The Role of miR-375-3p and miR-200b-3p in Gastrointestinal Stromal Tumors

 , , , , and
, , , , and 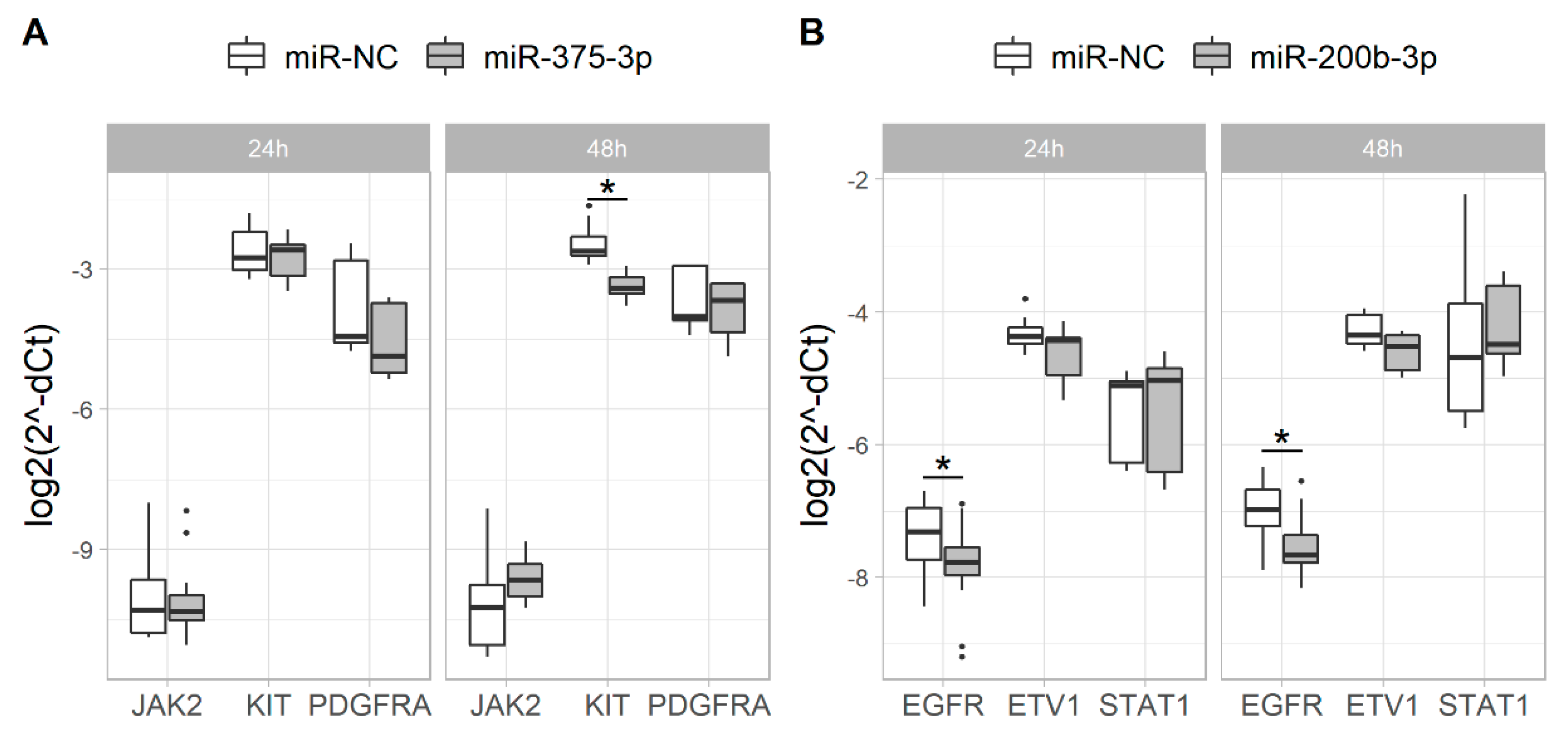{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Overexpression of miR-375-3p and miR-200b-3p Alters the Expression of Their Putative Target Genes
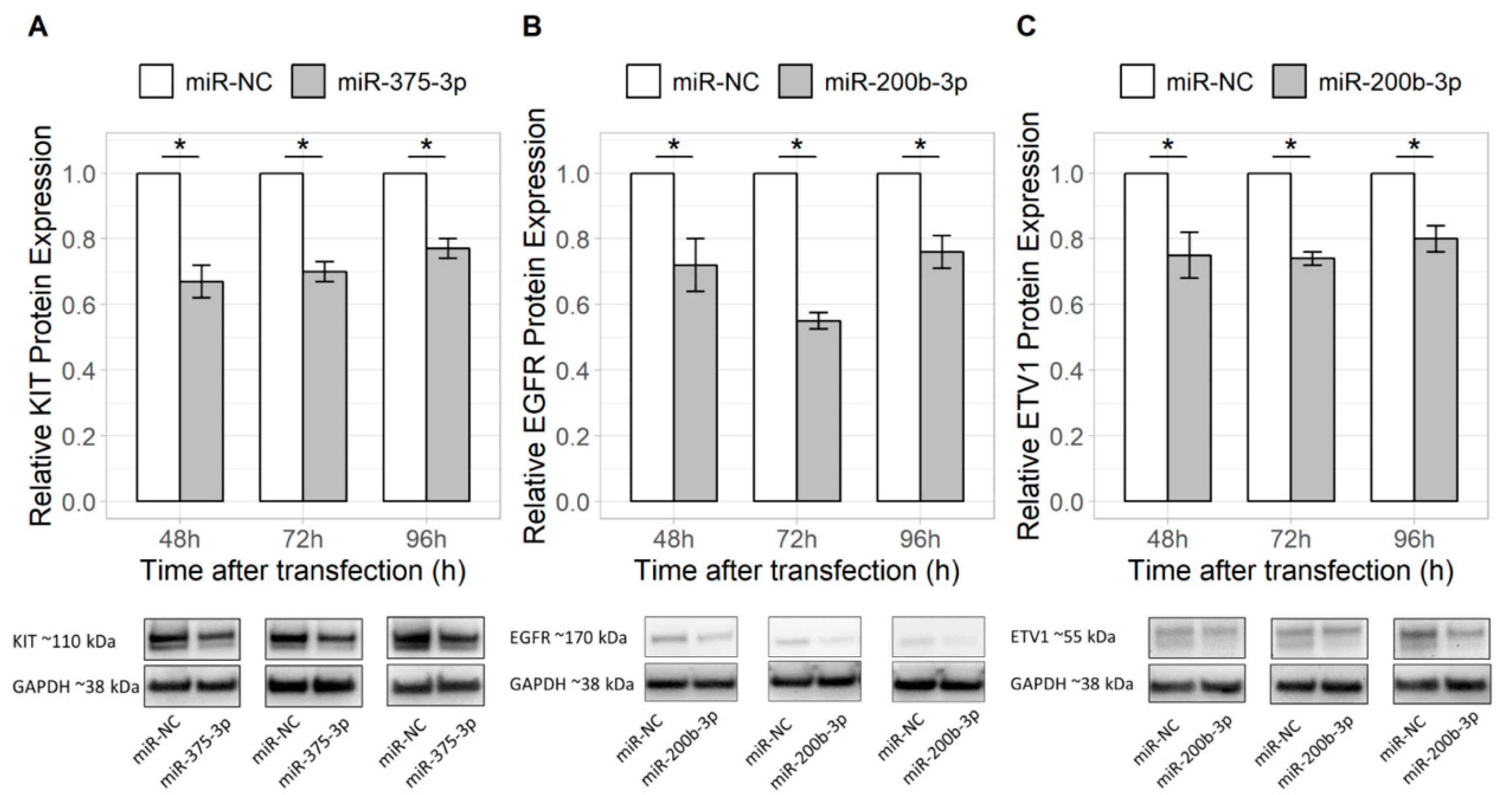
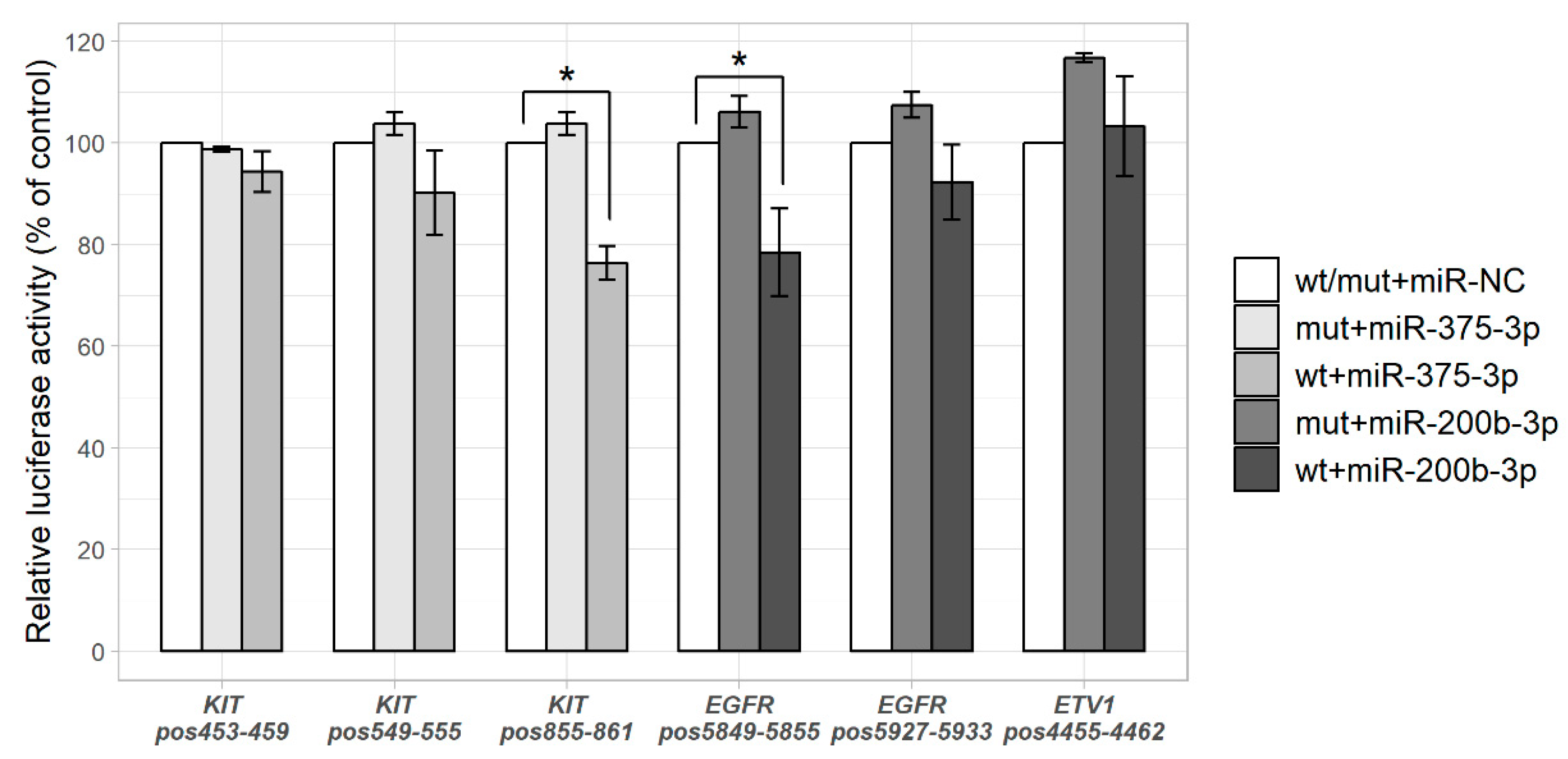
2.2. miR-375-3p and miR-200b-3p Directly Regulate Their Predicted Targets KIT and EGFR
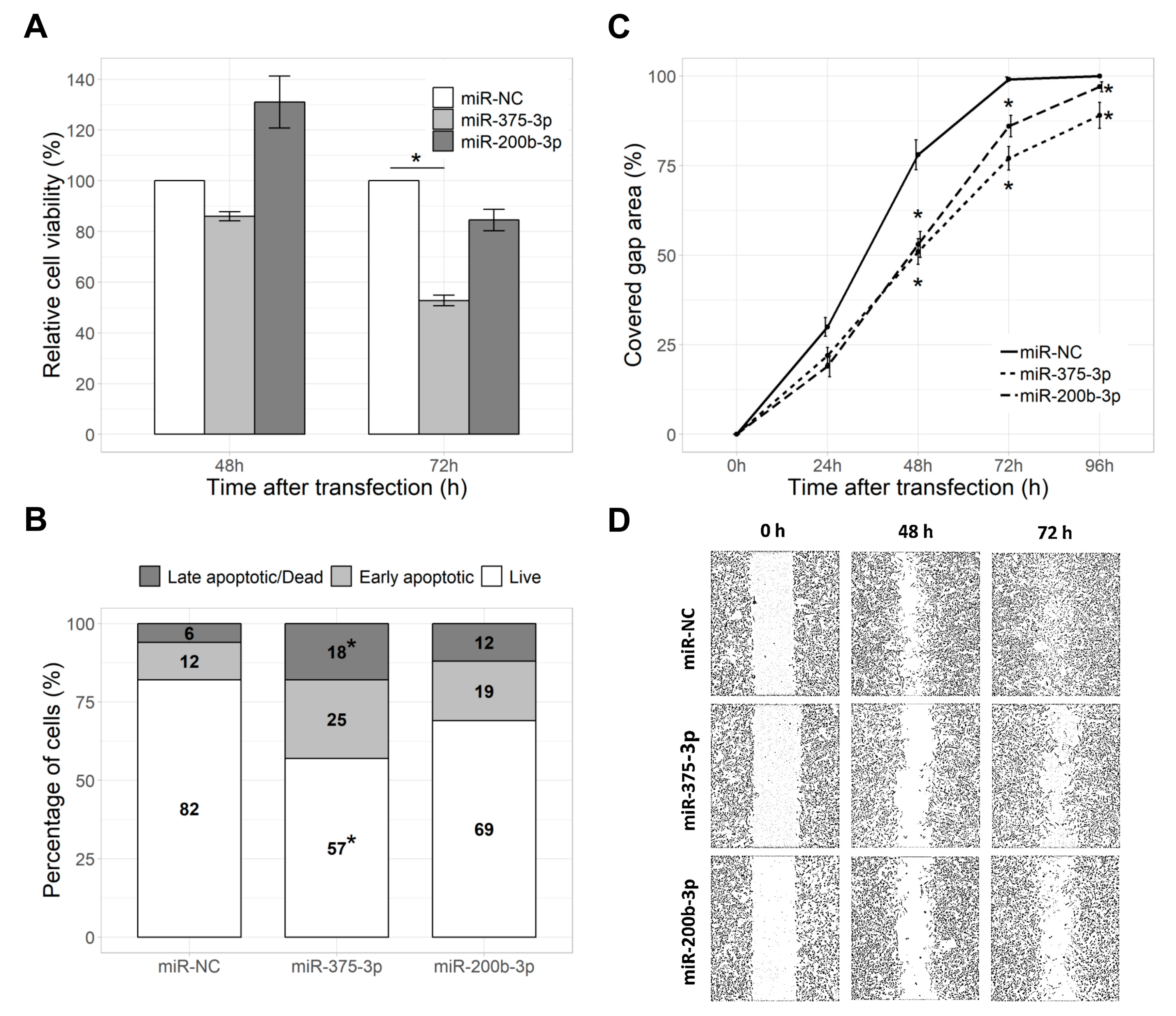
2.3. miR-375-3p Reduced Cell Viability and Proliferation
2.4. miR-375-3p and miR-200b-3p Reduced Cell Migration Rate
2.5. miR-375-3p and miR-200b-3p Did Not Affect Cell Apoptosis
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. miRNA Selection and Target Prediction
4.3. Cell Transfection
4.4. Quantitative Reverse Transcription PCR
4.5. Western Blot
4.6. Luciferase Reporter Assay
4.7. MTT Assay
4.8. Apoptosis Assay
4.9. Wound Healing Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 3′UTR | three prime untranslated region |
| ATCC | American type culture collection |
| DMSO | dimethyl sulfoxide |
| EGFR | epidermal growth factor receptor |
| ETV1 | ETS transcription factor 1 |
| FITC | fluorescein isothiocyanate |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| GIST | gastrointestinal stromal tumor |
| JAK | Janus kinase |
| MAPK | mitogen-activated protein kin |
| miRNA | microRNA |
| mTOR | mammalian target of rapamycin |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| mut | mutant |
| NC | negative control |
| PDGFRA | platelet-derived growth factor receptor-α |
| PI | propidium iodide |
| PI3K | phosphatidylinositol-3-kinase |
| RIPA | radioimmunoprecipitation assay |
| RNA | ribonucleic acid |
| RT-qPCR | Reverse transcription quantitative polymerase chain reaction |
| RTK | receptor tyrosine kinase |
| STAT | signal transducer and activator of transcription |
| wt | wild-type |
References
- Kitamura, Y. Gastrointestinal stromal tumors: Past, present, and future. J. Gastroenterol. 2008, 43, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Corless, C.L.; Barnett, C.M.; Heinrich, M.C. Gastrointestinal stromal tumours: Origin and molecular oncology. Nat. Rev. Cancer 2011, 11, 865–878. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, I.M.; Mariño-Enríquez, A.; Fletcher, J.A. What is New in Gastrointestinal Stromal Tumor? Adv. Anat. Pathol. 2017, 24, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Liegl-Atzwanger, B.; Fletcher, J.A.; Fletcher, C.D.M. Gastrointestinal stromal tumors. Virchows Arch. 2010, 456, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Chi, P.; Chen, Y.; Zhang, L.; Guo, X.; Wongvipat, J.; Shamu, T.; Fletcher, J.A.; Dewell, S.; Maki, R.G.; Zheng, D.; et al. ETV1 is a lineage survival factor that cooperates with KIT in gastrointestinal stromal tumours. Nature 2010, 467, 849–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Cheng, H.; Li, Z.; Pang, Y.; Jia, X.; Xie, F.; Hu, G.; Cai, Q.; Wang, Y. Genetic progression in gastrointestinal stromal tumors: Mechanisms and molecular interventions. Oncotarget 2017, 8, 60589–60604. [Google Scholar] [CrossRef] [Green Version]
- Lujambio, A.; Lowe, S.W. The microcosmos of cancer. Nature 2012, 482, 347–355. [Google Scholar] [CrossRef]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Streleckiene, G.; Inciuraite, R.; Juzenas, S.; Salteniene, V.; Steponaitiene, R.; Gyvyte, U.; Kiudelis, G.; Leja, M.; Ruzgys, P.; Satkauskas, S.; et al. Mir-20b and mir-451a are involved in gastric carcinogenesis through the PI3K/AKT/mTOR signaling pathway: Data from gastric cancer patients, cell lines and ins-gas mouse model. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [Green Version]
- Juzėnas, S.; Saltenienė, V.; Kupcinskas, J.; Link, A.; Kiudelis, G.; Jonaitis, L.; Jarmalaite, S.; Kupcinskas, L.; Malfertheiner, P.; Skieceviciene, J. Analysis of Deregulated microRNAs and Their Target Genes in Gastric Cancer. PLoS ONE 2015, 10, e0132327. [Google Scholar] [CrossRef]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as potential cancer biomarkers: The advantage and disadvantage. Clin. Epigenetics 2018, 10, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, M.Y.; Ferrajoli, A.; Sood, A.K.; Lopez-Berestein, G.; Calin, G.A. microRNA Therapeutics in Cancer—An Emerging Concept. EBioMedicine 2016, 12, 34–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Link, A.; Kupcinskas, J. MicroRNAs as non-invasive diagnostic biomarkers for gastric cancer: Current insights and future perspectives. World J. Gastroenterol. 2018, 24, 3313–3329. [Google Scholar] [CrossRef] [PubMed]
- Kupcinskas, J. Small molecules in rare tumors: Emerging role of microRNAs in GIST. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [Green Version]
- Gyvyte, U.; Juzenas, S.; Salteniene, V.; Kupcinskas, J.; Poskiene, L.; Kucinskas, L.; Jarmalaite, S.; Stuopelyte, K.; Steponaitiene, R.; Hemmrich-Stanisak, G.; et al. MiRNA profiling of gastrointestinal stromal tumors by next generation sequencing. Oncotarget 2017, 8, 37225–37238. [Google Scholar] [CrossRef]
- Yan, J.-W.; Lin, J.-S.; He, X.-X. The emerging role of miR-375 in cancer. Int. J. Cancer 2014, 135, 1011–1018. [Google Scholar] [CrossRef]
- Li, L.; Jia, L.; Ding, Y. Upregulation of miR-375 inhibits human liver cancer cell growth by modulating cell proliferation and apoptosis via targeting ErbB2. Oncol. Lett. 2018, 16, 3319–3326. [Google Scholar] [CrossRef]
- Yang, D.; Yan, R.; Zhang, X.; Zhu, Z.; Wang, C.; Liang, C.; Zhang, X. Deregulation of MicroRNA-375 inhibits cancer proliferation migration and chemosensitivity in pancreatic cancer through the association of HOXB3. Am. J. Transl. Res. 2016, 8, 1551. [Google Scholar]
- Mao, Q.; Quan, T.; Luo, B.; Guo, X.; Liu, L.; Zheng, Q. MiR-375 targets KLF4 and impacts the proliferation of colorectal carcinoma. Tumor Biol. 2016, 37, 463–471. [Google Scholar] [CrossRef]
- Guo, F.; Gao, Y.; Sui, G.; Jiao, D.; Sun, L.; Fu, Q.; Jin, C. miR-375-3p/YWHAZ/β-catenin axis regulates migration, invasion, EMT in gastric cancer cells. Clin. Exp. Pharmacol. Physiol. 2019, 46, 144–152. [Google Scholar] [CrossRef]
- Yuan, K.-T.; Li, B.-X.; Yuan, Y.-J.; Tan, M.; Tan, J.-F.; Dai, W.-G.; Feng, W.-D.; Zuo, J.-D. Deregulation of MicroRNA-375 Inhibits Proliferation and Migration in Gastric Cancer in Association With Autophagy-Mediated AKT/mTOR Signaling Pathways. Technol. Cancer Res. Treat. 2018, 17, 153303381880649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duensing, A.; Medeiros, F.; McConarty, B.; Joseph, N.E.; Panigrahy, D.; Singer, S.; Fletcher, C.D.M.; Demetri, G.D.; Fletcher, J.A. Mechanisms of oncogenic KIT signal transduction in primary gastrointestinal stromal tumors (GISTs). Oncogene 2004, 23, 3999–4006. [Google Scholar] [CrossRef] [Green Version]
- Bauer, S.; Duensing, A.; Demetri, G.D.; Fletcher, J.A. KIT oncogenic signaling mechanisms in imatinib-resistant gastrointestinal stromal tumor: PI3-kinase/AKT is a crucial survival pathway. Oncogene 2007, 26, 7560–7568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, F.; Ehlers, I.; Agosti, V.; Socci, N.D.; Viale, A.; Sommer, G.; Yozgat, Y.; Manova, K.; Antonescu, C.R.; Besmer, P. Oncogenic Kit signaling and therapeutic intervention in a mouse model of gastrointestinal stromal tumor. Proc. Natl. Acad. Sci. USA 2006, 103, 12843–12848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lasota, J.; Kowalik, A.; Felisiak-Golabek, A.; Ziȩba, S.; Wang, Z.F.; Miettinen, M. New Mechanisms of mTOR Pathway Activation in KIT-mutant Malignant GISTs. Appl. Immunohistochem. Mol. Morphol. 2017, 27, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Xu, Y.; Zhang, W.; Deng, Y.; Si, M.; Du, Y.; Yao, H.; Liu, X.; Ke, Y.; Si, J.; et al. MiR-375 frequently downregulated in gastric cancer inhibits cell proliferation by targeting JAK2. Cell Res. 2010, 20, 784–793. [Google Scholar] [CrossRef]
- Miao, L.; Liu, K.; Xie, M.; Xing, Y.; Xi, T. miR-375 inhibits Helicobacter pylori-induced gastric carcinogenesis by blocking JAK2–STAT3 signaling. Cancer Immunol. Immunother. 2014, 63, 699–711. [Google Scholar] [CrossRef]
- Paner, G.P.; Silberman, S.; Hartman, G.; Micetich, K.C.; Aranha, G.V.; Alkan, S. Analysis of signal transducer and activator of transcription 3 (STAT3) in gastrointestinal stromal tumors. Anticancer Res. 2003, 23, 2253. [Google Scholar]
- Huang, G.L.; Sun, J.; Lu, Y.; Liu, Y.; Cao, H.; Zhang, H.; Calin, G.A. MiR-200 family and cancer: From a meta-analysis view. Mol. Asp. Med. 2019, 70, 57–71. [Google Scholar] [CrossRef]
- Li, D.; Wang, H.; Song, H.; Xu, H.; Zhao, B.; Wu, C.; Hu, J.; Wu, T.; Xie, D.; Zhao, J.; et al. The microRNAs miR-200b-3p and miR-429-5p target the LIMK1/CFL1 pathway to inhibit growth and motility of breast cancer cells. Oncotarget 2017, 8, 85276. [Google Scholar] [CrossRef] [Green Version]
- Humphries, B.; Wang, Z.; Li, Y.; Jhan, J.R.; Jiang, Y.; Yang, C. ARHGAP18 downregulation by miR-200b suppresses metastasis of triple-negative breast cancer by enhancing activation of RhoA. Cancer Res. 2017, 77, 4051–4064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Guan, B.; Liu, J.; Zhang, Z.; He, S.; Zhan, Y.; Su, B.; Han, H.; Zhang, X.; Wang, B.; et al. MicroRNA-200b is downregulated and suppresses metastasis by targeting LAMA4 in renal cell carcinoma. EBioMedicine 2019, 44, 439–451. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, X.; Zhu, Y.; Zhu, J.; Lai, Q. miR-200b-3p inhibits proliferation and induces apoptosis in colorectal cancer by targeting Wnt1. Mol. Med. Rep. 2018, 18, 2571. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Li, Y.; Xu, W.; He, L.; Tan, Y.; Xu, H. Long non-coding RNA ZFAS1 regulates the malignant progression of gastric cancer via the microRNA-200b-3p/Wnt1 axis. Biosci. Biotechnol. Biochem. 2019, 83, 1289–1299. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Zhang, W.; Tan, J.; Ma, J.; Zhao, J. MiR-200b-3p Functions as an Oncogene by Targeting ABCA1 in Lung Adenocarcinoma. Technol. Cancer Res. Treat. 2019, 18, 153303381989259. [Google Scholar] [CrossRef]
- Zhang, Z.; Xing, T.; Chen, Y.; Xiao, J. Exosome-mediated miR-200b promotes colorectal cancer proliferation upon TGF-β1 exposure. Biomed. Pharmacother. 2018, 106, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Sigismund, S.; Avanzato, D.; Lanzetti, L. Emerging functions of the EGFR in cancer. Mol. Oncol. 2018, 12, 3–20. [Google Scholar] [CrossRef]
- Jiang, J.; Jin, M.-S.; Suo, J.; Wang, Y.-P.; He, L.; Cao, X.-Y. Evaluation of malignancy using Ki-67, p53, EGFR and COX-2 expressions in gastrointestinal stromal tumors. World J. Gastroenterol. 2012, 18, 2569. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Montgomery, E. Expression of EGFR in Gastric Stromal Tumors. Appl. Immunohistochem. Mol. Morphol. 2008, 16, 310–315. [Google Scholar] [CrossRef]
- Mahadevan, D.; Theiss, N.; Morales, C.; Stejskal, A.E.; Cooke, L.S.; Zhu, M.; Kurtzman, D.; Swart, R.; Ong, E.; Qi, W. Novel receptor tyrosine kinase targeted combination therapies for imatinib-resistant gastrointestinal stromal tumors (GIST). Oncotarget 2015, 6, 1954. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, T.; Sonobe, H.; Toyonaga, S.; Yamasaki, I. Conventional and Molecular Cytogenetic Characterization of a New Human Cell Line, GIST-T1, Established from Gastrointestinal Stromal Tumor. Lab. Investig. 2002, 82, 663–665. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, M.C.; Griffith, D.J.; Druker, B.J.; Wait, C.L.; Ott, K.A.; Zigler, A.J. Inhibition of c-kit receptor tyrosine kinase activity by STI 571, a selective tyrosine kinase inhibitor. Blood 2000, 96, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Young, L.; Sung, J.; Masters, J.R. Detection of mycoplasma in cell cultures. Nat. Protoc. 2010, 5, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4. [Google Scholar] [CrossRef] [PubMed]
- Ihle, M.A.; Trautmann, M.; Kuenstlinger, H.; Huss, S.; Heydt, C.; Fassunke, J.; Wardelmann, E.; Bauer, S.; Schildhaus, H.-U.; Buettner, R.; et al. miRNA-221 and miRNA-222 induce apoptosis via the KIT/AKT signalling pathway in gastrointestinal stromal tumours. Mol. Oncol. 2015, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gyvyte, U.; Lukosevicius, R.; Inciuraite, R.; Streleckiene, G.; Gudoityte, G.; Bekampyte, J.; Valentini, S.; Salteniene, V.; Ruzgys, P.; Satkauskas, S.; et al. The Role of miR-375-3p and miR-200b-3p in Gastrointestinal Stromal Tumors. Int. J. Mol. Sci. 2020, 21, 5151. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145151
Gyvyte U, Lukosevicius R, Inciuraite R, Streleckiene G, Gudoityte G, Bekampyte J, Valentini S, Salteniene V, Ruzgys P, Satkauskas S, et al. The Role of miR-375-3p and miR-200b-3p in Gastrointestinal Stromal Tumors. International Journal of Molecular Sciences. 2020; 21(14):5151. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145151
Chicago/Turabian StyleGyvyte, Ugne, Rokas Lukosevicius, Ruta Inciuraite, Greta Streleckiene, Greta Gudoityte, Justina Bekampyte, Serena Valentini, Violeta Salteniene, Paulius Ruzgys, Saulius Satkauskas, and et al. 2020. "The Role of miR-375-3p and miR-200b-3p in Gastrointestinal Stromal Tumors" International Journal of Molecular Sciences 21, no. 14: 5151. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21145151