Regulation of Drosophila Hematopoiesis in Lymph Gland: From a Developmental Signaling Point of View


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Drosophila Blood Cell Types and Functions
3. Overview of Drosophila Hematopoiesis from Embryos to Adults
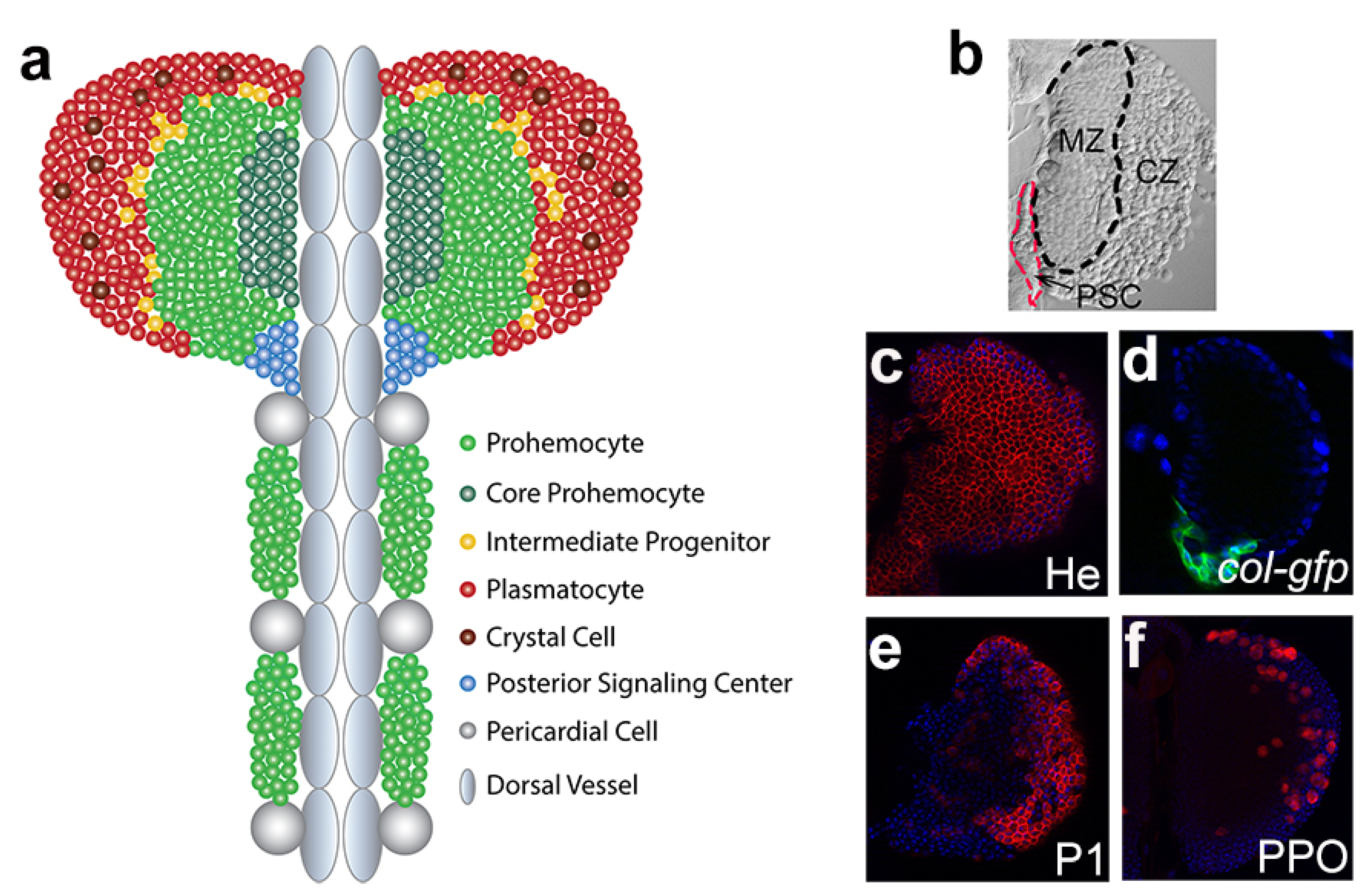
4. Drosophila Lymph Gland
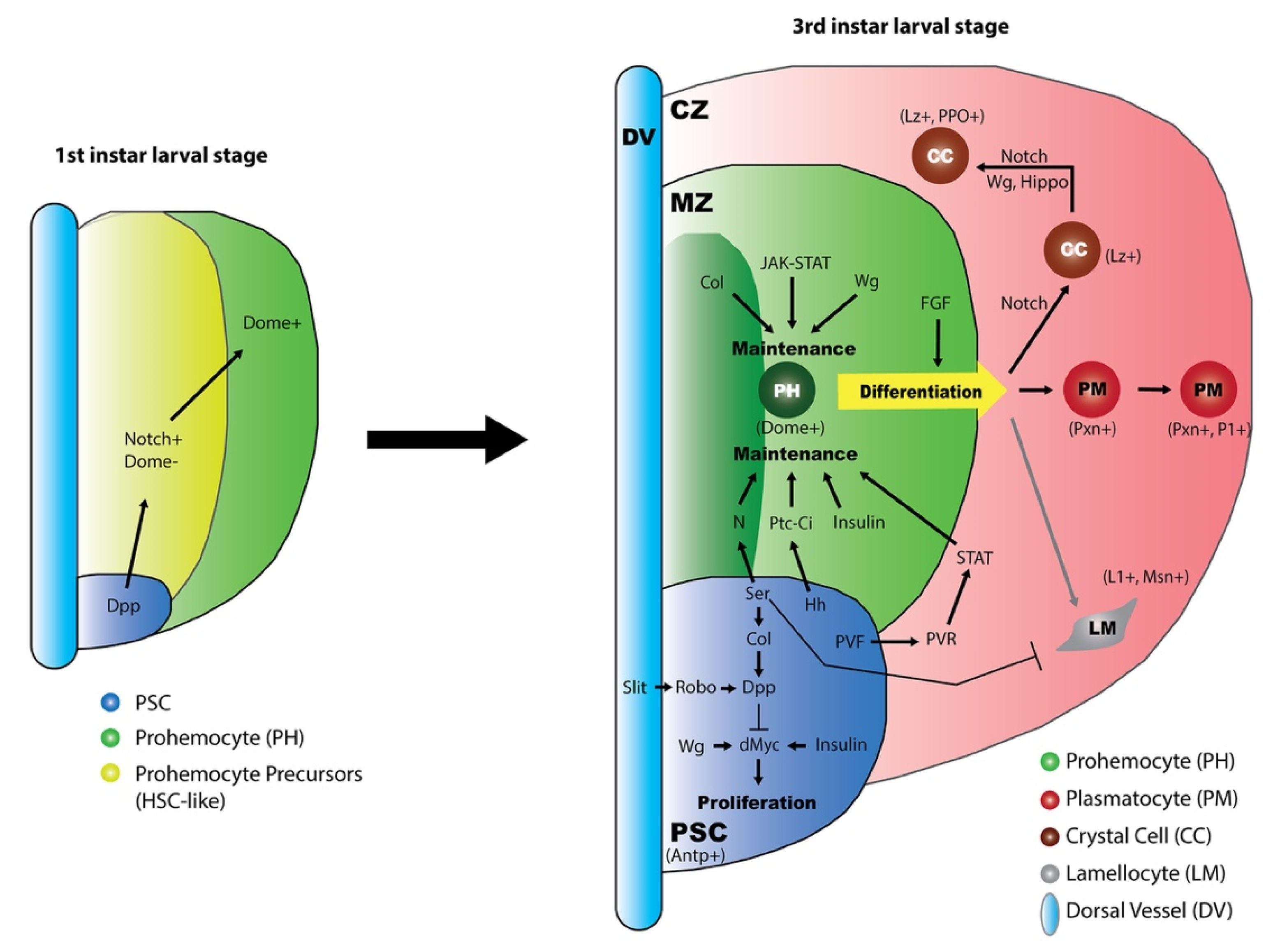
4.1. Lymph Gland Zones
4.2. Role of the PSC in the Lymph Gland
5. Regulatory Signaling during Lymph Gland Development
5.1. Notch Signaling Instructs Crystal Cell Fate Determination
5.2. Hh Signaling Maintains the Prohemocyte Potency
5.3. Wnt/Wg Signaling Modulates Hemocyte Proliferation and Differentiation
5.4. JAK-STAT Signaling Inhibits Prohemocyte Differentiation
5.5. BMP/Dpp Signaling Restricts PSC Size
6. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Bejsovec, A. Wingless Signaling: A Genetic Journey from Morphogenesis to Metastasis. Genetics 2018, 208, 1311–1336. [Google Scholar] [CrossRef] [Green Version]
- Kong, J.H.; Siebold, C.; Rohatgi, R. Biochemical mechanisms of vertebrate hedgehog signaling. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siebel, C.; Lendahl, U. Notch Signaling in Development, Tissue Homeostasis, and Disease. Physiol. Rev. 2017, 97, 1235–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massagué, J. TGFβ signalling in context. Nat. Rev. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar] [CrossRef]
- Grigorian, M.; Mandal, L.; Hartenstein, V. Hematopoiesis at the onset of metamorphosis: Terminal differentiation and dissociation of the Drosophila lymph gland. Dev. Genes Evol. 2011, 221, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Letourneau, M.; Lapraz, F.; Sharma, A.; Vanzo, N.; Waltzer, L.; Crozatier, M. Drosophila hematopoiesis under normal conditions and in response to immune stress. FEBS Lett. 2016, 590, 4034–4051. [Google Scholar] [CrossRef]
- Banerjee, U.; Girard, J.R.; Goins, L.M.; Spratford, C.M. Drosophila as a Genetic Model for Hematopoiesis. Genetics 2019, 211, 367–417. [Google Scholar] [CrossRef] [Green Version]
- Shim, J. Drosophila blood as a model system for stress sensing mechanisms. BMB Rep. 2015, 48, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Lebestky, T.; Chang, T.; Hartenstein, V.; Banerjee, U. Specification of Drosophila hematopoietic lineage by conserved transcription factors. Science 2000, 288, 146–149. [Google Scholar] [CrossRef]
- Hartenstein, V. Blood cells and blood cell development in the animal kingdom. Annu. Rev. Cell Dev. Biol. 2006, 22, 677–712. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Singh, A.; Mandal, S.; Mandal, L. Active hematopoietic hubs in Drosophila adults generate hemocytes and contribute to immune response. Dev. Cell 2015, 33, 478–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leitão, A.B.; Sucena, É. Drosophila sessile hemocyte clusters are true hematopoietic tissues that regulate larval blood cell differentiation. eLife 2015, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunt, S.; Hooley, C.; Hu, N.; Scahill, C.; Weavers, H.; Skaer, H. Hemocyte-secreted type IV collagen enhances BMP signaling to guide renal tubule morphogenesis in Drosophila. Dev. Cell 2010, 19, 296–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olofsson, B.; Page, D.T. Condensation of the central nervous system in embryonic Drosophila is inhibited by blocking hemocyte migration or neural activity. Dev. Biol. 2005, 279, 233–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van De Bor, V.; Zimniak, G.; Papone, L.; Cerezo, D.; Malbouyres, M.; Juan, T.; Ruggiero, F.; Noselli, S. Companion Blood Cells Control Ovarian Stem Cell Niche Microenvironment and Homeostasis. Cell Rep. 2015, 13, 546–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudzic, J.P.; Kondo, S.; Ueda, R.; Bergman, C.M.; Lemaitre, B. Drosophila innate immunity: Regional and functional specialization of prophenoloxidases. BMC Biol. 2015, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binggeli, O.; Neyen, C.; Poidevin, M.; Lemaitre, B. Prophenoloxidase activation is required for survival to microbial infections in Drosophila. PLoS Pathog. 2014, 10, e1004067. [Google Scholar] [CrossRef]
- Gajewski, K.M.; Sorrentino, R.P.; Lee, J.H.; Zhang, Q.; Russell, M.; Schulz, R.A. Identification of a crystal cell-specific enhancer of the black cells prophenoloxidase gene in Drosophila. Genesis 2007, 45, 200–207. [Google Scholar] [CrossRef]
- Lanot, R.; Zachary, D.; Holder, F.; Meister, M. Postembryonic Hematopoiesis in Drosophila. Dev. Biol. 2001, 230, 243–257. [Google Scholar] [CrossRef] [Green Version]
- Markus, R.; Laurinyecz, B.; Kurucz, E.; Honti, V.; Bajusz, I.; Sipos, B.; Somogyi, K.; Kronhamn, J.; Hultmark, D.; Ando, I. Sessile hemocytes as a hematopoietic compartment in Drosophila melanogaster. Proc. Acad. Natl. Sci. USA 2009, 106, 4805–4809. [Google Scholar] [CrossRef] [Green Version]
- Honti, V.; Csordas, G.; Markus, R.; Kurucz, E.; Jankovics, F.; Ando, I. Cell lineage tracing reveals the plasticity of the hemocyte lineages and of the hematopoietic compartments in Drosophila melanogaster. Mol. Immunol. 2010, 47, 1997–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makhijani, K.; Alexander, B.; Tanaka, T.; Rulifson, E.; Bruckner, K. The peripheral nervous system supports blood cell homing and survival in the Drosophila larva. Development 2011, 138, 5379–5391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makhijani, K.; Alexander, B.; Rao, D.; Petraki, S.; Herboso, L.; Kukar, K.; Batool, I.; Wachner, S.; Gold, K.S.; Wong, C.; et al. Regulation of Drosophila hematopoietic sites by Activin-beta from active sensory neurons. Nat. Commun. 2017, 8, 15990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzemien, J.; Oyallon, J.; Crozatier, M.; Vincent, A. Hematopoietic progenitors and hemocyte lineages in the Drosophila lymph gland. Dev. Biol. 2010, 346, 310–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crozatier, M.; Ubeda, J.M.; Vincent, A.; Meister, M. Cellular immune response to parasitization in Drosophila requires the EBF orthologue collier. PLoS Biol. 2004, 2, E196. [Google Scholar] [CrossRef] [Green Version]
- Krzemien, J.; Dubois, L.; Makki, R.; Meister, M.; Vincent, A.; Crozatier, M. Control of blood cell homeostasis in Drosophila larvae by the posterior signalling centre. Nature 2007, 446, 325–328. [Google Scholar] [CrossRef]
- Stofanko, M.; Kwon, S.Y.; Badenhorst, P. Lineage Tracing of Lamellocytes Demonstrates Drosophila Macrophage Plasticity. PLoS ONE 2010, 5, e14051. [Google Scholar] [CrossRef]
- Anderl, I.; Vesala, L.; Ihalainen, T.O.; Vanha-aho, L.-M.; Andó, I.; Rämet, M.; Hultmark, D. Transdifferentiation and Proliferation in Two Distinct Hemocyte Lineages in Drosophila melanogaster Larvae after Wasp Infection. PLoS Pathog. 2016, 12, e1005746. [Google Scholar] [CrossRef]
- Holz, A.; Bossinger, B.; Strasser, T.; Janning, W.; Klapper, R. The two origins of hemocytes in Drosophila. Development 2003, 130, 4955–4962. [Google Scholar] [CrossRef] [Green Version]
- Sanchez Bosch, P.; Makhijani, K.; Herboso, L.; Gold, K.S.; Baginsky, R.; Woodcock, K.J.; Alexander, B.; Kukar, K.; Corcoran, S.; Jacobs, T.; et al. Adult Drosophila Lack Hematopoiesis but Rely on a Blood Cell Reservoir at the Respiratory Epithelia to Relay Infection Signals to Surrounding Tissues. Dev. Cell 2019, 51, 787–803.e5. [Google Scholar] [CrossRef]
- Mackenzie, D.K.; Bussiere, L.F.; Tinsley, M.C. Senescence of the cellular immune response in Drosophila melanogaster. Exp. Gerontol. 2011, 46, 853–859. [Google Scholar] [CrossRef] [PubMed]
- El Shatoury, H.H. The structure of the lymph glands of Drosophila larvae. Wilhelm Roux Arch. Entwickl. Mech. Org. 1955, 147, 489–495. [Google Scholar] [CrossRef]
- Jung, S.H.; Evans, C.J.; Uemura, C.; Banerjee, U. The Drosophila lymph gland as a developmental model of hematopoiesis. Development 2005, 132, 2521–2533. [Google Scholar] [CrossRef] [Green Version]
- Mandal, L.; Martinez-Agosto, J.A.; Evans, C.J.; Hartenstein, V.; Banerjee, U. A Hedgehog- and Antennapedia-dependent niche maintains Drosophila haematopoietic precursors. Nature 2007, 446, 320–324. [Google Scholar] [CrossRef]
- Oyallon, J.; Vanzo, N.; Krzemien, J.; Morin-Poulard, I.; Vincent, A.; Crozatier, M. Two Independent Functions of Collier/Early B Cell Factor in the Control of Drosophila Blood Cell Homeostasis. PLoS ONE 2016, 11, e0148978. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Obregon, D.; Katz, M.J.; Durrieu, L.; Gandara, L.; Wappner, P. Context-specific functions of Notch in Drosophila blood cell progenitors. Dev. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Dey, N.S.; Ramesh, P.; Chugh, M.; Mandal, S.; Mandal, L. Dpp dependent Hematopoietic stem cells give rise to Hh dependent blood progenitors in larval lymph gland of Drosophila. elife 2016, 5. [Google Scholar] [CrossRef]
- Minakhina, S.; Steward, R. Hematopoietic stem cells in Drosophila. Development 2010, 137, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinenko, S.A.; Mandal, L.; Martinez-Agosto, J.A.; Banerjee, U. Dual role of wingless signaling in stem-like hematopoietic precursor maintenance in Drosophila. Dev. Cell 2009, 16, 756–763. [Google Scholar] [CrossRef] [Green Version]
- Lebestky, T.; Jung, S.H.; Banerjee, U. A Serrate-expressing signaling center controls Drosophila hematopoiesis. Genes Dev. 2003, 17, 348–353. [Google Scholar] [CrossRef] [Green Version]
- Pennetier, D.; Oyallon, J.; Morin-Poulard, I.; Dejean, S.; Vincent, A.; Crozatier, M. Size control of the Drosophila hematopoietic niche by bone morphogenetic protein signaling reveals parallels with mammals. Proc. Natl. Acad. Sci. USA 2012, 109, 3389–3394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, B.C.; Mukherjee, T.; Mandal, L.; Evans, C.J.; Sinenko, S.A.; Martinez-Agosto, J.A.; Banerjee, U. Interaction between differentiating cell- and niche-derived signals in hematopoietic progenitor maintenance. Cell 2011, 147, 1589–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benmimoun, B.; Polesello, C.; Haenlin, M.; Waltzer, L. The EBF transcription factor Collier directly promotes Drosophila blood cell progenitor maintenance independently of the niche. Proc. Natl. Acad. Sci. USA 2015, 112, 9052–9057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldeosingh, R.; Gao, H.; Wu, X.; Fossett, N. Hedgehog signaling from the Posterior Signaling Center maintains U-shaped expression and a prohemocyte population in Drosophila. Dev. Biol. 2018, 441, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Sinenko, S.A.; Shim, J.; Banerjee, U. Oxidative stress in the haematopoietic niche regulates the cellular immune response in Drosophila. EMBO Rep. 2011, 13, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Louradour, I.; Sharma, A.; Morin-Poulard, I.; Letourneau, M.; Vincent, A.; Crozatier, M.; Vanzo, N. Reactive oxygen species-dependent Toll/NF-kappaB activation in the Drosophila hematopoietic niche confers resistance to wasp parasitism. elife 2017, 6. [Google Scholar] [CrossRef]
- Tokusumi, Y.; Tokusumi, T.; Schulz, R.A. Mechanical stress to Drosophila larvae stimulates a cellular immune response through the JAK/STAT signaling pathway. Biochem. Biophys. Res. Commun. 2018, 502, 415–421. [Google Scholar] [CrossRef]
- Kopan, R.; Ilagan, M.X. The canonical Notch signaling pathway: Unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar] [CrossRef] [Green Version]
- Small, C.; Ramroop, J.; Otazo, M.; Huang, L.H.; Saleque, S.; Govind, S. An unexpected link between notch signaling and ROS in restricting the differentiation of hematopoietic progenitors in Drosophila. Genetics 2014, 197, 471–483. [Google Scholar] [CrossRef] [Green Version]
- Duvic, B.; Hoffmann, J.A.; Meister, M.; Royet, J. Notch signaling controls lineage specification during Drosophila larval hematopoiesis. Curr. Biol. 2002, 12, 1923–1927. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, G.B.; Martinez-Agosto, J.A. Yorkie and Scalloped signaling regulates Notch-dependent lineage specification during Drosophila hematopoiesis. Curr. Biol. 2014, 24, 2665–2672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terriente-Felix, A.; Li, J.; Collins, S.; Mulligan, A.; Reekie, I.; Bernard, F.; Krejci, A.; Bray, S. Notch cooperates with Lozenge/Runx to lock haemocytes into a differentiation programme. Development 2013, 140, 926–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, T.; Kim, W.S.; Mandal, L.; Banerjee, U. Interaction between Notch and Hif-alpha in development and survival of Drosophila blood cells. Science 2011, 332, 1210–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Wang, L.; Chi, C.; Lan, W.; Su, Y. The emerging roles of phosphatases in Hedgehog pathway. Cell Commun. Signal. 2017, 15, 35. [Google Scholar] [CrossRef] [Green Version]
- Giordani, G.; Barraco, M.; Giangrande, A.; Martinelli, G.; Guadagnuolo, V.; Simonetti, G.; Perini, G.; Bernardoni, R. The human Smoothened inhibitor PF-04449913 induces exit from quiescence and loss of multipotent Drosophila hematopoietic progenitor cells. Oncotarget 2016, 7, 55313–55327. [Google Scholar] [CrossRef] [Green Version]
- Tokusumi, T.; Tokusumi, Y.; Schulz, R.A. The mir-7 and bag of marbles genes regulate Hedgehog pathway signaling in blood cell progenitors in Drosophila larval lymph glands. Genesis 2018, 56, e23210. [Google Scholar] [CrossRef]
- Tokusumi, Y.; Tokusumi, T.; Stoller-Conrad, J.; Schulz, R.A. Serpent, suppressor of hairless and U-shaped are crucial regulators of hedgehog niche expression and prohemocyte maintenance during Drosophila larval hematopoiesis. Development 2010, 137, 3561–3568. [Google Scholar] [CrossRef] [Green Version]
- Hooper, J.E.; Scott, M.P. Communicating with Hedgehogs. Nat. Rev. Mol. Cell Biol. 2005, 6, 306–317. [Google Scholar] [CrossRef]
- Hao, Y.; Jin, L.H. Dual role for Jumu in the control of hematopoietic progenitors in the Drosophila lymph gland. elife 2017, 6. [Google Scholar] [CrossRef]
- Zhang, C.U.; Blauwkamp, T.A.; Burby, P.E.; Cadigan, K.M. Wnt-mediated repression via bipartite DNA recognition by TCF in the Drosophila hematopoietic system. PLoS Genet. 2014, 10, e1004509. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.U.; Cadigan, K.M. The matrix protein Tiggrin regulates plasmatocyte maturation in Drosophila larva. Development 2017, 144, 2415–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi, S.; Starz-Gaiano, M. Drosophila Jak/STAT Signaling: Regulation and Relevance in Human Cancer and Metastasis. Int. J. Mol. Sci. 2018, 19, 4056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin-Poulard, I.; Vincent, A.; Crozatier, M. The Drosophila JAK-STAT pathway in blood cell formation and immunity. JAKSTAT 2013, 2, e25700. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Kronhamn, J.; Ekstrom, J.O.; Korkut, G.G.; Hultmark, D. JAK/STAT signaling in Drosophila muscles controls the cellular immune response against parasitoid infection. EMBO Rep. 2015, 16, 1664–1672. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Wu, X.; Fossett, N. Upregulation of the Drosophila Friend of GATA gene U-shaped by JAK/STAT signaling maintains lymph gland prohemocyte potency. Mol. Cell Biol. 2009, 29, 6086–6096. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, G.B.; Martinez-Agosto, J.A. The TEAD family transcription factor Scalloped regulates blood progenitor maintenance and proliferation in Drosophila through PDGF/VEGFR receptor (Pvr) signaling. Dev. Biol. 2017, 425, 21–32. [Google Scholar] [CrossRef]
- Sinha, A.; Khadilkar, R.J.; Vinay, K.S.; Roychowdhury Sinha, A.; Inamdar, M.S. Conserved regulation of the Jak/STAT pathway by the endosomal protein asrij maintains stem cell potency. Cell Rep. 2013, 4, 649–658. [Google Scholar] [CrossRef] [Green Version]
- Sorrentino, R.P.; Tokusumi, T.; Schulz, R.A. The Friend of GATA protein U-shaped functions as a hematopoietic tumor suppressor in Drosophila. Dev. Biol. 2007, 311, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Terriente-Félix, A.; Pérez, L.; Bray, S.J.; Nebreda, A.R.; Milán, M. A Drosophila model of myeloproliferative neoplasm reveals a feed-forward loop in the JAK pathway mediated by p38 MAPK signalling. Dis. Mod. Mech. 2017, 10, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Minakhina, S.; Tan, W.; Steward, R. JAK/STAT and the GATA factor Pannier control hemocyte maturation and differentiation in Drosophila. Dev. Biol. 2011, 352, 308–316. [Google Scholar] [CrossRef]
- Khadilkar, R.J.; Rodrigues, D.; Mote, R.D.; Sinha, A.R.; Kulkarni, V.; Magadi, S.S.; Inamdar, M.S. ARF1-GTP regulates Asrij to provide endocytic control of Drosophila blood cell homeostasis. Proc. Natl. Acad. Sci. USA 2014, 111, 4898–4903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamaratoglu, F.; Affolter, M.; Pyrowolakis, G. Dpp/BMP signaling in flies: From molecules to biology. Semin. Cell Dev. Biol. 2014, 32, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin-Poulard, I.; Sharma, A.; Louradour, I.; Vanzo, N.; Vincent, A.; Crozatier, M. Vascular control of the Drosophila haematopoietic microenvironment by Slit/Robo signalling. Nat. Commun. 2016, 7, 11634. [Google Scholar] [CrossRef] [Green Version]
- Dragojlovic-Munther, M.; Martinez-Agosto, J.A. Extracellular matrix-modulated Heartless signaling in Drosophila blood progenitors regulates their differentiation via a Ras/ETS/FOG pathway and target of rapamycin function. Dev. Biol. 2013, 384, 313–330. [Google Scholar] [CrossRef] [Green Version]
- Milton, C.C.; Grusche, F.A.; Degoutin, J.L.; Yu, E.; Dai, Q.; Lai, E.C.; Harvey, K.F. The Hippo pathway regulates hematopoiesis in Drosophila melanogaster. Curr. Biol. 2014, 24, 2673–2680. [Google Scholar] [CrossRef] [Green Version]
- Khadilkar, R.J.; Tanentzapf, G. Septate junction components control Drosophila hematopoiesis through the Hippo pathway. Development 2019, 146. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, G.B.; Martinez-Agosto, J.A. Kicking it up a Notch for the best in show: Scalloped leads Yorkie into the haematopoietic arena. Fly (Austin) 2014, 8, 206–217. [Google Scholar] [CrossRef] [Green Version]
- Benmimoun, B.; Polesello, C.; Waltzer, L.; Haenlin, M. Dual role for Insulin/TOR signaling in the control of hematopoietic progenitor maintenance in Drosophila. Development 2012, 139, 1713–1717. [Google Scholar] [CrossRef] [Green Version]
- Tokusumi, Y.; Tokusumi, T.; Shoue, D.A.; Schulz, R.A. Gene Regulatory Networks Controlling Hematopoietic Progenitor Niche Cell Production and Differentiation in the Drosophila Lymph Gland. PLoS ONE 2012, 7, e41604. [Google Scholar] [CrossRef] [Green Version]
- Lam, V.; Tokusumi, T.; Tokusumi, Y.; Schulz, R.A. bantam miRNA is important for Drosophila blood cell homeostasis and a regulator of proliferation in the hematopoietic progenitor niche. Biochem. Biophys. Res. Commun. 2014, 453, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Tokusumi, T.; Tokusumi, Y.; Hopkins, D.W.; Schulz, R.A. Bag of Marbles controls the size and organization of the Drosophila hematopoietic niche through interactions with the Insulin-like growth factor pathway and Retinoblastoma-family protein. Development 2015, 142, 2261–2267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, H.; Sharma, S.K.; Mandal, S.; Mandal, L. Lar maintains the homeostasis of the hematopoietic organ in Drosophila by regulating insulin signaling in the niche. Development 2019, 146, dev178202. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.; Mukherjee, T.; Banerjee, U. Direct sensing of systemic and nutritional signals by haematopoietic progenitors in Drosophila. Nat. Cell Biol. 2012, 14, 394–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragojlovic-Munther, M.; Martinez-Agosto, J.A. Multifaceted roles of PTEN and TSC orchestrate growth and differentiation of Drosophila blood progenitors. Development 2012, 139, 3752–3763. [Google Scholar] [CrossRef] [Green Version]
- Kudo, T.; Jeknic, S.; Macklin, D.N.; Akhter, S.; Hughey, J.J.; Regot, S.; Covert, M.W. Live-cell measurements of kinase activity in single cells using translocation reporters. Nat. Protoc. 2018, 13, 155–169. [Google Scholar] [CrossRef]
- Viswanathan, R.; Necakov, A.; Trylinski, M.; Harish, R.K.; Krueger, D.; Esposito, E.; Schweisguth, F.; Neveu, P.; De Renzis, S. Optogenetic inhibition of Delta reveals digital Notch signalling output during tissue differentiation. EMBO Rep. 2019, 20, e47999. [Google Scholar] [CrossRef]
- Johnson, H.E.; Goyal, Y.; Pannucci, N.L.; Schupbach, T.; Shvartsman, S.Y.; Toettcher, J.E. The Spatiotemporal Limits of Developmental Erk Signaling. Dev. Cell 2017, 40, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Johnson, H.E.; Toettcher, J.E. Signaling Dynamics Control Cell Fate in the Early Drosophila Embryo. Dev. Cell 2019, 48, 361–370.e3. [Google Scholar] [CrossRef] [Green Version]
- Guglielmi, G.; Barry, J.D.; Huber, W.; De Renzis, S. An Optogenetic Method to Modulate Cell Contractility during Tissue Morphogenesis. Dev. Cell 2015, 35, 646–660. [Google Scholar] [CrossRef] [Green Version]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, W.; Liu, S.; Zhao, L.; Su, Y. Regulation of Drosophila Hematopoiesis in Lymph Gland: From a Developmental Signaling Point of View. Int. J. Mol. Sci. 2020, 21, 5246. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155246
Lan W, Liu S, Zhao L, Su Y. Regulation of Drosophila Hematopoiesis in Lymph Gland: From a Developmental Signaling Point of View. International Journal of Molecular Sciences. 2020; 21(15):5246. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155246
Chicago/Turabian StyleLan, Wenwen, Sumin Liu, Long Zhao, and Ying Su. 2020. "Regulation of Drosophila Hematopoiesis in Lymph Gland: From a Developmental Signaling Point of View" International Journal of Molecular Sciences 21, no. 15: 5246. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155246