Cell Sheets from Adipose Tissue MSC Induce Healing of Pressure Ulcer and Prevent Fibrosis via Trigger Effects on Granulation Tissue Growth and Vascularization

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
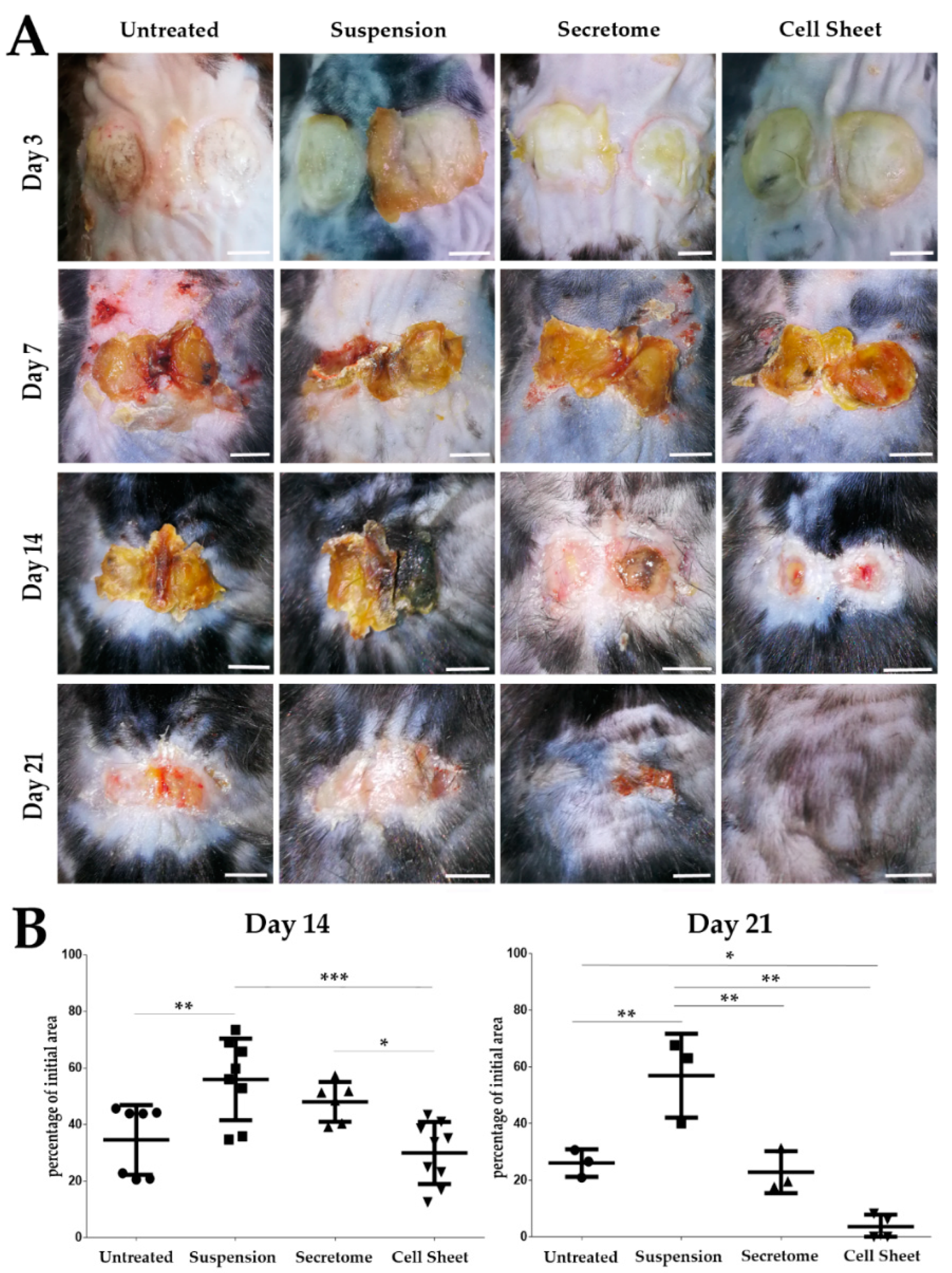
2.1. MSC Sheet Transplantation Accelerates Healing of Pressure Ulcer Defect
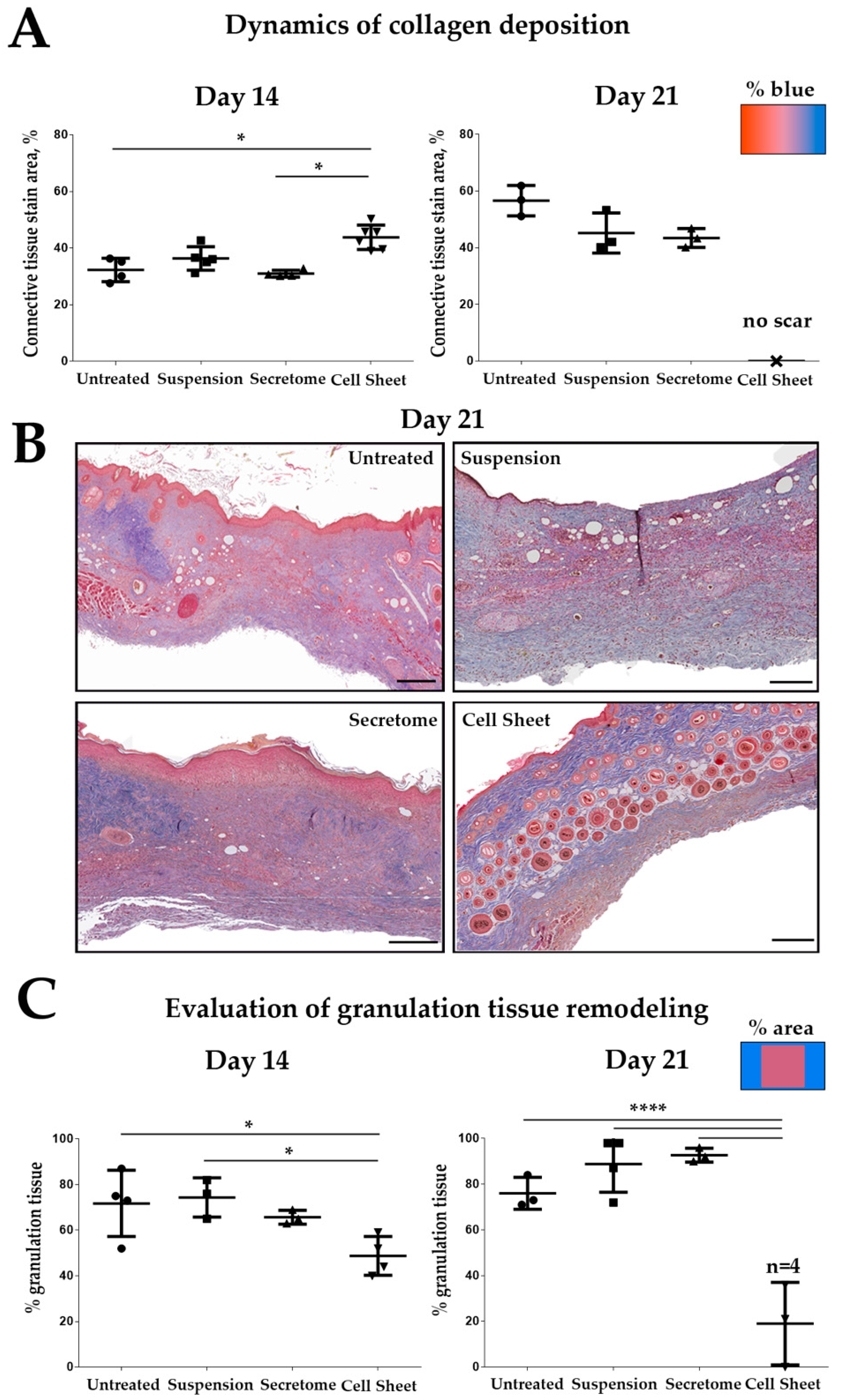
2.2. Cell Sheet Transplantation Induces Intensive Remodeling of Granulation Tissue
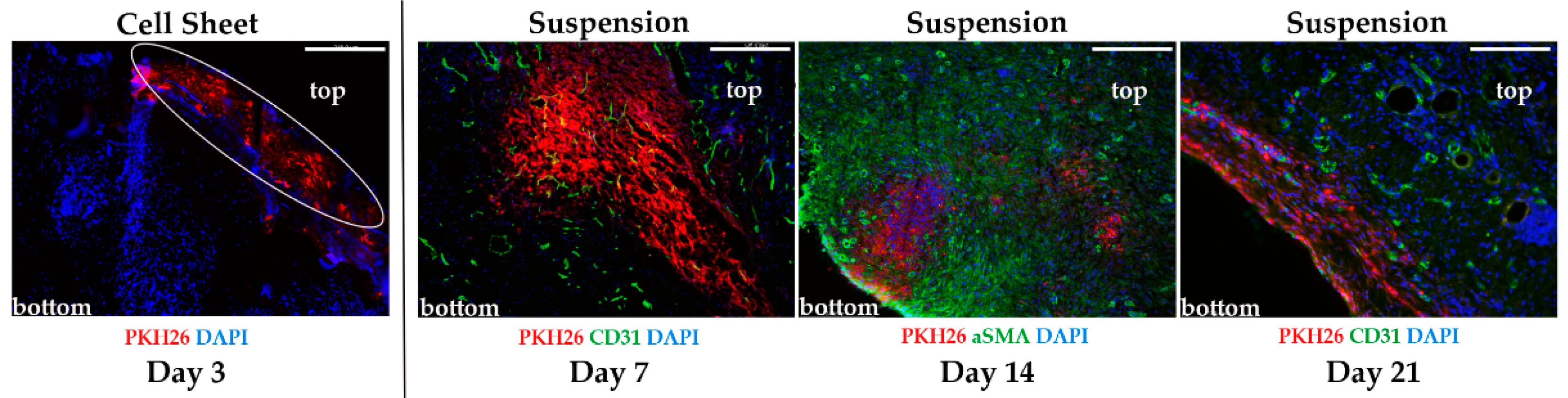
2.3. Evaluation of Transplant Retention after MSC Delivery by Suspension or Cell Sheets
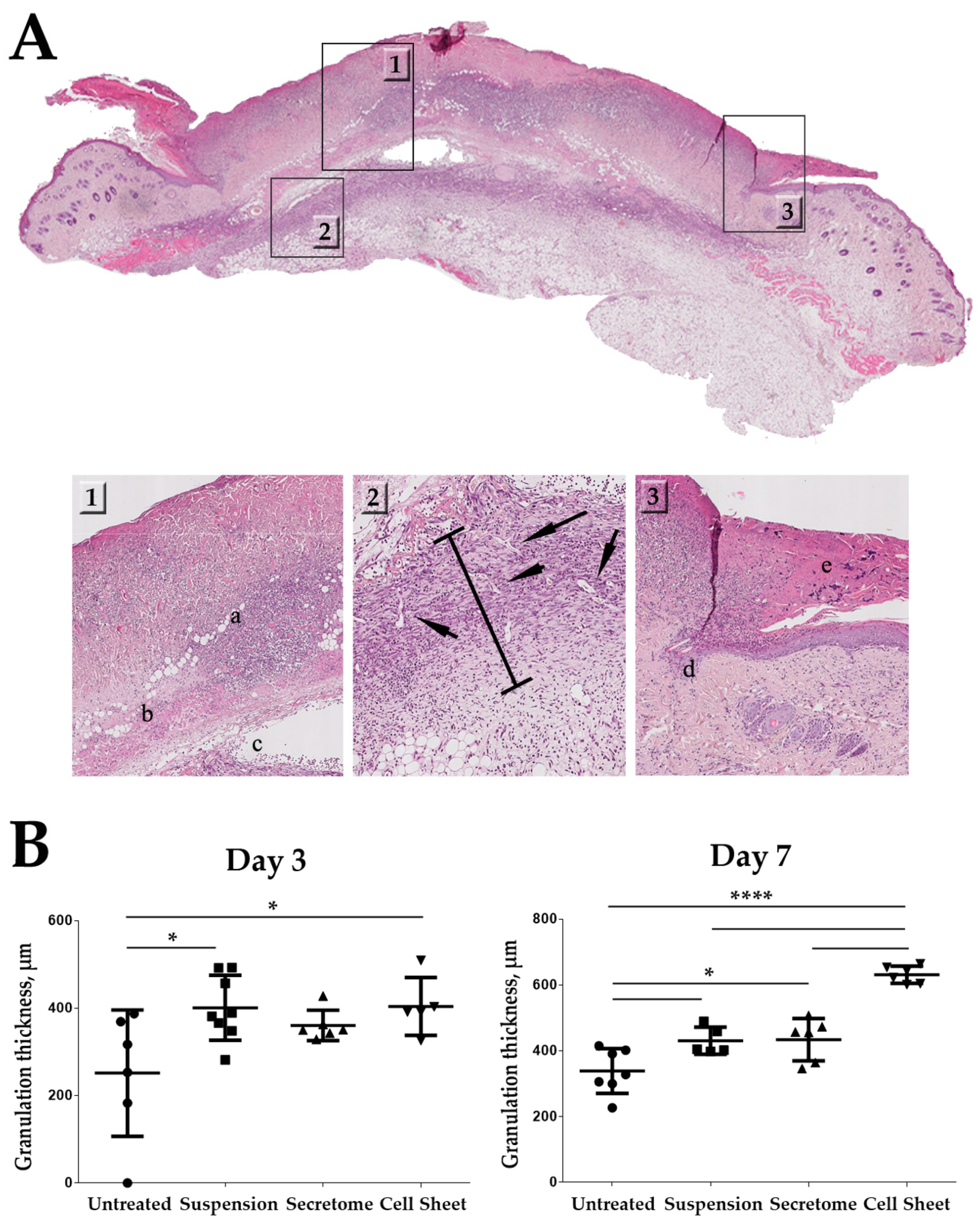
2.4. Cell Sheets Promote Formation of Granulation Tissue at Early Stages of Healing
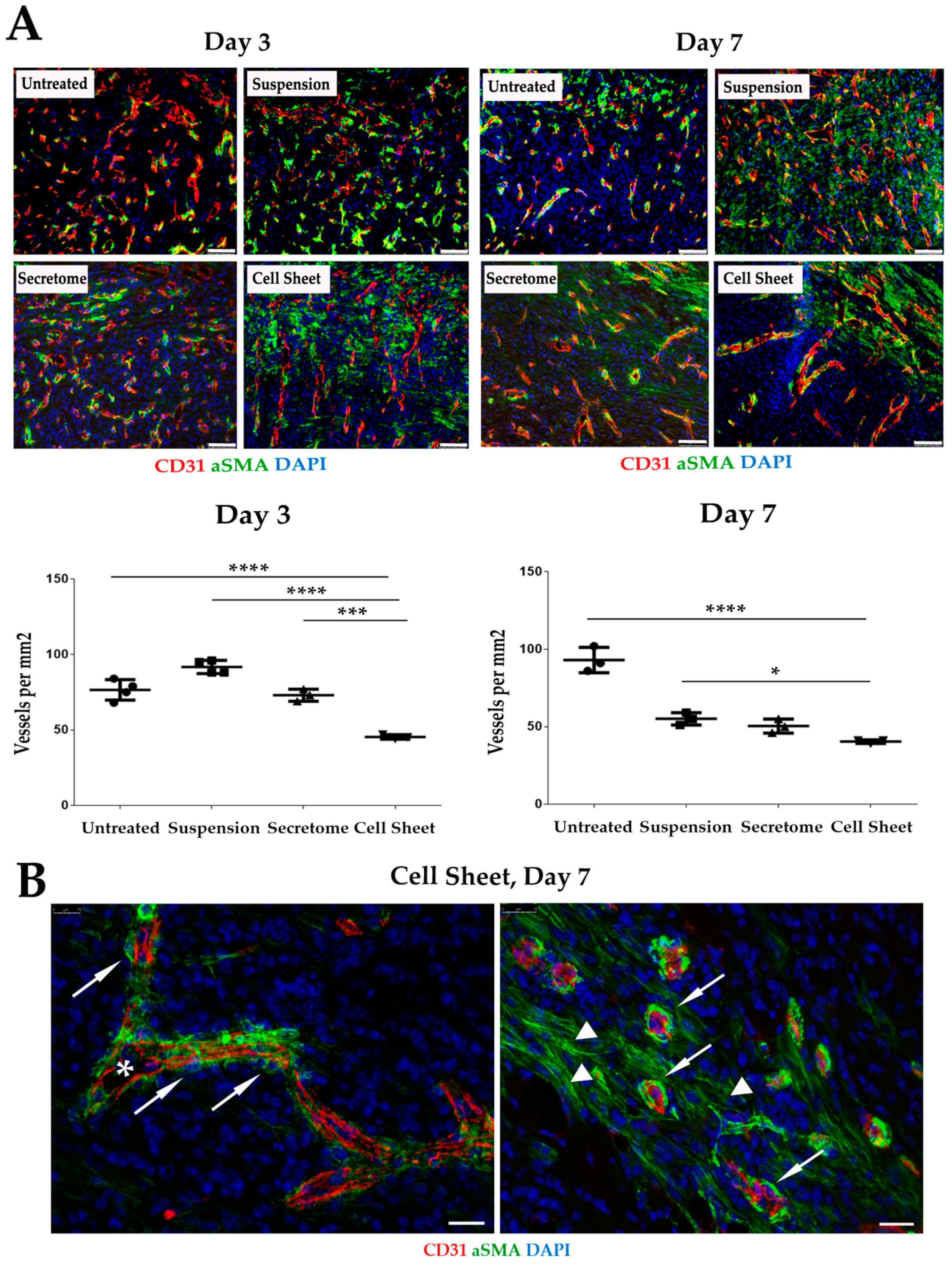
2.5. Vascularization of GT Is Modulated by MSC Secretome and CS
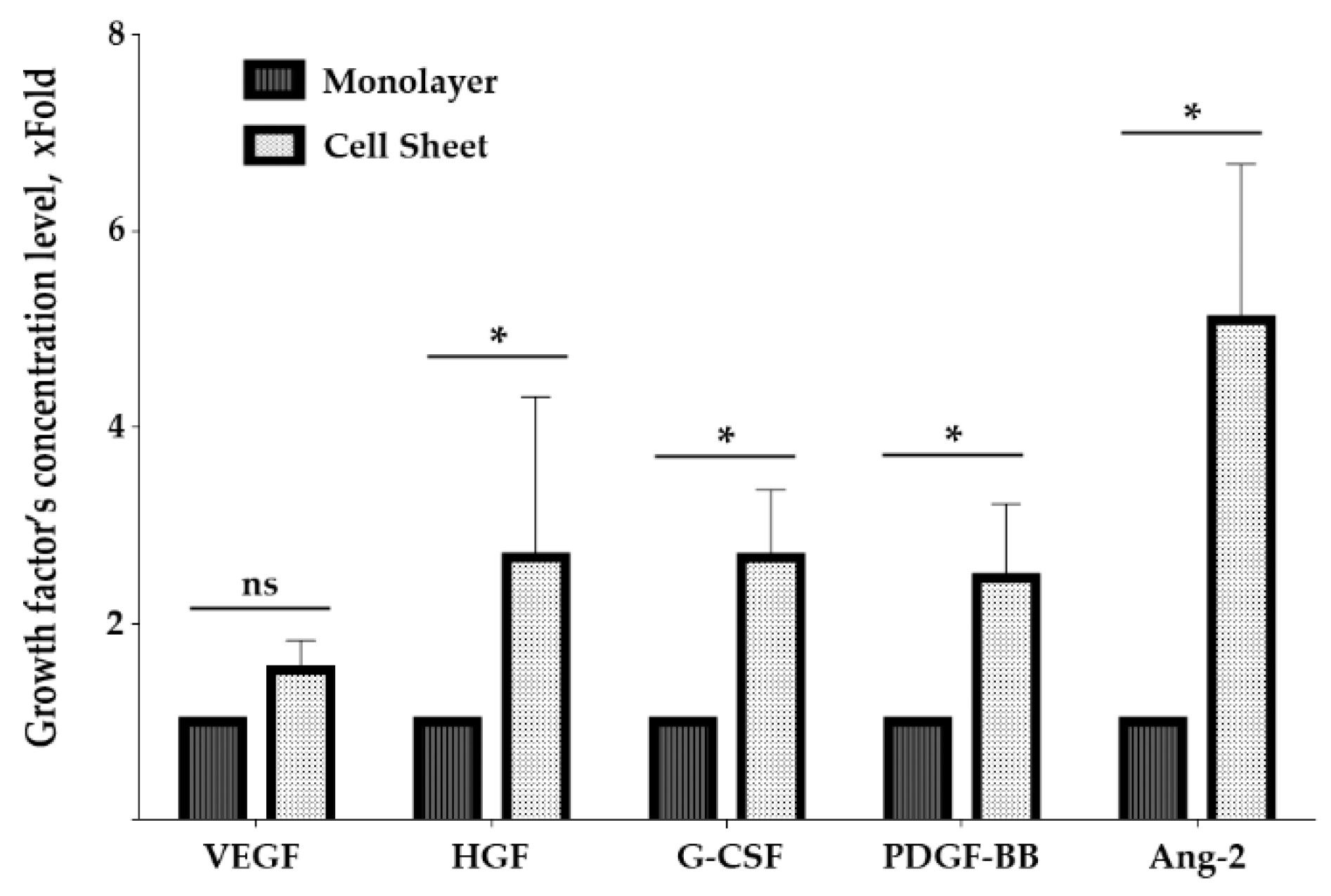
2.6. Assembly of MSC in Cell Sheets Increases Secretion of Growth Factors Involved in Blood Vessel Maturation
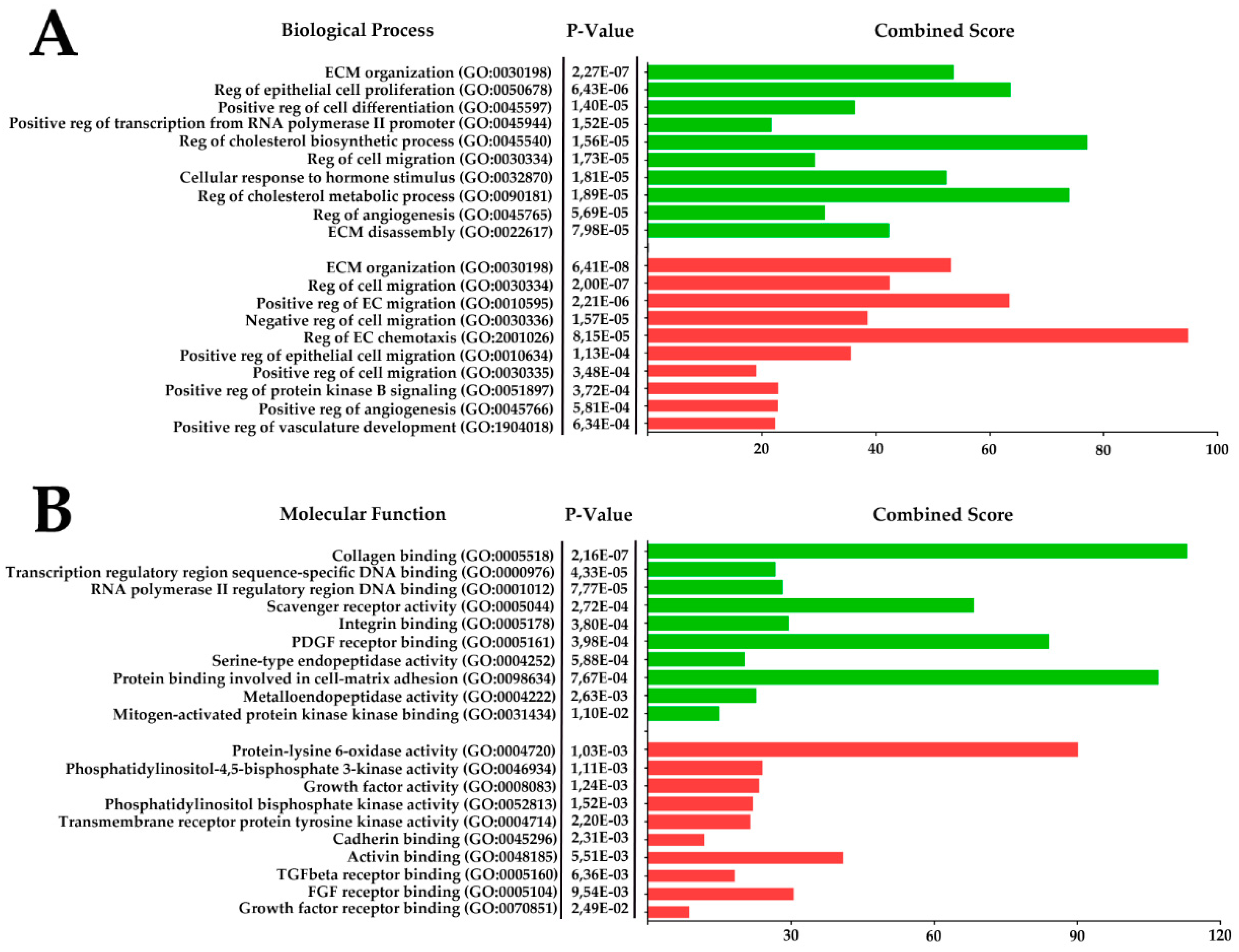
2.7. RNA-Sequencing of Cell Sheets Shows Significant Changes of MSC Transcriptome Profile Compared to Monolayer Culture
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. MSC Secretome Preparation
4.3. Fabrication of Cell Sheets from MSC
4.4. BioPlex Assay of Growth Factors in MSC Monolayer and Cell Sheet Secretomes
4.5. RNA Isolation and Transcriptome RNA-Sequencing Analysis
4.6. Pressure Ulcer Model
4.7. Delivery of MSC Suspension, Secretome or MSC Sheets
- (1)
- “Suspension”—injection suspended MSC to the edges and bottom of skin defect.
- (2)
- “Secretome”—injection of MSC secretome into the edges and bottom of skin defect.
- (3)
- “Cell Sheet”—application of CS to ulcer surface.
- (4)
- “Untreated”—animals without therapy.
4.8. Wound Assessment and Histological Examination
4.9. Histological Evaluaton of Healing Processes: Granulation Tissue Formation and Maturation
4.10. Immunofluorescent Staining and Vessel Density Analysis
4.11. PKH26 Labelling and Subsequent Detection of Transplanted Cells in Histological Preparations
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Ang-1 | Angiopoietin-1 |
| Ang-2 | Angiopoietin-2 |
| ANOVA | Analysis of Variation |
| αSMA | Alpha smooth muscle actin |
| BP | Biological process |
| CS | Cell sheet |
| DEG | Differentially expressed genes |
| DNA | Deoxyribonucleic acid |
| ECM | Extracellular matrix |
| ELISA | Enzyme-linked immunosorbent assay |
| FBS | Fetal bovine serum |
| FGF | Fibroblast growth factor |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| G-CSG | Granulocyte-colony stimulating factor |
| GO | Gene ontology |
| GT | Granulation tissue |
| HGF | Hepatocyte growth factor |
| LOX | Lysyl oxidase |
| MF | Molecular function |
| MSC | Mesenchymal stromal cell |
| NFKB1 | Nuclear factor κB |
| PDGF | Platelet-derived growth factor |
| RELA | REL-associated protein |
| RNA | Ribonucleic acid |
| SP-1 | Specificity protein 1 |
| TF | Transcription factor |
| TGF | Transforming growth factor |
| Tie-2 | Endothelial specific receptor tyrosine kinase |
| VEGF | Vascular endothelial growth factor |
References
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Jackson, C.J. Extracellular Matrix Reorganization During Wound Healing and Its Impact on Abnormal Scarring. Adv. Wound Care 2015, 4, 119–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.E.; Foster, D.S.; Hu, M.S.; Longaker, M.T. Wound healing and fibrosis: Current stem cell therapies. Transfusion 2019, 59, 884–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimiritsky, P.P.; Eremichev, R.Y.; Alexandrushkina, N.A.; Efimenko, A.Y.; Tkachuk, V.A.; Makarevich, P.I. Unveiling Mesenchymal Stromal Cells’ Organizing Function in Regeneration. Int. J. Mol. Sci. 2019, 20, 823. [Google Scholar] [CrossRef] [Green Version]
- Kosaric, N.; Kiwanuka, H.; Gurtner, G.C. Stem cell therapies for wound healing. Expert Opin. Biol. Ther. 2019, 19, 575–585. [Google Scholar] [CrossRef]
- Parekkadan, B.; Milwid, J.M. Mesenchymal stem cells as therapeutics. Annu. Rev. Biomed. Eng. 2010, 12, 87–117. [Google Scholar] [CrossRef] [Green Version]
- Squillaro, T.; Peluso, G.; Galderisi, U. Clinical Trials With Mesenchymal Stem Cells: An Update. Cell Transpl. 2016, 25, 829–848. [Google Scholar] [CrossRef] [Green Version]
- Aleksandrushkina, N.A.; Danilova, N.V.; Grigorieva, O.A.; Mal’kov, P.G.; Popov, V.S.; Efimenko, A.Y.; Makarevich, P.I. Cell Sheets of Mesenchymal Stromal Cells Effectively Stimulate Healing of Deep Soft Tissue Defects. Bull. Exp. Biol. Med. 2019, 167, 159–163. [Google Scholar] [CrossRef]
- Sagaradze, G.; Grigorieva, O.; Nimiritsky, P.; Basalova, N.; Kalinina, N.; Akopyan, Z.; Efimenko, A. Conditioned Medium from Human Mesenchymal Stromal Cells: Towards the Clinical Translation. Int. J. Mol. Sci. 2019, 20, 1656. [Google Scholar] [CrossRef] [Green Version]
- Boldyreva, M.A.; Shevchenko, E.K.; Molokotina, Y.D.; Makarevich, P.I.; Beloglazova, I.B.; Zubkova, E.S.; Dergilev, K.V.; Tsokolaeva, Z.I.; Penkov, D.; Hsu, M.N.; et al. Transplantation of Adipose Stromal Cell Sheet Producing Hepatocyte Growth Factor Induces Pleiotropic Effect in Ischemic Skeletal Muscle. Int. J. Mol. Sci. 2019, 20, 3088. [Google Scholar] [CrossRef] [Green Version]
- Hsu, M.N.; Liao, H.T.; Truong, V.A.; Huang, K.L.; Yu, F.J.; Chen, H.H.; Nguyen, T.K.N.; Makarevich, P.; Parfyonova, Y.; Hu, Y.C. CRISPR-based Activation of Endogenous Neurotrophic Genes in Adipose Stem Cell Sheets to Stimulate Peripheral Nerve Regeneration. Theranostics 2019, 9, 6099–6111. [Google Scholar] [CrossRef] [PubMed]
- Makarevich, P.I.; Boldyreva, M.A.; Gluhanyuk, E.V.; Efimenko, A.Y.; Dergilev, K.V.; Shevchenko, E.K.; Sharonov, G.V.; Gallinger, J.O.; Rodina, P.A.; Sarkisyan, S.S.; et al. Enhanced angiogenesis in ischemic skeletal muscle after transplantation of cell sheets from baculovirus-transduced adipose-derived stromal cells expressing VEGF165. Stem. Cell Res. Ther. 2015, 6, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, M.N.; Liao, H.T.; Li, K.C.; Chen, H.H.; Yen, T.C.; Makarevich, P.; Parfyonova, Y.; Hu, Y.C. Adipose-derived stem cell sheets functionalized by hybrid baculovirus for prolonged GDNF expression and improved nerve regeneration. Biomaterials 2017, 140, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Dergilev, K.; Tsokolaeva, Z.; Makarevich, P.; Beloglazova, I.; Zubkova, E.; Boldyreva, M.; Ratner, E.; Dyikanov, D.; Menshikov, M.; Ovchinnikov, A.; et al. C-Kit Cardiac Progenitor Cell Based Cell Sheet Improves Vascularization and Attenuates Cardiac Remodeling following Myocardial Infarction in Rats. Biomed. Res. Int. 2018, 2018, 3536854. [Google Scholar] [CrossRef]
- Takahashi, H.; Okano, T. Thermally-triggered fabrication of cell sheets for tissue engineering and regenerative medicine. Adv. Drug Deliv. Rev. 2019, 138, 276–292. [Google Scholar] [CrossRef]
- Jun, I.; Ahmad, T.; Bak, S.; Lee, J.Y.; Kim, E.M.; Lee, J.; Lee, Y.B.; Jeong, H.; Jeon, H.; Shin, H. Spatially Assembled Bilayer Cell Sheets of Stem Cells and Endothelial Cells Using Thermosensitive Hydrogels for Therapeutic Angiogenesis. Adv. Healthc. Mater. 2017, 6. [Google Scholar] [CrossRef]
- Cesarz, Z.; Tamama, K. Spheroid Culture of Mesenchymal Stem Cells. Stem. Cells Int. 2016, 2016, 9176357. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.; Lee, H.S.; Lee, G.Y.; Park, G.; Kim, T.M.; Shin, J.; Lee, C.; Oh, I.H. Shift of EMT gradient in 3D spheroid MSCs for activation of mesenchymal niche function. Sci. Rep. 2017, 7, 6859. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.; Venkatachalapathy, S.; Ratna, P.; Wang, Y.; Jokhun, D.S.; Nagarajan, M.; Shivashankar, G.V. Laterally confined growth of cells induces nuclear reprogramming in the absence of exogenous biochemical factors. Proc. Natl. Acad. Sci. USA 2018, 115, E4741–E4750. [Google Scholar] [CrossRef] [Green Version]
- Pennock, R.; Bray, E.; Pryor, P.; James, S.; McKeegan, P.; Sturmey, R.; Genever, P. Human cell dedifferentiation in mesenchymal condensates through controlled autophagy. Sci. Rep. 2015, 5, 13113. [Google Scholar] [CrossRef] [Green Version]
- Sagaradze, G.D.; Nimiritsky, P.P.; Akopyan, Z.A.; Makarevich, P.I.; Efimenko, A.Y. “Cell-Free Therapeutics” from Components Secreted by Mesenchymal Stromal Cells as a Novel Class of Biopharmaceuticals. IntechOpen 2018. [Google Scholar] [CrossRef] [Green Version]
- Makarevich, P.I.; Parfyonova, Y.V. Therapeutic Angiogenesis: Foundations and Practical Application. IntechOpen 2017. [Google Scholar] [CrossRef] [Green Version]
- Hsu, I.; Parkinson, L.G.; Shen, Y.; Toro, A.; Brown, T.; Zhao, H.; Bleackley, R.C.; Granville, D.J. Serpina3n accelerates tissue repair in a diabetic mouse model of delayed wound healing. Cell Death Dis. 2014, 5, e1458. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, R.; Luo, G.; Lei, Q.; Shu, Q.; Yao, Z.; Li, H.; Zhou, J.; Tan, J.; Yang, S.; et al. Biomimetic fibroblast-loaded artificial dermis with "sandwich" structure and designed gradient pore sizes promotes wound healing by favoring granulation tissue formation and wound re-epithelialization. Acta Biomater. 2016, 30, 246–257. [Google Scholar] [CrossRef]
- Chen, A.; He, H.; Ma, G.; Li, Y.; Jiang, S.; Xuan, X.; Song, Y.; Zhang, C.; Xiao, J.; Xu, Y.; et al. Biodegradable copolypeptide hydrogel prodrug accelerates dermal wound regeneration by enhanced angiogenesis and epithelialization. RSC Adv. 2018, 8, 10620–10626. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.E.; Wilgus, T.A. Vascular Endothelial Growth Factor and Angiogenesis in the Regulation of Cutaneous Wound Repair. Adv. Wound Care 2014, 3, 647–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinnell, F. Fibroblasts, myofibroblasts, and wound contraction. J. Cell Biol. 1994, 124, 401–404. [Google Scholar] [CrossRef]
- Lee, W.Y.; Chang, Y.H.; Yeh, Y.C.; Chen, C.H.; Lin, K.M.; Huang, C.C.; Chang, Y.; Sung, H.W. The use of injectable spherically symmetric cell aggregates self-assembled in a thermo-responsive hydrogel for enhanced cell transplantation. Biomaterials 2009, 30, 5505–5513. [Google Scholar] [CrossRef]
- Yu, J.; Wang, M.Y.; Tai, H.C.; Cheng, N.C. Cell sheet composed of adipose-derived stem cells demonstrates enhanced skin wound healing with reduced scar formation. Acta Biomater. 2018, 77, 191–200. [Google Scholar] [CrossRef]
- Tonnesen, M.G.; Feng, X.; Clark, R.A. Angiogenesis in wound healing. J. Investig. Dermatol. Symp. Proc. 2000, 5, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Müller, B.; Lang, S.; Dominietto, M.; Rudin, M.; Schulz, G.; Deyhle, H.; Germann, M.; Pfeiffer, F.; David, C.; Weitkamp, T. High-Resolution Tomographic Imaging of Microvessels; SPIE: Bellingham, WA, USA, 2008; Volume 7078. [Google Scholar]
- Stratman, A.N.; Malotte, K.M.; Mahan, R.D.; Davis, M.J.; Davis, G.E. Pericyte recruitment during vasculogenic tube assembly stimulates endothelial basement membrane matrix formation. Blood 2009, 114, 5091–5101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellstrom, M.; Gerhardt, H.; Kalen, M.; Li, X.; Eriksson, U.; Wolburg, H.; Betsholtz, C. Lack of pericytes leads to endothelial hyperplasia and abnormal vascular morphogenesis. J. Cell Biol. 2001, 153, 543–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Carlo, S.E.; Peduto, L. The perivascular origin of pathological fibroblasts. J. Clin. Investig. 2018, 128, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, S.N.; Iredale, J.P.; Henderson, N.C. Origins of fibrosis: Pericytes take centre stage. F1000Prime Rep. 2013, 5, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiPietro, L.A. Angiogenesis and scar formation in healing wounds. Curr. Opin. Rheumatol. 2013, 25, 87–91. [Google Scholar] [CrossRef]
- Thurston, G.; Daly, C. The complex role of angiopoietin-2 in the angiopoietin-tie signaling pathway. Cold Spring Harb. Perspect. Med. 2012, 2, a006550. [Google Scholar] [CrossRef] [Green Version]
- Brudno, Y.; Ennett-Shepard, A.B.; Chen, R.R.; Aizenberg, M.; Mooney, D.J. Enhancing microvascular formation and vessel maturation through temporal control over multiple pro-angiogenic and pro-maturation factors. Biomaterials 2013, 34, 9201–9209. [Google Scholar] [CrossRef] [Green Version]
- Lobov, I.B.; Brooks, P.C.; Lang, R.A. Angiopoietin-2 displays VEGF-dependent modulation of capillary structure and endothelial cell survival in vivo. Proc. Natl. Acad. Sci. USA 2002, 99, 11205–11210. [Google Scholar] [CrossRef] [Green Version]
- Teichert, M.; Milde, L.; Holm, A.; Stanicek, L.; Gengenbacher, N.; Savant, S.; Ruckdeschel, T.; Hasanov, Z.; Srivastava, K.; Hu, J.; et al. Pericyte-expressed Tie2 controls angiogenesis and vessel maturation. Nat. Commun. 2017, 8, 16106. [Google Scholar] [CrossRef]
- Logsdon, E.A.; Finley, S.D.; Popel, A.S.; Mac Gabhann, F. A systems biology view of blood vessel growth and remodelling. J. Cell Mol. Med. 2014, 18, 1491–1508. [Google Scholar] [CrossRef]
- Shi, Y.N.; Zhu, N.; Liu, C.; Wu, H.T.; Gui, Y.; Liao, D.F.; Qin, L. Wnt5a and its signaling pathway in angiogenesis. Clin. Chim. Acta 2017, 471, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Rodriguez-Sinovas, A.; Martinez-Gonzalez, J. Lysyl oxidase as a potential therapeutic target. Drug News Perspect. 2008, 21, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.A.; Chen, J.C.; Cerise, J.E.; Jahoda, C.A.; Christiano, A.M. Microenvironmental reprogramming by three-dimensional culture enables dermal papilla cells to induce de novo human hair-follicle growth. Proc. Natl. Acad. Sci. USA 2013, 110, 19679–19688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadler, I.; Zhang, R.Y.; Oskoui, P.; Whittaker, M.S.; Lanzafame, R.J. Development of a simple, noninvasive, clinically relevant model of pressure ulcers in the mouse. J. Investig. Surg. 2004, 17, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yu, Q.; Xu, C.-B. A convenient method for quantifying collagen fibers in atherosclerotic lesions by ImageJ software. Int. J. Clin. Exp. Med. 2017, 10, 14904–14910. [Google Scholar]
- Gurevich, D.B.; Severn, C.E.; Twomey, C.; Greenhough, A.; Cash, J.; Toye, A.M.; Mellor, H.; Martin, P. Live imaging of wound angiogenesis reveals macrophage orchestrated vessel sprouting and regression. EMBO J. 2018, 37. [Google Scholar] [CrossRef]
- Lamas, J.R.; Fernandez-Gutierrez, B.; Mucientes, A.; Marco, F.; Lopiz, Y.; Jover, J.A.; Abasolo, L.; Rodriguez-Rodriguez, L. RNA sequencing of mesenchymal stem cells reveals a blocking of differentiation and immunomodulatory activities under inflammatory conditions in rheumatoid arthritis patients. Arthritis Res. Ther. 2019, 21, 112. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | TF | Overlapped Genes N | List of Overlapped Genes |
|---|---|---|---|
| 1 | SP1 | 28 | C4A, AGTR1, EDNRB, AGT, PPL, MMP2, TNC, EGR1, LSP1, CCND2, TNFSF10, MME, FBLN1, CYP27A1, SOX9, COMP, SLC39A8, COL18A1, C4B, SREBF1, HGF, TCN2, ISG20, APOE, SOD2, CYP19A1, PTN, CHI3L1 |
| 2 | NFKB1 | 22 | EGR1, BGN, CYP19A1, PTGFR, TNC, SOD2, ADORA1, CXCL12, TNFSF13B, MMP2, CD74, FGF7, IRF4, AGT, SLC25A27, PLA2G2A, CCND2, BCL2L11, IRF7, VCAM1, A2M, TNFSF10 |
| 3 | RELA | 20 | IRF7, EGR1, FGF7, ADORA1, CXCL12, MMP2, SLC25A27, SOD2, VCAM1, BCL2L11, TNFSF10, PLA2G2A, TNC, CD74, CYP19A1, CCND2, PTGFR, AGT, BGN, IRF4 |
| 4 | FOXO3 | 8 | BCL6, TXNIP, TNFSF10, BCL2L11, CCND2, VEGFA, CDKN2B, VEGFB |
| 5 | USF1 | 8 | FMO3, TCN2, AGT, CYP19A1, SLC1A3,LIPC, CTSD, ISG20 |
| 6 | SREBF1 | 5 | LRP1, ACACB, LDLR, FASN, CIC |
| 7 | STAT3 | 10 | CCND2, CFB, PROS1, HGF, MMP2, CYP19A1, A2M, CHI3L1, DIRAS3, BCL6 |
| 8 | HIF1A | 12 | TGFB3, TLR6, VEGFA, ARNT, TIMP2, SOCS1, VEGFB, MMP2, CXCL12, EDNRB, ACE, AGTR1 |
| 9 | FOXO1 | 4 | TNFSF10, EGR1, ANGPT2, TXNIP |
| 10 | JUN | 10 | DCN, VCAM1, CYP19A1, MMP2, SOD2, PTN, TNC, MGP, FGF7, LBP |
| 11 | DNMT1 | 4 | ESR1, IL32, CDKN2B, VEGFA |
| 12 | SP3 | 8 | FBLN1, HGF, SLC1A3, ACE, TCN2, MMP2, AGTR1, CYP27A1 |
| 13 | CREB1 | 7 | NR4A3, AQP3, BCL2L11, BDKRB2, SOX9, MMP2, CYP19A1 |
| 14 | MYC | 7 | VEGFA, JUNB, HLA-B, TFAP4, SHMT1, MST1, BCL2 |
| 15 | ESR1 | 6 | ESR1, BCL2, VEGFA, ZEB1, CEBPB, JUNB |
| 16 | ETS2 | 4 | EGR1, ANGPT2, MMP2, TNC |
| 17 | WT1 | 5 | WTAP, BCL2, VEGFA, VEGFB, JUNB |
| 18 | ATF4 | 4 | IRF7, HRK, DDIT4, APOE |
| 19 | PPARA | 4 | TXNIP, SOD2, G0S2, CD36 |
| 20 | SPI1 | 5 | JCHAIN, BCL6, MME, CTSS, CTSK |
| 21 | MYCN | 4 | EFNB3, CTSD, MXI1, CLU |
| 22 | USF2 | 4 | TCN2, CTSD, CYP19A1, LIPC |
| 23 | HDAC1 | 5 | HLA-DRA, EGR1, CCND2, TXNIP, SFRP1 |
| 24 | STAT1 | 4 | UPP1, XAF1, STAT2, MUC1 |
| 25 | BRCA1 | 4 | ESR1, IRF9, VEGFA, DDIT3 |
| 26 | TP53 | 8 | BDKRB2, GPNMB, PDGFRB, AQP3, EGR1, PMAIP1, CTSD, MMP2 |
| 27 | ETS1 | 5 | DUSP6, ANGPT2, EGR1, TMEM158, TNC |
| 28 | YY1 | 5 | SAP30, VWF, LSS, VEGFB, LDLR |
| 29 | BRCA1 | 4 | EGR1, CTSD, IRF7, CYP19A1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alexandrushkina, N.; Nimiritsky, P.; Eremichev, R.; Popov, V.; Arbatskiy, M.; Danilova, N.; Malkov, P.; Akopyan, Z.; Tkachuk, V.; Makarevich, P. Cell Sheets from Adipose Tissue MSC Induce Healing of Pressure Ulcer and Prevent Fibrosis via Trigger Effects on Granulation Tissue Growth and Vascularization. Int. J. Mol. Sci. 2020, 21, 5567. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155567
Alexandrushkina N, Nimiritsky P, Eremichev R, Popov V, Arbatskiy M, Danilova N, Malkov P, Akopyan Z, Tkachuk V, Makarevich P. Cell Sheets from Adipose Tissue MSC Induce Healing of Pressure Ulcer and Prevent Fibrosis via Trigger Effects on Granulation Tissue Growth and Vascularization. International Journal of Molecular Sciences. 2020; 21(15):5567. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155567
Chicago/Turabian StyleAlexandrushkina, Natalya, Peter Nimiritsky, Roman Eremichev, Vladimir Popov, Mikhail Arbatskiy, Natalia Danilova, Pavel Malkov, Zhanna Akopyan, Vsevolod Tkachuk, and Pavel Makarevich. 2020. "Cell Sheets from Adipose Tissue MSC Induce Healing of Pressure Ulcer and Prevent Fibrosis via Trigger Effects on Granulation Tissue Growth and Vascularization" International Journal of Molecular Sciences 21, no. 15: 5567. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155567