McGrath, B.; Winata, E. Absorbable Tissue Dressing Assemblies, Systems, and Methods Formed from Hydrophilic Polymer Sponge Structures such as Chitosan. US Patent 8,269,058 B2, 18 September 2012. [Google Scholar]
- McCarthy, S.J.; Gregory, K.W.; Wiesmann, W.P.; Campbell, T.D. Wound Dressing and Method for Controlling Severe, Life-Threatening Bleeding. US Patent 8,668,924 B2, 11 March 2014. [Google Scholar]
- Lucchesi, L.; Xie, H. Wound Dressing Devices and Methods. US Patent 9,205,170 B2, 8 December 2015. [Google Scholar]
- Finkielsztein, S.; Vournakis, J.N. Hemostatic Compositions and Therapeutic Regimens. US Patent 9,139,664 B2, 22 September 2015. [Google Scholar]
- D’huart, J.B.; Dallas, C. Cactaceae-Based Formulation Having the Property of Fixing Fats, and Method for Obtaining Same. US Patent 2004,0,126,444 A1, 1 July 2004. [Google Scholar]
- Johnson, E.L.; Nichols, E.J. High Tap Density Chitosan, and Methods of Production. US Patent 6,130,321, 10 October 2000. [Google Scholar]
- Yadav, M.; Goswami, P.; Paritosh, K.; Kumar, M.; Pareek, N.; Vivekanand, V. Seafood waste: A source for preparation of commercially employable chitin/chitosan materials. Bioresour. Bioprocess. 2019, 6, 1–20. [Google Scholar] [CrossRef]
- Fernando, L.A.T.; Poblete, M.R.S.; Ongkiko, A.G.M.; Diaz, L.J.L. Chitin extraction and synthesis of chitin-based polymer films from Philiphine blue swimming crab (Portumus pelagicus) shells. Procedia Chem. 2016, 19, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Huet, G.; Hadad, C.; Husson, E.; Laclef, S.; Lambertyn, V.; Farias, M.A.; Jamali, A.; Courty, M.; Alayoubi, R.; Gosselin, I.; et al. Straightforward extraction and selective bioconversion of high purity chitin from Bombyx eri larva: Toward an integrated insect biorefinery. Carbohydr. Polym. 2020, 228, 115382. [Google Scholar] [CrossRef]
- Sajomsang, W.; Gonil, P. Preparation and characterization of alpha-chitin from cicada sloughs. Mater. Sci. Eng. C 2010, 30, 357–363. [Google Scholar] [CrossRef]
- Song, C.; Yu, H.; Zhang, M.; Yang, Y.; Zhang, G. Physiochemical properties and antioxidant activity of chitosan from the blowfly Chrysomya megacephala larvae. Int. J. Biol. Macromol. 2013, 60, 347–354. [Google Scholar] [CrossRef]
- Paulino, A.T.; Simionato, J.I.; Garcia, J.C.; Nozaki, J. Characterization of chitosan and chitin produced from silkworm crysalides. Carbohydr. Polym. 2006, 64, 98–103. [Google Scholar] [CrossRef]
- Kaya, M.; Baran, T.; Erdogan, S.; Mentes, A.; Ozusaglam, M.A.; Cakmak, Y.S. Physicochemical comparison of chitin and chitosan obtained from larvae and adult colorado potato beetle (Leptinotarsa decemlineata). Mater. Sci. Eng. C 2014, 45, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Baublys, V.; Can, E.; Satkauskiene, I.; Bitim, B.; Tubelyte, V.; Baran, T. Comparison of physicochemical properties of chitins isolated from an insect (Melolontha melolontha) and a crustacean species (Oniscus asellus). Zoomorphology 2014, 133, 285–293. [Google Scholar] [CrossRef]
- Kaya, M.; Baran, T.; Karaarslan, M. A new method for fast chitin extraction from shells of crab, crayfish and shrimp. Nat. Prod. Res. 2015, 29, 1477–1480. [Google Scholar] [CrossRef]
- Marei, N.H.; Samiee, E.A.E.; Salah, T.; Saad, G.R.; Elwahy, A.H.M. Isolation and characterization of chitosan from different local insects in Egypt. Int. J. Biol. Macromol. 2016, 82, 871–877. [Google Scholar] [CrossRef]
- Kim, M.-W.; Song, Y.-S.; Han, Y.-S.; Jo, Y.-H.; Choi, M.-H.; Park, Y.-K.; Kang, S.-H.; Kim, S.-A.; Choi, C.; Jung, W.-J. Production of chitin and chitosan from the exoskeleton of adult two-spotted field crickets (Gryllus bimaculatus). Entomol. Res. 2017, 47, 279–285. [Google Scholar] [CrossRef]
- Ibitoye, E.B.; Lokman, I.H.; Hezmee, M.N.; Goh, Y.M.; Zuki, A.B.; Jimoh, A.A. Extraction and physcicochemical characterization of chitin and chitosan isolated from house cricket. Biomed. Mater. 2018, 13, 025009. [Google Scholar] [CrossRef] [Green Version]
- Shin, C.-S.; Kim, D.-Y.; Shin, W.-S. Characterization of chitosan extracted from mealworm beetle (Tenebrio molitor, Zophobas morio) and rhinoceros beetle (Allomyrina dichotoma) and their antibacterial activities. Int. J. Biol. Macromol. 2019, 125, 72–77. [Google Scholar] [CrossRef]
- Kaya, M.; Baublys, V.; Satkauskiene, I.; Akyuz, B.; Bulut, E.; Tubelyte, V. First chitin extraction from Plumatella repens (Bryozoa) with comparison to chitins of insects and fungal origin. Int. J. Biol. Macromol. 2015, 79, 126–132. [Google Scholar] [CrossRef]
- Ifuku, S. Chitin and chitosan nanofibers: Preparation and chemical modifications. Molecules 2014, 19, 18367–18380. [Google Scholar] [CrossRef]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdou, E.S.; Nagy, K.S.A.; Elsabee, M.Z. Extraction and characterization of chitin and chitosan from local sources. Bioresour. Technol. 2008, 99, 1359–1367. [Google Scholar] [CrossRef]
- Knorr, D. Use of chitinous polymers in food: A challenge for food research and development. Food Technol. 1984, 38, 85–89, 92–97. [Google Scholar]
- Percot, A.; Viton, C.; Domard, A. Optimization of chitin extraction from shrimp shells. Biomacromolecules 2003, 4, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Karaarslan, M.; Baran, T.; Can, E.; Ekemen, G.; Duman, F. The quick extraction of chitin from an epizoic crustacean species (Chelonibia patula). Nat. Prod. Res. 2014, 28, 2186–2190. [Google Scholar] [CrossRef]
- Mohan, K.; Ravichandran, S.; Muralisankar, T.; Uthayakumar, V.; Chandiransekar, R.; Rajeevgandhi, C.; Rajan, D.K.; Seedevi, P. Extraction and charcterization of chitin from sea snail Conus inscriptus (Reeve, 1843). Int. J. Biol. Macrmolecules 2019, 126, 555–560. [Google Scholar] [CrossRef]
- Charoevuttitham, P.; Shi, J.; Mittal, G.S. Chitin extraction from black tiger shrimp (Panaeus monodon) waste using organic acids. Sep. Sci. Technol. 2006, 41, 1135–1153. [Google Scholar] [CrossRef]
- Limam, Z.; Selmi, S.; Sadok, S.; el Abed, A. Extraction and characterization of chitin and chitosan from crustacean by-products: Biological and physicochemical properties. Afr. J. Biotechnol. 2011, 10, 640–647. [Google Scholar]
- de Moura, C.M.; de Moura, J.M.; Soares, N.M.; Pinto, L.A.d. Evaluation of molar weight and deacetylation degree of chitosan during chitin deacetylation reaction: Used to produce biofilm. Chem. Eng. Process. Process Intesification 2011, 50, 351–355. [Google Scholar] [CrossRef]
- Ploydee, E.; Chaiyanan, S. Production of high viscosity chitosan from biological purified chitin isolated by microbial fermentation and deproteinization. Int. J. Polym. Sci. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shimahara, K.; Takiguchi, Y. Preparation of crustacean chitin. Methods Enzymol. 1988, 161, 417–423. [Google Scholar]
- Kjartansson, G.T.; Zivanovic, S.; Kristbergsson, K.; Weiss, J. Sonication-assisted extraction of chitin from shells of fresh water prawns (Macrobrachium rosenbergii). J. Agric. Food Chem. 2006, 54, 3317–3323. [Google Scholar] [CrossRef]
- Mohammed, M.H.; Williams, P.A.; Tverezovskaya, O. Extraction of chitin from prawn shells and conversion to low molecular mass chitosan. Food Hydrocoll. 2013, 31, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Ifuku, S.; Nogi, M.; Abe, K.; Yoshioka, M.; Morimoto, M.; Saimoto, H.; Yano, H. Preparation of chitin nanofibers with a uniform width as alpha-chitin from crab shells. Biomacromolecules 2009, 10, 1584–1588. [Google Scholar] [CrossRef] [PubMed]
- Fadlaoui, S.; el Asri, O.; Mohammed, L.; Sihame, A.; Omar, A.; Melhaoui, M. Isolation and characterization of chitin from shells of the freshwater crab Potamon algeriense. Prog. Chem. Appl. Chitin Its Deriv. 2019, 24, 23–35. [Google Scholar] [CrossRef]
- Pădurețu, C.-C.; Isopescu, R.; Rău, I.; Apetroaei, M.R.; Schröder, V. Influence of the parameters of chitin deacetylation process on the chitosan obtained from crab shell waste. Korean J. Chem. Eng. 2019, 36, 1890–1899. [Google Scholar] [CrossRef]
- Abdulkarim, A.; Isa, M.T.; Abdulsalam, S.; Muhammad, A.J.; Ameh, A.O. Extraction and characterization of chitin and chitosan from mussel shell. Civ. Environ. Res. 2013, 3, 108–114. [Google Scholar]
- Kaya, M.; Sargin, I.; Tozak, K.O.; Baran, T.; Erdogan, S.; Sezen, G. Chitin extraction and characterization from Daphnia magna resting eggs. Int. J. Biol. Macromol. 2013, 61, 459–464. [Google Scholar] [CrossRef]
- Kaya, M.; Seyyar, O.; Baran, T.; Erdogan, S.; Kar, M. A physicochemical characterization of fully acetylated chitin structure isolated from two spider species: With new surface methodology. Int. J. Biol. Macromol. 2014, 65, 553–558. [Google Scholar] [CrossRef]
- Ifuku, S.; Nomura, R.; Morimoto, M.; Saimoto, H. Preparation of chitin nanofibers from mushrooms. Materials 2011, 4, 1417–1425. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Rath, P.K. Extraction and characterization of chitin and chitosan from (Labeo rohit) fish scales. Procedia Mater. Sci. 2014, 6, 482–489. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Rath, P.; Kumar, A.S.H.; Tiwari, T.N. Extraction and characterization of chitin and chitosan from fishery waste by chemical method. Environ. Technol. Innov. 2015, 3, 77–85. [Google Scholar] [CrossRef]
- Soon, C.Y.; Tee, Y.B.; Tan, C.H.; Rosnita, A.T.; Khalina, A. Extraction and physicochemical characterization of chitin and chitosan from Zophobas morio larvae in varying sodium hydroxide concentration. Int. J. Biol. Macromol. 2018, 108, 135–142. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Insects | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sources | Demineralization | Deproteinization | Yield | References | ||||||||||
| Scientific Name | Common Name | Type of Acid | Concentration | Temperature (°C) | Solution-to-Solid Ratio (mL/g) | Duration | Type of Alkali | Concentration | Temperature (°C) | Solution-to-Solid Ratio (mL/g) | Duration | |||
| N/A | Beetle larvae cuticle | HCl | 1 N | 100 | N/A | 20 min | NaOH | 1 N | 80 | N/A | 24–36 h | a 15%–20% | [10] | |
| Reflux with 0.4% of Na2CO3, 20 h | ||||||||||||||
| Bombus terrestris | Bumblebee | HCl | 1 M | 100 | N/A | 20 min | NaOH | 1 M | 85 | N/A | 24 h | a 6.89% | [55] | |
| Periostracum Cicadae | Cicada slough | HCl | 1 N | 100 | N/A | 20 min | NaOH | 1 N | 80 | N/A | 36 h | a 36.6% | [86] | |
| Refluxed with NaCO3,1 2 h | ||||||||||||||
| Holotrichia parallela | Dark black chafer beetle | HCl | 1 M | 100 | 50 | 30 min | NaOH | 1 M | 80 | 50 | 24 h | a 15% | [11] | |
| Chyrysomya megacephala | Oriental latrine fly | Oxalic acid | N/A | N/A | N/A | 3 h | NaOH | 1 M | 95 | N/A | 6 h | b 26.2 | [87] | |
| Leptinotarsa decemlineata | Potato beetle | HCl | 2 M | 65–75 | N/A | 2 h | NaOH | 2 M | 80–90 | N/A | 16 h | a 20% b 72% | [89] | |
| Leptinotarsa decemlineata | Larvae | HCl | 2 M | 65–75 | N/A | 2 h | NaOH | 2 M | 80–90 | N/A | 16 h | a 7% b 67% | [89] | |
| Melolontha melolontha | Cockchafer beetle | HCl | 4 M | 75 | N/A | 2 h | NaOH | 4 M | 150 | N/A | 18 h | a 13%–14% | [90] | |
| Palomena prasina | Green shield bug | HCl | 2 M | 100 | N/A | 2 h | NaOH | 2 M | 140 | N/A | 20 h | a 10.8% | [96] | |
| Schistocerca gregaria | Desert locust | HCl | 1 M | RT | 15 | N/A | NaOH | 1 M | 100 | N/A | 8 h | a 12.2% | [92] | |
| Apis mellifera | Western honeybee | HCl | 1 M | RT | 15 | N/A | NaOH | 1 M | 100 | N/A | 8 h | a 2.5% | [92] | |
| Calosoma rugosa | Beetles | HCl | 1 M | RT | 15 | N/A | NaOH | 1 M | 100 | N/A | 8 h | a 5% | [92] | |
| Gryllus bimaculatus | Crickets | HCl | 2 N | 21 | N/A | 3 h | NaOH | 1.25 N | 95 | N/A | 3 h | a 2.35% b 1.79% | [93] | |
| Brachytrupes portentosus | House cricket | Oxalic acid | N/A | RT | 20 | 3 h | NaOH | 1 M | 95 | 20 | 6 h | a 4.3%–7.1% b 2.4%–5.8% | [94] | |
| Tenebrio molitor | Mealworm beetle (larvae) | HCl | 7% | 25 | N/A | 24 h | NaOH | 10% | N/A | 80 | 24 h | a 4.6% b 80% | [95] | |
| Tenebrio molitor | Mealworm beetle (adult) | HCl | 7% | 25 | N/A | 24 h | NaOH | 10% | N/A | 80 | 24 h | a 8.4% b 78.33% | [95] | |
| Zophobas morio | Superwrm | HCl | 7% | 25 | N/A | 24 h | NaOH | 10% | N/A | 80 | 24 h | a 3.9% b 83.33% | [95] | |
| Allomyrina dichotoma | Rhinoceros beetle (larvae) | HCl | 7% | 25 | N/A | 24 h | NaOH | 10% | N/A | 80 | 24 h | a 10.5% b 83.37% | [95] | |
| Allomyrina dichotoma | Rhinoceros beetle (pupa) | HCl | 7% | 25 | N/A | 24 h | NaOH | 10% | N/A | 80 | 24 h | a 12.7% b 83.37% | [95] | |
| Allomyrina dichotoma | Rhinoceros beetle (adult) | HCl | 7% | 25 | N/A | 24 h | NaOH | 10% | N/A | 80 | 24 h | a 14.2% b 75% | [95] | |
| Bombyx mori | Silkworm | HCl | 1 N | 100 | N/A | 20 min | NaOH | 1 N | 80 | N/A | 24–36 h | a 15%–20% b 70%–80% | [10] | |
| Reflux with 0.4% of Na2CO3, 20h | ||||||||||||||
| Bombyx mori | Silkworm | HCl | 1 M | 100 | 10 | 20 min | NaOH | 1 M | 80 | 10 | 24 h | a 2.59%–4.23% b 73%–96.75% | [88] | |
| Bombyx eri | larvae | HCl | 1 M | 80 | N/A | 35 | NaOH | 1 M | 80 | NA | 24 h | a 45% | [85] | |
| Crustaceans, Mollusks, and Other Aquatic Invertebrates | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sources | Demineralization | Deproteinization | Yield | References | |||||||||
| Scientific Name | Common Name | Type of Acid | Concentration | Temperature (°C) | Solution-to-Solid Ratio (mL/g) | Duration | Type of Alkali | Concentration | Temperature (°C) | Solution-to-Solid Ratio (mL/g) | Duration | ||
| Solenocera melantho | Pink shrimp | HCl | 1.7 N | RT | 9 | N/A | NaOH | 2.5 N | 75 | 5 | N/A | a 22% | [56] |
| Palaemon fabricius | Shrimp | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 22% | [13] |
| Squilla mantis | Squilla shrimp | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 24% | [13] |
| Parapenaeopsis stylifera | Shrimp shells | HCl | 0.25 M | RT | N/A | N/A | NaOH | 1 M | RT | N/A | N/A | a 20% | [101] |
| Pandalus borealus | Shrimp shells | HCl | 1 M | 100 | N/A | 20 min | NaOH | 1 M | 85 | N/A | 24 h | a 6.89% | [55] |
| Repeated several times during 24 h | |||||||||||||
| Penaeus aztecus | Brown shrimp shells | HCl | 1 M | RT | N/A | N/A | NaOH | 1 M | 105–110 | N/A | N/A | a 21.53% | [99] |
| Penaeus monodon | Black tiger shrimp waste | HCOOH C6H8O7 | 0.25 M 0.25 M | N/A | 30 28 | 30 min | NaOH | 1 M | 95 | 15 | 6 h | N/A | [104] |
| Penaeus durarum | Pink shrimp shells | HCl | 1 N | 100 | N/A | 20 min | NaOH | 1 N | 80 | N/A | 36 h | a 23.72% | [86] |
| Refluxed with NaCO3, 12 h | |||||||||||||
| Parapenaeus longirostris | Shrimp | HCl | 1.23 N | RT | N/A | 1 h | NaOH | 3% | 100 | N/A | 15 min | N/A | [105] |
| Squilla mantis | Squilla shrimp | HCl | 1.23 N | RT | N/A | 1 h | NaOH | 3% | 100 | N/A | 15 min | N/A | [105] |
| Farfantepenaeus brasiliensis | Pink shrimp | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | N/A | [106] |
| Penaeus carinatus | Shrimps | HCl | 1% | RT | N/A | 24 h | NaOH | 2%–4% | +100 | N/A | 1 h | b 35.49% | [24] |
| Penaeus monodon | Shrimps | HCl | 1% | RT | N/A | 24 h | NaOH | 2%–4% | +100 | N/A | 1 h | b 35.49% | [24] |
| Lipopenaeus vannamei | White shrimps | HCl | 4% | RT | N/A | 4 h | NaOH | 5% | 90 | (w/v 1:10) | 12 h | N/A | [107] |
| N/A | Shrimp shells | HCl | N/A | RT | N/A | 15 min | NaOH | N/A | N/A | N/A | 20 min | a 14.8% | [91] |
| NaClO treatments for 10 min each before demineralization and deproteinization | |||||||||||||
| Penaeus japonicus | Prawns | HCl | 2 N | RT | 50 | 48 h | NaOH | 1 N | 100 | 100 | 96 h | NA | [108] |
| Macrobrachium rosenbergii | Prawns shells | HCl | 0.25 M | 40 | N/A | 4 h | NaOH | 0.25 M | N/A | N/A | 4 h | a 8.28%–5.02% | [109] |
| Litopenaeus vannameil | Prawn shells | HCl | 0.5%–1% | 25 | 4–10 | 24 h | NaOH | 5% | 60 | 8 | 2 h | a 35.00% b 25.00% | [110] |
| Penaeus monodon | Giant tiger prawn | HCl | 1 M | RT | 15 | N/A | NaOH | 1 M | 100 | N/A | 8 h | a 10% | [92] |
| Paralithodes camstschatecus | Crabs | HCl | 2 N | RT | 50 | 48 h | NaOH | 1 N | 100 | 100 | 96 h | NA | [108] |
| Grapsus marmoratus | Marbled crab | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 10% | [13] |
| Portunus puber | Red crab | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 10% | [13] |
| Maia squinado | Spider crab | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 16% | [13] |
| Paralithodes camtschaticus | Crab | HCl | 7% | RT | N/A | 48 h | KOH | 5% | N/A | N/A | 6 h | a 12.1% | [111] |
| N/A | Rice-field crab shells | HCl | 1 N | 100 | N/A | 20 min | NaOH | 1 N | 80 | N/A | 36 h | a 15.2% | [86] |
| Refluxed with NaCO3, 12 h | |||||||||||||
| N/A | Crab shells | HCl | 1 N | 100 | N/A | 20 min | NaOH | 1 N | 80 | N/A | 36 h | a 16.73% | [86] |
| Refluxed with NaCO3, 12 h | |||||||||||||
| N/A | Crab | HCl | N/A | RT | N/A | 15 min | NaOH | N/A | N/A | N/A | 20 min | a 13.4% | [91] |
| NaClO treatments for 10 min each before demineralization and deproteinization | |||||||||||||
| Portunus pelagicus | Blue swimming crab | HCl | 2 N | RT | 50 | 48 h | NaOH | 1 N | 100 | 100 | 96 h | NA | [84] |
| Potamon algeriense | Crab | HCl | 1 N | RT | 15 | 6 h | NaOH | 3% | 121 | 10 | 20 min | a 8.27% b 5.89% | [112] |
| Autoclave at 15 psi | |||||||||||||
| Macropipus holsatus | Crab | HCl | 4% | RT | 15 | 1 h | NaOH | 3% | 65 | 20 | 2 h | a 12.23% b 9.52% | [113] |
| Panulirus japonicus | Lobsters | HCl | 2 N | RT | 50 | 48 h | NaOH | 1 N | 100 | 100 | 96 h | NA | [108] |
| Homarus vulgaris | Lobster | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 17% | [13] |
| Scyllarus arctus | Locust lobster | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 25% | [13] |
| Palinurus vulgaris | Spiny lobster | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 32% | [13] |
| Astacus fluviatilis | Crayfish | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 36% | [13] |
| Procambarus clarkii | Crayfish | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 20.6% | [13] |
| N/A | Crayfish | HCl | N/A | RT | N/A | 15 min | NaOH | N/A | N/A | N/A | 20 min | a 15.3% | [91] |
| NaClO treatments for 10 min each before demineralization and deproteinization | |||||||||||||
| Sepia officinalis | Cuttlefish | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 20% | [13] |
| Loligo vulgaris | Squid | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 40% | [13] |
| N/A | Cuttlefish pens | HCl | 1 M | RT | N/A | N/A | NaOH | 1 M | 105–110 | N/A | N/A | a 5.4% | [99] |
| N/A | Squid pens | HCl | 1 M | RT | N/A | N/A | NaOH | 1 M | 105–110 | N/A | N/A | a 49% | [99] |
| N/A | Mussel shells | HCl | 0.68 M | RT | 10 | 6 h | NaOH | 0.62 M | N/A | 10 | 16 h | a 23.25% | [114] |
| Oniscus asellus | Woodlouse | HCl | 4 M | 75 | N/A | 2 h | NaOH | 4 M | 150 | N/A | 18 h | a 6%–7% | [90] |
| Reflux in magnetic stirrer | Heat in magnetic stirrer | ||||||||||||
| Lepas anatifera | Barnacles | HCl | 0.55 M | RT | N/A | 1–24 h | NaOH | 0.3 M | 80–85 | N/A | 1 h | a 7% | [13] |
| Chelonibia patula | Barnacles | HCl | 1 M | N/A | N/A | 10 min | NaOH | 2 M | N/A | N/A | 20 min | a 3.1% | [102] |
| Daphnia magna | Water flea | HCl | 1 M | 65–75 | N/A | 6 h | NaOH | 1 M | 65 | N/A | 20h | a 13%–21% | [115] |
| Conus inscriptus | Sea snail | HCl | 1 M | 60 | N/A | 30 min | NaOH | 3 M | 80 | N/A | 2 h | a 21.65% | [103] |
| Others | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sources | Demineralization | Deproteinization | Yield | References | |||||||||
| Scientific Name | Common Name | Type of Acid | Concentration | Temperature (°C) | Solution-to-Solid Ratio (mL/g) | Duration | Type of Alkali | Concentration | Temperature (°C) | Solution-to-Solid Ratio (mL/g) | Duration | ||
| Pleurotus eryngii | Mushrooms | HCl | 2 M | RT | N/A | 48 h | NaOH | 2% | 100 | N/A | 48 h | N/A | [117] |
| Agaricus bisporus | Mushrooms | HCl | 2 M | RT | N/A | 48 h | NaOH | 2% | 100 | N/A | 48 h | N/A | [117] |
| Lentinula edodes | Mushrooms | HCl | 2 M | RT | N/A | 48 h | NaOH | 2% | 100 | N/A | 48 h | N/A | [117] |
| Grifola frondosa | Mushrooms | HCl | 2 M | RT | N/A | 48 h | NaOH | 2% | 100 | N/A | 48 h | N/A | [117] |
| Hypsizygus marmoreus | Mushrooms | HCl | 2 M | RT | N/A | 48 h | NaOH | 2% | 100 | N/A | 48 h | N/A | [117] |
| N/A | Chicken feet | HCl | 0.5–1.5 N | N/A | 10–20 | N/A | NaOH | 0.1–2 N | N/A | 5–20 | N/A | a 15.2% | [70] |
| Geolycosa vultuosa | Spider | HCl | 4 M | 60–65 | N/A | 1 h | NaOH | 1 M | 130–135 | N/A | 16 h | a 8%–8.5% | [116] |
| Hogna radiata | Spider | HCl | 4 M | 60–65 | N/A | 1 h | NaOH | 1 M | 130–135 | N/A | 16 h | a 6.5%–7% | [116] |
| Labeo rohit | Fish scales | HCl | 1% | RT | N/A | 36 h | NaOH | 2 N | RT | N/A | 36 h | N/A | [118] |
| Labeo rohit | Fish scales | HCl | 1% | RT | N/A | N/A | NaOH | 0.5 N | RT | N/A | 18 h | N/A | [119] |
| Plumatella repens | Creeping bryozoan or moss animal | HCl | 2 M | 100 | N/A | 2 h | NaOH | 2 M | 140 | N/A | 20h | a 13.3% | [96] |
| Fomes fomentarius | Fungi | HCl | 2 M | 100 | N/A | 2 h | NaOH | 2 M | 140 | N/A | 20h | a 2.4% | [96] |
| Zophobas morio | Superworm larvae | HCl | 1 M | 35 | 20 | 30 min | NaOH | 0.5–2 M | 80 | 20 | 20h | a 4.77%–5.43% b 65.84%–75.52% | [120] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zainol Abidin, N.A.; Kormin, F.; Zainol Abidin, N.A.; Mohamed Anuar, N.A.F.; Abu Bakar, M.F. The Potential of Insects as Alternative Sources of Chitin: An Overview on the Chemical Method of Extraction from Various Sources. Int. J. Mol. Sci. 2020, 21, 4978. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144978
Zainol Abidin NA, Kormin F, Zainol Abidin NA, Mohamed Anuar NAF, Abu Bakar MF. The Potential of Insects as Alternative Sources of Chitin: An Overview on the Chemical Method of Extraction from Various Sources. International Journal of Molecular Sciences. 2020; 21(14):4978. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144978
Chicago/Turabian StyleZainol Abidin, Nurul Alyani, Faridah Kormin, Nurul Akhma Zainol Abidin, Nor Aini Fatihah Mohamed Anuar, and Mohd Fadzelly Abu Bakar. 2020. "The Potential of Insects as Alternative Sources of Chitin: An Overview on the Chemical Method of Extraction from Various Sources" International Journal of Molecular Sciences 21, no. 14: 4978. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21144978