Alterations of STEP46 and STEP61 Expression in the Rat Retina with Age and AMD-Like Retinopathy Development

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
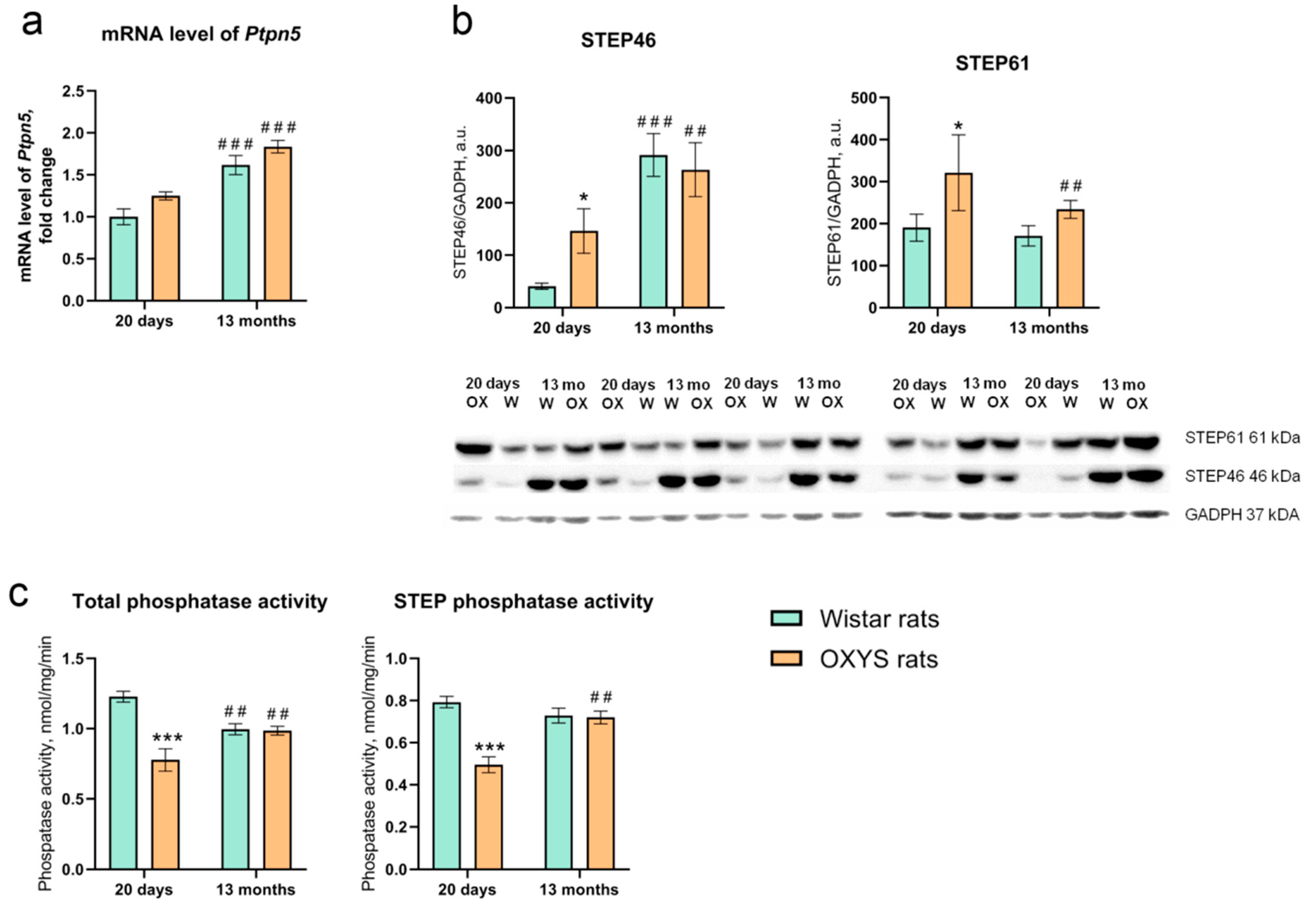
2.1. Age-Dependent Alterations of Ptpn5 mRNA Expression in the Retina of OXYS and Wistar Rats
2.2. Alterations of STEP46 and STEP61 Protein Levels with Age and the Development of AMD-Like Retinopathy
2.3. Total Phosphatase and STEP Phosphatase Activities in the Retina of OXYS and Wistar Rats
2.4. Effects of TC-2153 on the Ptpn5 mRNA level, STEP46 and STEP61 Protein Levels, and Total Phosphatase and STEP Activities in the OXYS Rat Retina
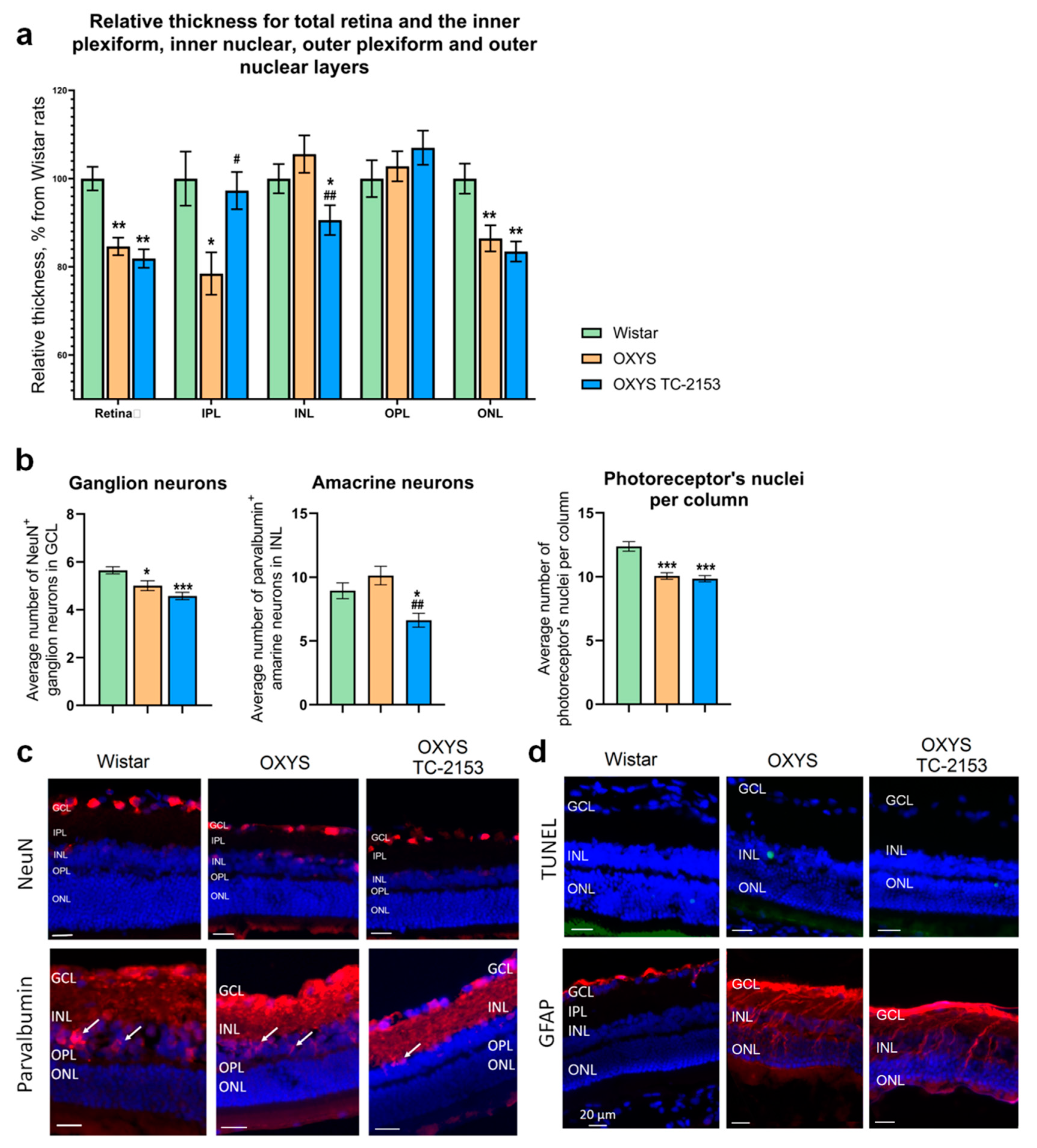
2.5. Effects of the STEP Inhibitor TC-2153 on Retinal Apoptosis and Thickness in OXYS Rats
2.6. Effects of the STEP Inhibitor TC-2153 on the Amounts of Amacrine Neurons, Ganglion Neurons, and Photoreceptors
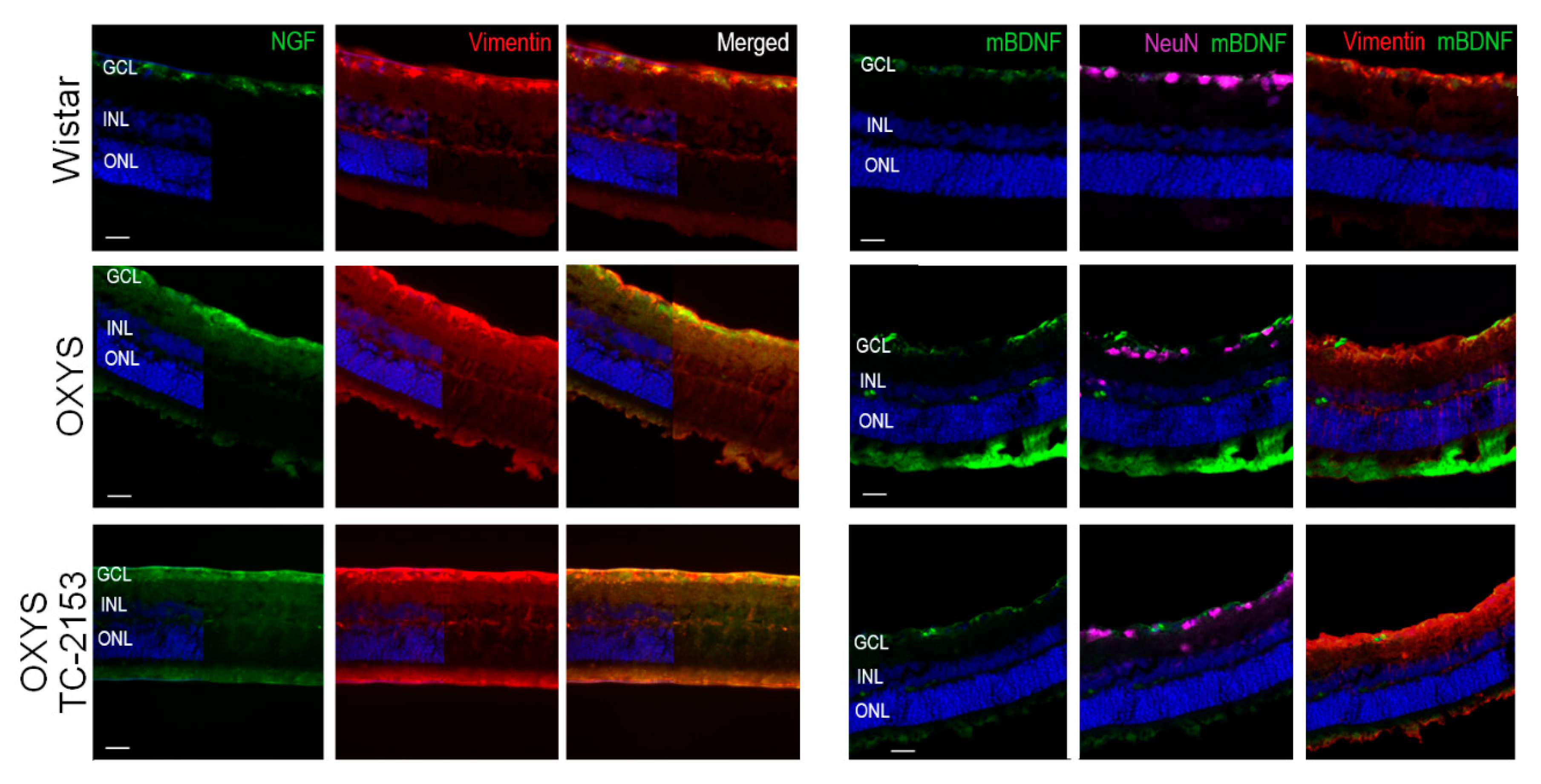
2.7. Effects of the STEP Inhibitor TC-2153 on Macroglia and NGF and BDNF Expression
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals and Experimental Procedures
4.3. Western Blot Analysis
4.4. Quantitative Reverse-Transcription PCR (qPCR)
4.5. A Real-Time Spectrophotometric Assay of Phosphatase and STEP Activities
4.6. Immunohistochemistry and the TUNEL Assay
4.7. Measurement of Retinal Thickness and Cell Counting
4.8. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease |
| AMD | Age-related macular degeneration |
| ANOVA | Analysis of variance |
| GCL | Ganglion cell layer |
| INL | Inner nuclear layer |
| IPL | Inner plexiform layer |
| ONL | Outer nuclear layer |
| OPL | Outer plexiform layer |
| P | Postnatal day |
| RPE | Retinal pigment epithelium |
| SDS | Sodium dodecyl sulfate |
| STEP | Striatal-enriched tyrosine protein phosphatase |
| TC-2153 | 8-(trifluoromethyl)-1,2,3,4,5-benzopentathiepin-6-amine |
References
- Kurup, P.; Zhang, Y.; Venkitaramani, D.V.; Xu, J.; Lombroso, P.J. The role of STEP in Alzheimer’s disease. Channels 2010, 4, 347–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Kurup, P.; Nairn, A.; Lombroso, P.J. Striatal-enriched protein tyrosine phosphatase in Alzheimer’s disease. Adv. Pharm. 2012, 64, 303–325. [Google Scholar]
- Kamceva, M.; Benedict, J.; Nairn, A.C.; Lombroso, P.J. Role of Striatal-Enriched Tyrosine Phosphatase in Neuronal Function. Neural Plast. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorber, B.; Berry, M.; Hendriks, W.J.A.J.; Hertog, J.D.; Pulido, R.; Logan, A. Stimulated regeneration of the crushed adult rat optic nerve correlates with attenuated expression of the protein tyrosine phosphatases RPTPα, STEP, and LAR. Mol. Cell. Neurosci. 2004, 27, 404–416. [Google Scholar] [CrossRef]
- Kaarniranta, K.; Salminen, A.; Haapasalo, A.; Soininen, H.; Hiltunen, M. Age-Related Macular Degeneration (AMD): Alzheimer’s Disease in the Eye? J. Alzheimer’s Dis. 2011, 24, 615–631. [Google Scholar] [CrossRef] [Green Version]
- Fritsche, L.; Fariss, R.N.; Stambolian, D.; Abecasis, G.R.; Curcio, C.A.; Swaroop, A. Age-related macular degeneration: Genetics and biology coming together. Annu. Rev. Genom. Hum. Genet. 2014, 15, 151–171. [Google Scholar] [CrossRef] [Green Version]
- DeAngelis, M.M.; Owen, L.A.; Morrison, M.A.; Morgan, D.J.; Li, M.; Shakoor, A.; Vitale, A.; Iyengar, S.; Stambolian, D.; Kim, I.; et al. Genetics of age-related macular degeneration (AMD). Hum. Mol. Genet. 2017, 26. [Google Scholar] [CrossRef] [Green Version]
- Kolosova, N.G.; Stefanova, N.A.; Korbolina, E.E.; Fursova, A.Z.; Kozhevnikova, O.S. Senescence-accelerated OXYS rats: A genetic model of premature aging and age-related diseases. Adv. Gerontol. 2014, 4, 294–298. [Google Scholar] [CrossRef]
- Kozhevnikova, O.S.; Korbolina, E.E.; Stefanova, N.A.; Muraleva, N.A.; Orlov, Y.L.; Kolosova, N.G. Association of AMD-like retinopathy development with an Alzheimer’s disease metabolic pathway in OXYS rats. Biogerontology 2013, 14, 753–762. [Google Scholar] [CrossRef]
- Kolosova, N.G.; Kozhevnikova, O.S.; Telegina, D.V.; Fursova, A.Z.; Stefanova, N.A.; Muraleva, N.; Venanzi, F.; Sherman, M.Y.; Kolesnikov, S.I.; Sufianov, A.; et al. p62 /SQSTM1 coding plasmid prevents age related macular degeneration in a rat model. Aging 2018, 10, 2136–2147. [Google Scholar] [CrossRef]
- Telegina, D.V.; Kozhevnikova, O.S.; Bayborodin, S.I.; Kolosova, N.G. Contributions of age-related alterations of the retinal pigment epithelium and of glia to the AMD-like pathology in OXYS rats. Sci. Rep. 2017, 7, 41533. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, E.; Kulikov, A.; Kulikov, E.K.A.A. Striatal-enriched Tyrosine Protein Phosphatase (STEP) in the Mechanisms of Depressive Disorders. Curr. Protein Pept. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, E.; Khotskin, N.; Illarionova, N.; Sorokin, I.; Bazhenova, E.; Kondaurova, E.; Volcho, K.P.; Khomenko, T.; Salakhutdinov, N.; Ponimaskin, E.; et al. Inhibitor of Striatal-Enriched Protein Tyrosine Phosphatase, 8-(Trifluoromethyl)-1,2,3,4,5-Benzopentathiepin-6-Amine hydrochloride (TC-2153), Produces Antidepressant-Like Effect and Decreases Functional Activity and Protein Level of 5-HT2A Receptor in the Brain. Neuroscience 2018, 394, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Khomenko, T.; Tolstikova, T.; Bolkunov, A.; Dolgikh, M.; Pavlova, A.; Korchagina, D.; Volcho, K.P.; Salakhutdinov, N. 8-(Trifluoromethyl)-1,2,3,4,5-benzopentathiepin-6-amine: Novel Aminobenzopentathiepine having In Vivo Anticonvulsant and Anxiolytic Activities. Lett. Drug Des. Discov. 2009, 6, 464–467. [Google Scholar] [CrossRef]
- Xu, J.; Kurup, P.; Baguley, T.D.; Foscue, E.; Ellman, J.A.; Nairn, A.; Lombroso, P. Inhibition of the tyrosine phosphatase STEP61 restores BDNF expression and reverses motor and cognitive deficits in phencyclidine-treated mice. Cell. Mol. Life Sci. 2015, 73, 1503–1514. [Google Scholar] [CrossRef] [Green Version]
- Telegina, D.V.; E Korbolina, E.; I Ershov, N.; Kolosova, N.G.; Kozhevnikova, O.S. Identification of functional networks associated with cell death in the retina of OXYS rats during the development of retinopathy. Cell Cycle 2015, 14, 3544–3556. [Google Scholar] [CrossRef] [Green Version]
- Szabó, K.; Énzsöly, A.; Dékány, B.; Szabó, A.; Hajdú, R.I.; Radovits, T.; Mátyás, C.; Oláh, A.; Laurik, L.K.; Somfai, G.M.; et al. Histological Evaluation of Diabetic Neurodegeneration in the Retina of Zucker Diabetic Fatty (ZDF) Rats. Sci. Rep. 2017, 7, 1–17. [Google Scholar] [CrossRef]
- Telegina, D.V.; Kolosova, N.G.; Kozhevnikova, O.S. Immunohistochemical localization of NGF, BDNF, and their receptors in a normal and AMD-like rat retina. BMC Med. Genom. 2019, 12, 48. [Google Scholar] [CrossRef]
- Boulanger, L.M.; Lombroso, P.J.; Raghunathan, A.; During, M.J.; Wahle, P.; Naegele, J.R. Cellular and molecular characterization of a brain-enriched protein tyrosine phosphatase. J. Neurosci. 1995, 15, 1532–1544. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Olausson, P.; Venkitaramani, D.V.; Ruchkina, I.; Moran, T.D.; Tronson, N.C.; Mills, E.; Hakim, S.; Salter, M.W.; Taylor, J.R.; et al. The Striatal-Enriched Protein Tyrosine Phosphatase Gates Long-Term Potentiation and Fear Memory in the Lateral Amygdala. Boil. Psychiatry 2007, 61, 1049–1061. [Google Scholar] [CrossRef] [Green Version]
- Cases, S.; Saavedra, A.; Tyebji, S.; Giralt, A.; Alberch, J.; Perez-Navarro, E. Age-related changes in STriatal-Enriched protein tyrosine Phosphatase levels: Regulation by BDNF. Mol. Cell. Neurosci. 2018, 86, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Braithwaite, S.P.; Paul, S.; Nairn, A.C.; Lombroso, P.J. Synaptic plasticity: One STEP at a time. Trends Neurosci. 2006, 29, 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Kurup, P.; Foscue, E.; Lombroso, P.J. Striatal-enriched protein tyrosine phosphatase regulates the PTPα/Fyn signaling pathway. J. Neurochem. 2015, 134, 629–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castonguay, D.; Dufort-Gervais, J.; Menard, C.; Chatterjee, M.; Quirion, R.; Bontempi, B.; Schneider, J.S.; Arnsten, A.F.; Nairn, A.; Norris, C.M.; et al. The Tyrosine Phosphatase STEP Is Involved in Age-Related Memory Decline. Curr. Boil. 2018, 28, 1079–1089.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Xie, J.-W.; Yang, J.; Cao, Y.-P. Tyrosine phosphatase STEP61negatively regulates amyloid β-mediated ERK/CREB signaling pathways via α7 nicotinic acetylcholine receptors. J. Neurosci. Res. 2013, 91, 1581–1590. [Google Scholar] [CrossRef]
- Kozhevnikova, O.S.; Korbolina, E.E.; Ershov, N.I.; Kolosova, N.G. Rat retinal transcriptome: Effects of aging and AMD-like retinopathy. Cell Cycle 2013, 12, 1745–1761. [Google Scholar] [CrossRef] [Green Version]
- Stenkamp, D.L. Development of the Vertebrate Eye and Retina. Prog. Mol. Boil. Transl. Sci. 2015, 134, 397–414. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Kurup, P.; Bartos, J.A.; Patriarchi, T.; Hell, J.W.; Lombroso, P.J. Striatal-enriched Protein-tyrosine Phosphatase (STEP) Regulates Pyk2 Kinase Activity. J. Boil. Chem. 2012, 287, 20942–20956. [Google Scholar] [CrossRef] [Green Version]
- Kozhevnikova, O.S.; Telegina, D.V.; Devyatkin, V.A.; Kolosova, N.G. Involvement of the autophagic pathway in the progression of AMD-like retinopathy in senescence-accelerated OXYS rats. Biogerontology 2018, 19, 223–235. [Google Scholar] [CrossRef]
- Kozhevnikova, O.S.; Telegina, D.V.; Tyumentsev, M.A.; Kolosova, N.G. Disruptions of Autophagy in the Rat Retina with Age During the Development of Age-Related-Macular-Degeneration-like Retinopathy. Int. J. Mol. Sci. 2019, 20, 4804. [Google Scholar] [CrossRef] [Green Version]
- Rajagopal, S.; Deb, I.; Poddar, R.; Paul, S. Aging is associated with dimerization and inactivation of the brain-enriched tyrosine phosphatase STEP. Neurobiol. Aging 2016, 41, 25–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudnitskaya, E.A.; Kolosova, N.G.; Stefanova, N. Impact of changes in neurotrophic supplementation on development of Alzheimer’s disease-like pathology in OXYS rats. Biochemistry (Moscow) 2017, 82, 318–329. [Google Scholar] [CrossRef] [PubMed]
- Telegina, D.V.; Suvorov, G.K.; Kozhevnikova, O.S.; Kolosova, N.G. Mechanisms of Neuronal Death in the Cerebral Cortex during Aging and Development of Alzheimer’s Disease-Like Pathology in Rats. Int. J. Mol. Sci. 2019, 20, 5632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudnitskaya, E.A.; Kozlova, T.A.; Burnyasheva, A.O.; Kolosova, N.G.; Stefanova, N.A. Alterations of hippocampal neurogenesis during development of Alzheimer’s disease-like pathology in OXYS rats. Exp. Gerontol. 2019, 115, 32–45. [Google Scholar] [CrossRef]
- Xu, J.; Chatterjee, M.; Baguley, T.D.; Brouillette, J.; Kurup, P.; Ghosh, D.; Kanyo, J.; Zhang, Y.; Seyb, K.; Ononenyi, C.; et al. Inhibitor of the Tyrosine Phosphatase STEP Reverses Cognitive Deficits in a Mouse Model of Alzheimer’s Disease. PLoS Boil. 2014, 12, e1001923. [Google Scholar] [CrossRef] [Green Version]
- Lombroso, P.J.; Ogren, M.; Kurup, P.; Nairn, A.C. Molecular underpinnings of neurodegenerative disorders: Striatal-enriched protein tyrosine phosphatase signaling and synaptic plasticity. F1000Research 2016, 5, 2932. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Tikhonova, M.A.; Kulikova, E.A.; Volcho, K.P.; Khomenko, T.M.; Salakhutdinov, N.F.; Popova, N.K. Antidepressant Activity of 8-(trifluoromethyl)-1,2,3,4,5-benzopentathiepin- 6-amine hydrochloride (TC-2153): Comparison with Classical Antidepressants. Lett. Drug Des. Discov. 2013, 11, 169–173. [Google Scholar] [CrossRef]
- Xu, J.; Kurup, P.; Nairn, A.C.; Lombroso, P.J. Synaptic NMDA Receptor Activation Induces Ubiquitination and Degradation of STEP61. Mol. Neurobiol. 2017, 55, 3096–3111. [Google Scholar] [CrossRef]
- Karasawa, T.; Lombroso, P.J. Disruption of striatal-enriched protein tyrosine phosphatase (STEP) function in neuropsychiatric disorders. Neurosci. Res. 2014, 89, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Olausson, P.; Venkitaramani, D.V.; Moran, T.D.; Salter, M.W.; Taylor, J.R.; Lombroso, P.J. The tyrosine phosphatase STEP constrains amygdala-dependent memory formation and neuroplasticity. Neuroscience 2012, 225, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Telegina, D.V.; Kozhevnikova, O.S.; Kolosova, N.G. Molecular mechanisms of cell death in retina during development of age-related macular degeneration. Adv. Gerontol. 2017, 7, 17–24. [Google Scholar] [CrossRef]
- Kothmann, W.W.; Trexler, E.B.; Whitaker, C.M.; Li, W.; Massey, S.C.; O’Brien, J. Nonsynaptic NMDA receptors mediate activity-dependent plasticity of gap junctional coupling in the AII amacrine cell network. J. Neurosci. 2012, 32, 6747–6759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowie, D. Redefining the classification of AMPA-selective ionotropic glutamate receptors. J. Physiol. 2011, 590, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Veruki, M.L.; Zhou, Y.; Castilho, Á.; Morgans, C.W.; Hartveit, E. Extrasynaptic NMDA Receptors on Rod Pathway Amacrine Cells: Molecular Composition, Activation, and Signaling. J. Neurosci. 2018, 39, 627–650. [Google Scholar] [CrossRef]
- Telegina, D.V.; Kozhevnikova, O.S.; Kolosova, N.G. Changes in Retinal Glial Cells with Age and during Development of Age-Related Macular Degeneration. Biochemistry (Moscow) 2018, 83, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Kulikov, A.V.; Tikhonova, M.A.; Kulikova, E.A.; Volcho, K.P.; Khomenko, T.M.; Salakhutdinov, N.F.; Popova, N.K. A new synthetic varacin analogue, 8-(trifluoromethyl)-1,2,3,4,5-benzopentathiepin-6-amine hydrochloride (TC-2153), decreased hereditary catalepsy and increased the BDNF gene expression in the hippocampus in mice. Psychopharmacology 2011, 221, 469–478. [Google Scholar] [CrossRef]
- Li, L.; Shi, L.; Xu, Y.-M.; Yang, X.; Suo, Z.-W.; Hu, X.-D. GABAergic Inhibition Regulated Pain Sensitization through STEP61 Signaling in Spinal Dorsal Horn of Mice. Anesthesiology 2015, 122, 686–697. [Google Scholar] [CrossRef]
- Mota, S.; Ferreira, I.; Rego, A.C. Dysfunctional synapse in Alzheimer’s disease—A focus on NMDA receptors. Neuropharmacology 2014, 76, 16–26. [Google Scholar] [CrossRef]
- Wang, R.; Reddy, P.H. Role of Glutamate and NMDA Receptors in Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 57, 1041–1048. [Google Scholar] [CrossRef] [Green Version]
- Gojon, G.; A Morales, G. SG1002 and Catenated Divalent Organic Sulfur Compounds as Promising Hydrogen Sulfide Prodrugs. Antioxid. Redox Signal. 2020. [Google Scholar] [CrossRef]
- Kulikov, A.V.; Naumenko, V.S.; Voronova, I.P.; Tikhonova, M.A.; Popova, N.K. Quantitative RT-PCR assay of 5-HT1A and 5-HT2A serotonin receptor mRNAs using genomic DNA as an external standard. J. Neurosci. Methods 2005, 141, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Naumenko, V.S.; Osipova, D.V.; Kostina, E.V.; Kulikov, A.V. Utilization of a two-standard system in real-time PCR for quantification of gene expression in the brain. J. Neurosci. Methods 2008, 170, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, E.A.; Fursenko, D.V.; Bazhenova, E.Y.; Kulikov, A.V. Pargyline and p-Chlorophenylalanine Decrease Expression of Ptpn5 Encoding Striatal-Enriched Protein Tyrosine Phosphatase (STEP) in the Mouse Striatum. Mol. Boil. 2020, 54, 274–280. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Wistar | OXYS | OXYS TC-2153 | F, p | |
|---|---|---|---|---|
| Total phosphatase activity, nmole/(mg·min) | 0.997 ± 0.039 | 0.986 ± 0.031 | 1.044 ± 0.038 | F2,21 = 0.74 |
| STEP phosphatase activity, nmole/(mg·min) | 0.729 ± 0.036 | 0.720 ± 0.031 | 0.773 ± 0.028 | F2,21 = 0.80 |
| Ptpn5 mRNA level, fold change relative to Wistar rats | 1.00 ± 0.070 | 1.052 ± 0.119 | 1.062 ± 0.083 | F2,21 = 0.13 |
| STEP46 protein level, a.u. | 92.24 ± 17.98 | 92.13 ± 14.32 | 118.18 ± 20.54 | F2,15 = 0.71 |
| STEP61 protein level, a.u. | 89.04 ± 10.41 | 97.26 ± 12.52 | 101.38 ± 10.23 | F2,23 = 0.33 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Telegina, D.V.; Kulikova, E.A.; Kozhevnikova, O.S.; Kulikov, A.V.; Khomenko, T.M.; Volcho, K.P.; Salakhutdinov, N.F.; Kolosova, N.G. Alterations of STEP46 and STEP61 Expression in the Rat Retina with Age and AMD-Like Retinopathy Development. Int. J. Mol. Sci. 2020, 21, 5182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155182
Telegina DV, Kulikova EA, Kozhevnikova OS, Kulikov AV, Khomenko TM, Volcho KP, Salakhutdinov NF, Kolosova NG. Alterations of STEP46 and STEP61 Expression in the Rat Retina with Age and AMD-Like Retinopathy Development. International Journal of Molecular Sciences. 2020; 21(15):5182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155182
Chicago/Turabian StyleTelegina, Darya V., Elizabeth A. Kulikova, Oyuna S. Kozhevnikova, Alexander V. Kulikov, Tatyana M. Khomenko, Konstantin P. Volcho, Nariman F. Salakhutdinov, and Nataliya G. Kolosova. 2020. "Alterations of STEP46 and STEP61 Expression in the Rat Retina with Age and AMD-Like Retinopathy Development" International Journal of Molecular Sciences 21, no. 15: 5182. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155182