Diversity and Antimicrobial Activity of Endophytic Fungi Isolated from Chloranthus japonicus Sieb in Qinling Mountains, China


Abstract
:1. Introduction
2. Results
2.1. Isolation, Sequencing, Identification, and Diversity Analyses of the Endophytic Fungi from C. japonicus
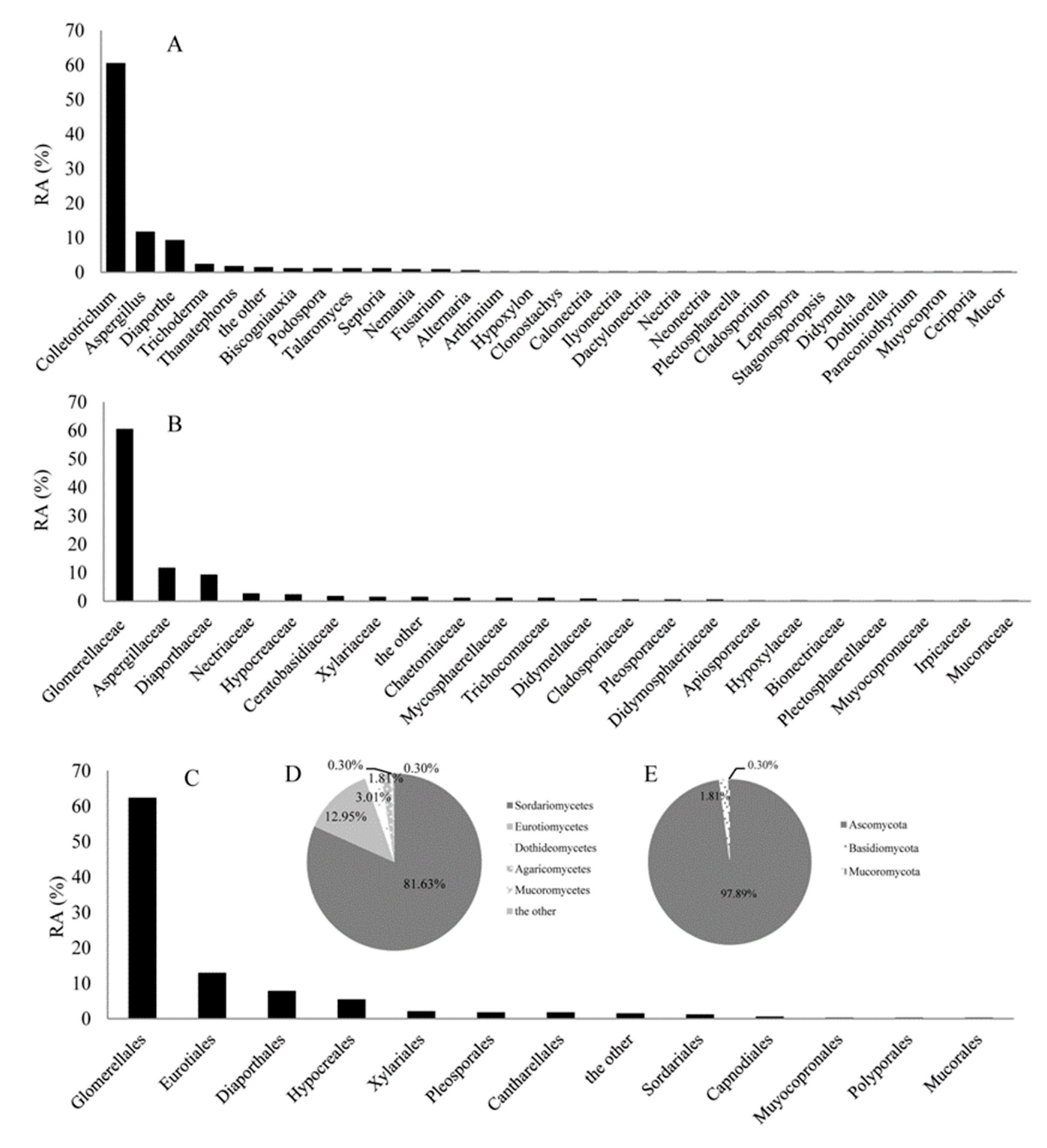
2.2. The Relative Abundance (RA) Analyses of Endophytic fungi from C. japonicus
2.3. Antimicrobial Activity Screening of the Ethyl Acetate Extracts from Endophytic Fungal Culture Filtrates
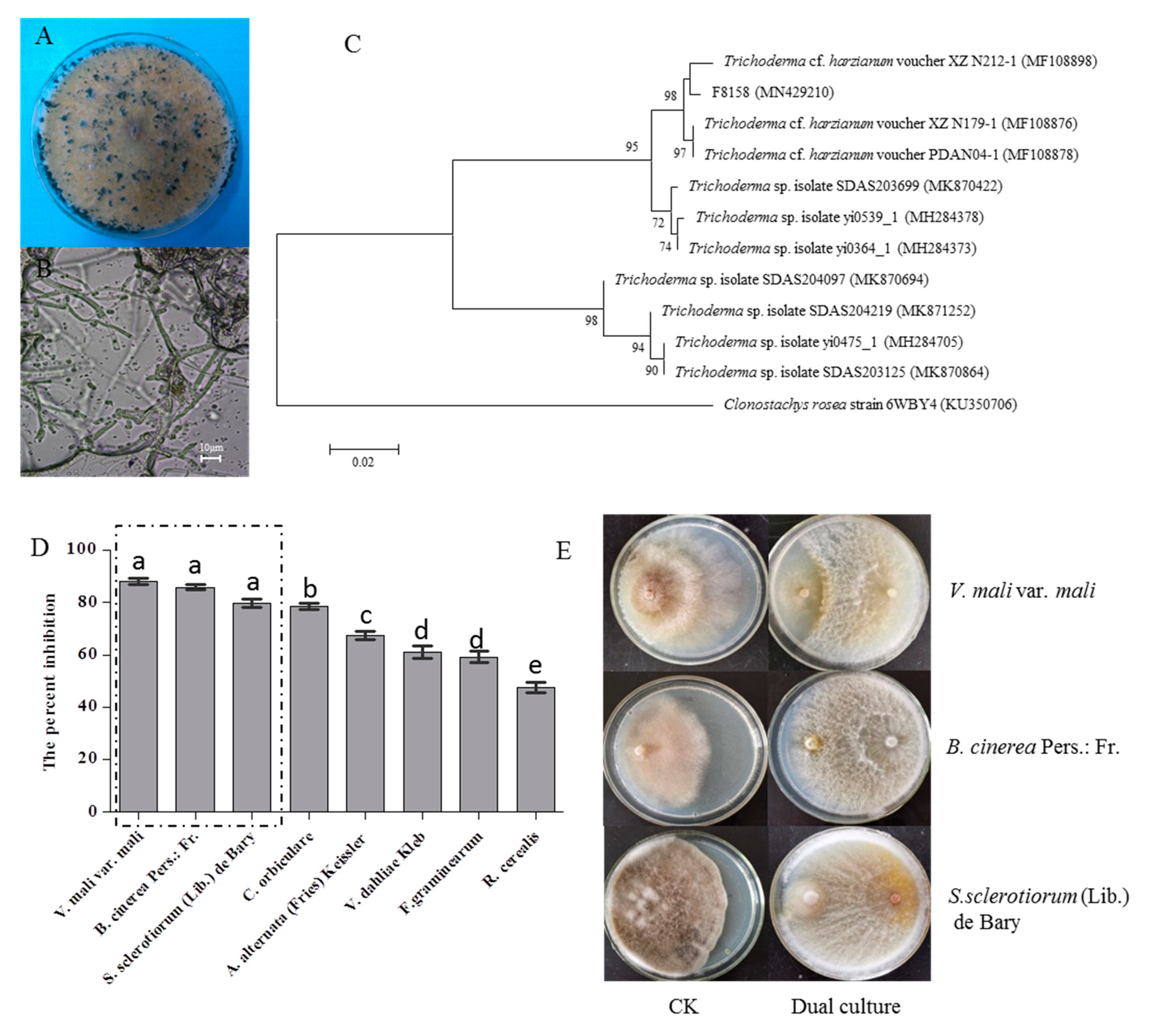
2.4. Identification of the Strain F8158 and Analysis of Its Antagonistic Effect for Different Pathogens
3. Discussion
4. Materials and Methods
4.1. Source of Plant Samples
4.2. Fungal Isolation and Cultivation
4.3. Molecular Identification and Phylogenetic Analyses
4.4. Crude Extract Preparation of Fungal Fermentation Broth
4.5. Antimicrobial Activity
4.6. Dual Culture Assay to Detect the Antagonistic Potential of Endophytic Fungi against Different Pathogenic Fungi
4.7. Diversity Analyses of the Endophytic Fungi
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BLAST | Basic Local Alignment Search Tool |
| IF | Isolation frequency |
| ITS | Internal transcribed spacer |
| PCR | Polymerase chain reaction |
| RA | Relative abundance |
| SSA | Similarity threshold |
References
- Ma, T.; Hu, Y.; Russo, I.R.M.; Nie, Y.; Yang, T.; Xiong, L.; Ma, S.; Meng, T.; Han, H.; Zhang, X.; et al. Walking in a heterogeneous landscape: Dispersal, gene flow and conservation implications for the giant panda in the Qinling Mountains. Evol. Appl. 2018, 11, 1859–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Zheng, X.; Cheng, F.; Zhu, X.; Hou, L.; Li, J.; Zhang, S. Fungal community structure of fallen pine and oak wood at different stages of ecomposition in the Qinling Mountains. China. Sci. Rep. 2017, 7, 13866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Duan, R.; Wang, S.; Wang, Z.; Fan, W. Species presence frequency and diversity in different patch types along an altitudinal gradient: Larix chinensis Beissn in Qinling Mountains (China). PeerJ 2016, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.H.; Chu, H.L.; Dou, Q.; Xie, Q.Z.; Tang, M.; Sung, C.K.; Wang, C.Y. Phosphorus and nitrogen drive the seasonal dynamics of bacterial communities in Pinus forest fhizospheric soil of the Qinling Mountains. Front. Microbiol. 2018, 9, 1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, P.L.; Cao, Y.L.; Yan, H.; Pan, L.L.; Liu, S.C.; Gong, N.B.; Lu, Y.; Chen, C.X.; Zhong, H.M.; Guo, Y.; et al. Lindenane Disesquiterpenoids with Anti-HIV-1 Activity from Chloranthus japonicus. J. Nat. Prod. 2011, 74, 1408–1413. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Qin, X.J.; Li, X.H.; Yu, Q.; Ni, W.; He, L.; Liu, H.Y. Japonicones A-C: Three lindenane sesquiterpenoid dimers with a 12-membered ring core from Chloranthus japonicus. Tetrahedron Lett. 2019, 60, 713–717. [Google Scholar] [CrossRef]
- Kim, S.Y.; Nagashima, H.; Tanaka, N.; Kashiwada, Y.; Kobayashi, J.; Kojoma, M. Hitorins A and B, Hexacyclic C25 Terpenoids from Chloranthus japonicus. Org. Lett. 2016, 18, 5420–5423. [Google Scholar] [CrossRef]
- Du, W.; Yao, Z.; Li, J.; Sun, C.; Xia, J.; Wang, B.; Shi, D.; Ren, L. Diversity and antimicrobial activity of endophytic fungi isolated from Securinega suffruticosa in the Yellow River Delta. PLoS ONE 2020, 15, e0229589. [Google Scholar] [CrossRef]
- Khan, A.L.; Hussain, J.; Al-Harrasi, A.; Al-Rawahi, A.; Lee, I.J. Endophytic fungi: Resource for gibberellins and crop abiotic stress resistance. Crit. Rev. Biotechnol. 2013, 35, 62–74. [Google Scholar] [CrossRef]
- Gupta, S.; Chaturvedi, P.; Kulkarni, M.G.; Van Staden, J. A critical review on exploiting the pharmaceutical potential of plant endophytic fungi. Biotechnol. Adv. 2020, 39, 107462. [Google Scholar] [CrossRef]
- Arora, P.; Wani, Z.A.; Ahmad, T.; Sultan, P.; Gupta, S.; Riyaz-Ul-Hassan, S. Community structure, spatial distribution, diversity and functional characterization of culturable endophytic fungi associated with Glycyrrhiza glabra L. Fungal Boil. 2019, 123, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Kasaei, A.; Dehkordi, M.M.; Mahjoubi, F.; Saffar, B. Isolation of Taxol-Producing Endophytic Fungi from Iranian Yew Through Novel Molecular Approach and Their Effects on Human Breast Cancer Cell Line. Curr. Microbiol. 2017, 62, 164–709. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.; Vasundhara, M. Isolation of taxol producing endophytic fungus Alternaria brassicicola from non-Taxus medicinal plant Terminalia arjuna. World J. Microbiol. Biotechnol. 2019, 35, 74. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.M.; Zhou, Y.Q.; Zhou, X.L.; Xia, X.H.; Wei, Y.; He, L.L.; Tang, H.Z.; Yu, L.Y. Diversity and bioactive potential of culturable fungal endophytes of Dysosma versipellis; a rare medicinal plant endemic to China. Sci. Rep. 2018, 8, 5929. [Google Scholar] [CrossRef]
- Payne, D.J.; Gwynn, M.N.; Holmes, D.J.; Pompliano, D.L. Drugs for bad bugs: Confronting the challenges of antibacterial discovery. Nat. Rev. Drug Discov. 2006, 6, 29–40. [Google Scholar] [CrossRef]
- Sunil, K.D.; Manish, K.G.; Ved, P.; Sanjai, S. Endophytic fungi: A source of potential antifungal compounds. J. Fungi 2018, 4, 77–119. [Google Scholar]
- Rinu, K.; Sati, P.; Pandey, A. Trichoderma gamsii(NFCCI 2177): A newly isolated endophytic, psychrotolerant, plant growth promoting, and antagonistic fungal strain. J. Basic Microbiol. 2013, 54, 408–417. [Google Scholar] [CrossRef]
- Redda, E.T.; Ma, J.; Mei, J.; Li, M.; Wu, B.; Jiang, X. Antagonistic Potential of Different Isolates of Trichoderma against Fusarium oxysporum, Rhizoctonia solani, and Botrytis cinerea. Eur. J. Exp. Boil. 2018, 8. [Google Scholar] [CrossRef]
- Toghueo, R.M.K.; Boyom, F.F. Endophytic Fungi from Terminalia Species: A Comprehensive Review. J. Fungi 2019, 5, 43. [Google Scholar] [CrossRef]
- Li, P.; Wu, Z.; Liu, T.; Wang, Y. Biodiversity, Phylogeny, and Antifungal Functions of Endophytic Fungi Associated with Zanthoxylum bungeanum. Int. J. Mol. Sci. 2016, 17, 1541. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.K.; Tuo, L.; Huang, D.L.; Osterman, I.A.; Tyurin, A.P.; Liu, S.W.; Lukyanov, D.A.; Sergiev, P.V.; Dontsova, O.A.; Korshun, V.A.; et al. Diversity, Novelty, and Antimicrobial Activity of Endophytic Actinobacteria From Mangrove Plants in Beilun Estuary National Nature Reserve of Guangxi, China. Front. Microbiol. 2018, 9, 868–879. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, S.J.; Li, J.J.; Liang, Z.; Zhao, C.Q. Novel Natural Products from Extremophilic Fungi. Mar. Drugs. 2018, 16, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, A.; Rodrigues, M.J.; Pereira, C.G.; Oliveira, M.; Barreira, L.; Varela, J.; Trampetti, F.; Custódio, L. Natural products from extreme marine environments: Searching for potential industrial uses within extremophile plants. Ind. Crop. Prod. 2016, 94, 299–307. [Google Scholar] [CrossRef]
- Sayed, A.M.; Hassan, M.H.; Alhadrami, H.A.; Hassan, H.M.; Goodfellow, M.; Rateb, M.E. Extreme environments: Microbiology leading to specialized metabolites. J. Appl. Microbiol. 2019, 128, 630–657. [Google Scholar] [CrossRef] [Green Version]
- El-Hawary, S.S.; Moawad, A.S.; Bahr, H.S.; Abdelmohsen, U.R.; Mohammed, R. Natural product diversity from the endophytic fungi of the genus Aspergillus. RSC Adv. 2020, 10, 22058–22079. [Google Scholar] [CrossRef]
- Nicoletti, R.; Vinale, F. Bioactive Compounds from Marine-Derived Aspergillus, Penicillium, Talaromyces and Trichoderma Species. Mar. Drugs 2018, 16, 408. [Google Scholar] [CrossRef] [Green Version]
- Chepkirui, C.; Stadler, M. The genus Diaporthe: A rich source of diverse and bioactive metabolites. Mycol. Prog. 2017, 16, 477–494. [Google Scholar] [CrossRef]
- Liu, Z.; Zhao, J.Y.; Sun, S.-F.; Li, Y.; Qu, J.; Liu, H.T.; Liu, Y. Sesquiterpenes from an Endophytic Aspergillus flavus. J. Nat. Prod. 2019, 82, 1063–1071. [Google Scholar] [CrossRef]
- Guo, L.; Niu, S.; Chen, S.; Liu, L. Diaporone A, a new antibacterial secondary metabolite from the plant endophytic fungus Diaporthe sp. J. Antibiot. 2019, 73, 116–119. [Google Scholar] [CrossRef]
- Song, Y.P.; Miao, F.P.; Liu, X.H.; Yin, X.; Ji, N. Cyclonerane Derivatives from the Algicolous Endophytic Fungus Trichoderma asperellum A-YMD-9-2. Mar. Drugs 2019, 17, 252. [Google Scholar] [CrossRef] [Green Version]
- da Silva, J.A.T.; de Medeiros, E.V.; da Silva, J.M.; Tenório, D.d.A.; Moreira, K.A.; da Silva Nascimento, T.C.E.; Souza-Motta, C. Antagonistic activity of Trichoderma spp. against Scytalidium lignicola CMM 1098 and antioxidant enzymatic activity in cassava. Phytoparasitica 2017, 45, 219–225. [Google Scholar] [CrossRef]
- Wonglom, P.; Daengsuwan, W.; Ito, S.I.; Sunpapao, A. Biological control of Sclerotium fruit rot of snake fruit and stem rot of lettuce by Trichoderma sp. T76-12/2 and the mechanisms involved. Physiol. Mol. Plant Pathol. 2019, 107, 1–7. [Google Scholar] [CrossRef]
- Yuan, S.; Li, M.; Fang, Z.; Liu, Y.; Shi, W.; Pan, B.; Wu, K.; Shi, J.; Shen, B.; Shen, Q. Biological control of tobacco bacterial wilt using Trichoderma harzianum amended bioorganic fertilizer and the arbuscular mycorrhizal fungi Glomus mosseae. Boil. Control. 2016, 92, 164–171. [Google Scholar] [CrossRef]
- Tan, X.M.; Wang, C.L.; Chen, X.M.; Zhou, Y.Q.; Wang, Y.Q.; Luo, A.X.; Liu, Z.H.; Guo, S.X. In vitro seed germination and seedling growth of an endangered epiphytic orchid, Dendrobium officinale, endemic to China using mycorrhizal fungi (Tulasnella sp.). Sci. Hortic. 2014, 165, 62–68. [Google Scholar] [CrossRef]
- Qin, S.; Li, J.; Chen, H.H.; Zhao, G.Z.; Zhu, W.Y.; Jiang, C.L.; Xu, L.H.; Li, W.J. Isolation, Diversity, and Antimicrobial Activity of Rare Actinobacteria from Medicinal Plants of Tropical Rain Forests in Xishuangbanna, China. Appl. Environ. Microbiol. 2009, 75, 6176–6186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.M.; Chen, X.M.; Wang, C.L.; Jin, X.H.; Cui, J.L.; Chen, J.; Guo, S.X.; Zhao, L.F. Isolation and Identification of Endophytic Fungi in Roots of Nine Holcoglossum Plants (Orchidaceae) Collected from Yunnan, Guangxi, and Hainan Provinces of China. Curr. Microbiol. 2011, 64, 140–147. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: Berkeley, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Wei, W.; Zhou, Y.; Chen, F.; Yan, X.; Lai, Y.; Wei, C.; Chen, X.; Xu, J.; Wang, X. Isolation, Diversity, and Antimicrobial and Immunomodulatory Activities of Endophytic Actinobacteria From Tea Cultivars Zijuan and Yunkang-10 (Camellia sinensis var. assamica). Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Xing, Y.-M.; Chen, J.; Cui, J.L.; Chen, X.M.; Guo, S.X. Antimicrobial Activity and Biodiversity of Endophytic Fungi in Dendrobium devonianum and Dendrobium thyrsiflorum from Vietman. Curr. Microbiol. 2010, 62, 1218–1224. [Google Scholar] [CrossRef]
- Wang, S.S.; Liu, J.; Sun, J.; Sun, Y.-F.; Liu, J.N.; Jia, N.; Fan, B.; Dai, X.-F. Diversity of culture-independent bacteria and antimicrobial activity of culturable endophytic bacteria isolated from different Dendrobium stems. Sci. Rep. 2019, 9, 10389. [Google Scholar] [CrossRef] [PubMed]
- Fedor, P.J.; Spellerberg, I.F. Shannon-Wiener Index. Reference Module. Earth Syst. Environ. Sci. 2013, 1, 1–4. [Google Scholar]



{kind=link}
{kind=link}
| No. | Closest Species | Isolates Numbers | Identity (%) | N | IF |
|---|---|---|---|---|---|
| 1 | Arthrinium hydei CBS 114.990 (KF144890) | F8164 | 95.42 | 1 | 0.30 |
| 2 | Aspergillus flavipes AfH14F02 (MK952227) | F8142, F8204, F8225 | 98.73 | 3 | 0.90 |
| 3 | Aspergillus terreus MD32_7 (JQ697508) | F8121, F8159, F8172, F8191 | 99.46 | 20 | 6.02 |
| 4 | Aspergillus pseudoglaucus ALE-85 (MF380824) | F8123 | 99.24 | 1 | 0.30 |
| 5 | Aspergillus niger strain H1 (KJ778683) | F8143, F8150, F8186 | 99.30 | 6 | 1.81 |
| 6 | Aspergillus oryzae isolate 118 (MH345958) | F8160, F8161, F8224 | 99.82 | 4 | 1.20 |
| 7 | Aspergillus sp. isolate SW511 (MH509427) | F8190, F8243, F8238 | 99.47 | 3 | 0.90 |
| 8 | Aspergillus tubingensis BSZ-6(2) (KJ190960) | F8208 | 99.81 | 1 | 0.30 |
| 9 | Aspergillus flavus strain S2599 (MG575474) | F8111 | 99.64 | 1 | 0.30 |
| 10 | Clonostachys rosea QLF2 (FJ025204) | F8195 | 99.26 | 1 | 0.30 |
| 11 | Thanatephorus cucumeris AG-7 (AY154305) | F8154, F8157 | 98.71 | 6 | 1.80 |
| 12 | Podospora nannopodalis CBS 113.680 (MH862937) | F8109 | 99.02 | 1 | 0.30 |
| 13 | Podospora setosa CBS 613.84 (MH861787) | F8117, F8156 | 99.21 | 3 | 0.90 |
| 14 | Cladosporium tenuissimum 7P6 (KY400093) | F8144 | 99.81 | 1 | 0.30 |
| 15 | Diaporthe eres HUTA286 (KU377286) | F8107, F8205 | 99.26 | 7 | 2.11 |
| 16 | Diaporthe longicolla ALE-196 (MF380785) | F8124 | 97.95 | 1 | 0.30 |
| 17 | Diaporthe nobilis ZZDN1 (MG736062) | F8128, F8196, F8207 | 99.28 | 5 | 1.51 |
| 18 | Diaporthe gulyae SJC3-2 (KX077244) | F8116, F8179, F8192 | 99.63 | 11 | 3.31 |
| 19 | Diaporthe longicolla ALE-196 (MF380785) | F8124 | 97.95 | 1 | 0.30 |
| 20 | Diaporthe vaccinii Lan1 (KC488258) | F8137, F8180, F8206 | 99.11 | 3 | 0.90 |
| 21 | Diaporthe citrichinensis ZJUD034B (KJ210539) | F8193, F8194, F8237 | 97.32 | 3 | 0.90 |
| 22 | Stagonosporopsis cucurbitacearum D49 (MF401570) | F8151 | 99.06 | 1 | 0.30 |
| 23 | Didymella bellidis YS24 (MN443603) | F8165 | 100.00 | 1 | 0.30 |
| 24 | Dothiorella gregaria hz-S8 (FJ517548) | F8178 | 100.00 | 1 | 0.30 |
| 25 | Paraconiothyrium brasiliense 1-53 (JF502455) | F8181 | 99.62 | 1 | 0.30 |
| 26 | Colletotrichum boninense JL7 (KM513575) | F8108 | 99.82 | 1 | 0.30 |
| 27 | Colletotrichum sp. SL10 (KP689235) | F8110, F8135, F8129, F8170, F8176, F8201, F8210, F8213, F8235, F8115, F8120 | 99.10 | 31 | 9.34 |
| 28 | Colletotrichum acutatum JS4 (KM513609) | F8114, F8118, F8129, F8134, F8140, F8152, F8162, F8167, F8188, F8190, F8198, F8231 | 100.00 | 55 | 16.57 |
| 29 | Colletotrichum truncatum SL3-CJL17 (KP900265) | F8119, F8141, F8171, F8174, F8175, F8177, F8187, F8202, F8211, F8212, F8221, F8244 | 99.81 | 109 | 32.83 |
| 30 | Colletotrichum fructicola JXNC-7 (MK041519) | F8136, F8155 | 99.81 | 2 | 0.60 |
| 31 | Colletotrichum gloeosporioides HBxn-3 (HQ645077) | F8203 | 99.26 | 1 | 0.30 |
| 32 | Colletotrichum higginsianum XN4-5 (JF830783) | F8239, F8240 | 99.63 | 2 | 0.60 |
| 33 | Trichoderma cf. harzianum PDAN04-1 (MF108878) | F8158 | 99.66 | 2 | 0.60 |
| 34 | Trichoderma sp. SDAS203699 (MK870422) | F8166, F8182, F8197, F8220, F8223, F8226 | 98.99 | 6 | 1.81 |
| 35 | Hypoxylon fragiforme RY-3 (MK429859) | F8218 | 98.97 | 1 | 0.30 |
| 36 | Ceriporia alachuana CEAL72 (MF358878) | F8112 | 99.36 | 1 | 0.30 |
| 37 | Mucor fragilis BC3 (MK910073) | F8199 | 99.50 | 1 | 0.30 |
| 38 | Muyocopron lithocarpi MFLUCC:17-1500 (MT137781) | F8248 | 99.82 | 1 | 0.30 |
| 39 | Septoria arundinacea LHH10-8 (JX077003) | F8125, F8185, F8219, F8246 | 100.00 | 4 | 1.20 |
| 40 | Fusarium sambucinum CBS 146.95 (KM231813) | F8126, F8132 | 98.79 | 2 | 0.60 |
| 41 | Fusarium verticillioides G4 (MK264336) | F8200 | 100.00 | 1 | 0.30 |
| 42 | Calonectria eucalypti (KM357290) | F8168 | 99.03 | 1 | 0.30 |
| 43 | Ilyonectria robusta (MN121555) | F8145 | 99.40 | 1 | 0.30 |
| 44 | Dactylonectria alcacerensis JZB3310013 (MN944923) | F8217 | 99.42 | 1 | 0.30 |
| 45 | Nectria pseudotrichia (MN121548) | F8131 | 98.58 | 2 | 0.60 |
| 46 | Neonectria sp. nc_gw_9967b (KF428559) | F8153 | 98.82 | 1 | 0.30 |
| 47 | Plectosphaerella sp. GZUIFR-QL9.9.1 (MK880441) | F8247 | 98.86 | 1 | 0.30 |
| 48 | Alternaria alternata Acf-4 (MK795217) | F8214, F8215 | 99.63 | 2 | 0.60 |
| 49 | Talaromyces funiculosus S01324 (MG744693) | F8138, F8149 | 99.46 | 2 | 0.60 |
| 50 | Talaromyces pinophilus KR9 (MF153381) | F8230 | 99.83 | 1 | 0.30 |
| 51 | Talaromyces purpureogenus NFML_CH66 (KM458841) | F8222 | 98.73 | 1 | 0.30 |
| 52 | Nemania sp. 1 XS-2016 VD026 (KT588467) | F8183, F8229, F8113 | 99.26 | 3 | 0.90 |
| 53 | Biscogniauxia sp. strain LPS-70 (MF379340) | F8130, F8147 | 99.63 | 2 | 0.60 |
| 54 | Leptospora rubella strain ZLVG 319 (HE774478) | F8127 | 96.38 | 1 | 0.30 |
| 55 | Ascomycota sp. isolate FL15 (MK370697 | F8146 | 90.00 | 1 | 0.30 |
| 56 | Sordariomycetes sp. 329 isolate FL0587 (JQ760299) | F8184, F8209, F8233, F8241, F8245 | 86.76 | 5 | 1.51 |
| Total | 130 | 130 | 332 | 100 | |
| Species Richness (S) | 56 | ||||
| Shannon-Wiener index (H′) | 2.7076 | ||||
| No. | Phylum | Class | Order | Family | Genus | Number (N) |
|---|---|---|---|---|---|---|
| 1 | Ascomycota | Sordariomycetes | Xylariales | Apiosporaceae | Arthrinium | 1 |
| 2 | Hypocreales | Xylariaceae | Nemania | 3 | ||
| 3 | Biscogniauxia | 4 | ||||
| 4 | Hypoxylaceae | Hypoxylon | 1 | |||
| 5 | Bionectriaceae | Clonostachys | 1 | |||
| 6 | Nectriaceae | Fusarium | 3 | |||
| 7 | Calonectria | 1 | ||||
| 8 | Ilyonectria | 1 | ||||
| 9 | Dactylonectria | 1 | ||||
| 10 | Nectria | 1 | ||||
| 11 | Neonectria | 1 | ||||
| 12 | Hypocreaceae | Trichoderma | 8 | |||
| 13 | Sordariales | Chaetomiaceae | Podospora | 4 | ||
| 14 | Diaporthales | Diaporthaceae | Diaporthe | 31 | ||
| 15 | Glomerellales | Glomerellaceae | Colletotrichum | 201 | ||
| 16 | Plectosphaerellaceae | Plectosphaerella | 1 | |||
| 17 | Eurotiomycetes | Eurotiales | Aspergillaceae | Aspergillus | 39 | |
| 18 | Trichocomaceae | Talaromyces | 4 | |||
| 19 | Dothideomycetes | Capnodiales | Cladosporiaceae | Cladosporium | 1 | |
| 20 | Leptospora | 1 | ||||
| 21 | Pleosporales | Mycosphaerellaceae | Septoria | 4 | ||
| 22 | Didymellaceae | Stagonosporopsis | 1 | |||
| 23 | Didymella | 1 | ||||
| 24 | Dothiorella | 1 | ||||
| 25 | Pleosporaceae | Alternaria | 2 | |||
| 26 | Didymosphaeriaceae | Paraconiothyrium | 1 | |||
| 27 | Muyocopronales | Muyocopronaceae | Muyocopron | 1 | ||
| 28 | Basidiomycota | Agaricomycetes | Cantharellales | Ceratobasidiaceae | Thanatephorus | 6 |
| 29 | Polyporales | Irpicaceae | Ceriporia | 1 | ||
| 30 | Mucoromycota | Mucoromycetes | Mucorales | Mucoraceae | Mucor | 1 |
| Total | 3 | 5 | 12 | 21 | 30 | 325 |
| Isolates No. | Taxa (Accession Number) | Inhibition Zone in Diameter on Petri Plate (mm) | ||||||
|---|---|---|---|---|---|---|---|---|
| B. cereus | E. coli | B. subtilis | S. aureus | P. aeruginosa | R. solanacearum | C. albicans | ||
| F8107 | Diaporthe eres (MN429162) | 12.02 ± 0.11 | 10.44 ± 0.04 | 10.93 ± 0.05 | 13.27 ± 0.16 | - | 10.10 ± 0.07 | - |
| F8121 | Aspergillus terreus (MN429176) | 17.14 ± 0.13 | 10.08 ± 0.14 | 20.65 ± 0.40 | 16.12 ± 0.08 | - | - | 9.09 ± 0.04 |
| F8127 | Leptospora rubella (MN429181) | 11.34 ± 0.12 | - | 14.26 ± 0.09 | 12.58 ± 0.15 | - | 8.96 ± 0.08 | - |
| F8157 | Thanatephorus cucumeris (MN429209) | - | - | 12.14 ± 0.14 | - | 10.32 ± 0.20 | 8.27 ± 0.18 | 8.94 ± 0.05 |
| F8158 | Trichoderma cf. harzianum (MN429210) | 19.15 ± 0.13 | 10.81 ± 0.28 | 19.12 ± 0.15 | 19.36 ± 0.14 | 10.21 ± 0.12 | 12.02 ± 0.03 | 10.81 ± 0.16 |
| F8159 | Aspergillus terreus (MN429211) | 15.33 ± 0.13 | 10.76 ± 0.25 | 21.15 ± 0.27 | 17.37 ± 0.11 | - | 16.48 ± 0.13 | 9.75 ± 0.17 |
| F8160 | Aspergillus oryzae (MN429212) | 11.21 ± 0.14 | - | 12.13 ± 0.10 | 12.12 ± 0.08 | - | 11.02 ± 0.03 | - |
| F8172 | Aspergillus terreus (MN429223) | 17.31 ± 0.19 | 10.84 ± 0.03 | 20.82 ± 0.17 | 16.39 ± 0.09 | - | 12.46 ± 0.10 | 10.21 ± 0.12 |
| F8189 | Colletotrichum acutatum (MN429239) | 12.30 ± 0.17 | - | 16.04 ± 0.13 | 13.17 ± 0.18 | - | 11.05 ± 0.09 | 9.14 ± 0.07 |
| F8191 | Aspergillus terreus (MN429241) | 20.56 ± 0.30 | - | 20.62 ± 0.41 | 15.38 ± 0.08 | - | 11.28 ± 0.15 | 9.34 ± 0.12 |
| F8204 | Aspergillus flavipes (MN429254) | 12.29 ± 0.18 | - | 10.27 ± 0.18 | - | - | 8.33 ± 0.13 | - |
| F8219 | Septoria arundinacea (MN429268) | 10.21 ± 0.13 | - | 10.98 ± 0.08 | 11.29 ± 0.15 | - | 10.85 ± 0.12 | |
| F8225 | Aspergillus flavipes (MN429274) | 11.13 ± 0.12 | - | 15.31 ± 0.24 | 16.23 ± 0.12 | - | 13.94 ± 0.08 | - |
| Positive control-1 | Ampicillin sodium | 20.48 ± 0.53 | 15.46 ± 0.22 | 23.34 ± 0.38 | 21.49 ± 0.27 | 17.34 ± 0.26 | 19.55 ± 0.21 | - |
| Positive control-2 | Actidione | - | - | - | - | - | - | 21.15 ± 0.84 |
| Negative control | Methanol | - | - | - | - | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, C.; Ma, S.; Shi, X.; Xue, W.; Liu, C.; Ding, H. Diversity and Antimicrobial Activity of Endophytic Fungi Isolated from Chloranthus japonicus Sieb in Qinling Mountains, China. Int. J. Mol. Sci. 2020, 21, 5958. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21175958
An C, Ma S, Shi X, Xue W, Liu C, Ding H. Diversity and Antimicrobial Activity of Endophytic Fungi Isolated from Chloranthus japonicus Sieb in Qinling Mountains, China. International Journal of Molecular Sciences. 2020; 21(17):5958. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21175958
Chicago/Turabian StyleAn, Chao, Saijian Ma, Xinwei Shi, Wenjiao Xue, Chen Liu, and Hao Ding. 2020. "Diversity and Antimicrobial Activity of Endophytic Fungi Isolated from Chloranthus japonicus Sieb in Qinling Mountains, China" International Journal of Molecular Sciences 21, no. 17: 5958. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21175958