Inflammatory Cytokine-Producing Cells and Inflammation Markers in the Synovium of Osteoarthritis Patients Evidenced in Human Herpesvirus 7 Infection

 , and
, and 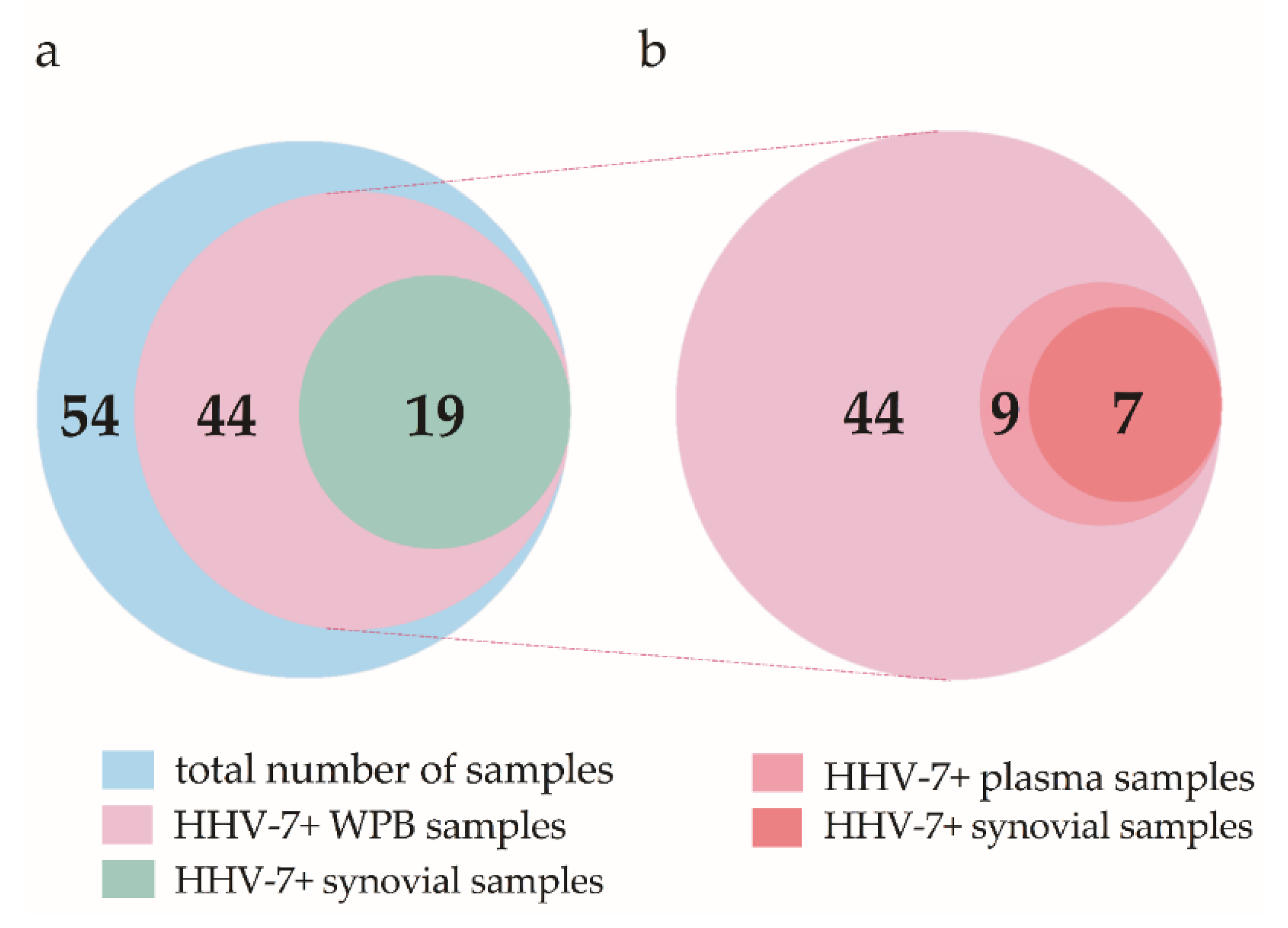{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Nested Polymerase Chain Reaction
2.2. Plasma Levels of TNF and IL-6
2.3. Assessment of Synovitis Applying the Krenn Scoring System
2.4. Histopathology and Immunohistochemical Detection of Antigens within the Synovial Membrane
3. Discussion
4. Materials and Methods
4.1. Patients’ Characteristics
4.2. Blood Sample Collection and Detection of TNF and IL-6 Levels
4.3. Nested Polymerase Chain Reaction
4.4. Light Microscopy and Immunohistochemistry
4.5. Scoring of Synovitis by Krenn
4.6. Statistical Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fusco, M.; Skaper, S.D.; Coaccioli, S.; Varrassi, G.; Paladini, A. Degenerative Joint Diseases and Neuroinflammation. Pain Pract. 2017, 17, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Bortoluzzi, A.; Furini, F.; Generali, E.; Silvagni, E.; Luciano, N.; Scirè, C.A. One year in review 2018: Novelties in the treatment of rheumatoid arthritis. Clin. Exp. Rheumatol. 2018, 36, 347–361. [Google Scholar] [PubMed]
- Fu, K.; Robbins, S.; McDougall, J.J. Osteoarthritis: The genesis of pain. Rheumatology 2017, 57, iv43–iv50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruyère, O.; Cooper, C.; Pelletier, J.-P.; Branco, J.C.; Brandi, M.L.; Guillemin, F.; Hochberg, M.C.; Kanis, J.; Kvien, T.K.; Martel-Pelletier, J.; et al. An algorithm recommendation for the management of knee osteoarthritis in Europe and internationally: A report from a task force of the European Society for Clinical and Economic Aspects of Osteoporosis and Osteoarthritis (ESCEO). Semin. Arthritis Rheum. 2014, 44, 253–263. [Google Scholar] [CrossRef]
- Goldring, M.B.; Otero, M.; Plumb, D.A.; Dragomir, C.; Favero, M.; Hachem, K.E.; Hashimoto, K.; Roach, H.; Olivotto, E.; Borzì, R.; et al. Roles of inflammatory and anabolic cytokines in cartilage metabolism: Signals and multiple effectors converge upon MMP-13 regulation in osteoarthritis. Eur. Cells Mater. 2011, 21, 202–220. [Google Scholar] [CrossRef]
- Rahmati, M.; Mobasheri, A.; Mozafari, M. Inflammatory mediators in osteoarthritis: A critical review of the state-of-the-art, current prospects, and future challenges. Bone 2016, 85, 81–90. [Google Scholar] [CrossRef]
- Li, Y.-S.; Luo, W.; Zhu, S.; Lei, G. T Cells in Osteoarthritis: Alterations and Beyond. Front. Immunol. 2017, 8, 356. [Google Scholar] [CrossRef] [Green Version]
- Griffin, T.M.; Scanzello, C.R. Innate inflammation and synovial macrophages in osteoarthritis pathophysiology. Clin. Exp. Rheumatol. 2019, 37, 57–63. [Google Scholar]
- Berenbaum, F. Osteoarthritis as an inflammatory disease (osteoarthritis is not osteoarthrosis!). Osteoarthr. Cartil. 2013, 21, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Iwanaga, T.; Shikichi, M.; Kitamura, H.; Yanase, H.; Nozawa-Inoue, K. Morphology and Functional Roles of Synoviocytes in the Joint. Arch. Histol. Cytol. 2000, 63, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Shikichi, M.; Kitamura, H.P.; Yanase, H.; Konno, A.; Takahashi-Iwanaga, H.; Iwanaga, T. Three-dimensional Ultrastructure of Synoviocytes in the Horse Joint as Revealed by the Scanning Electron Microscope. Arch. Histol. Cytol. 1999, 62, 219–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, M.J.; Herndler-Brandstetter, D.; Tariq, M.A.; Nicholson, T.A.; Philp, A.M.; Smith, H.L.; Davis, E.T.; Jones, S.W.; Lord, J.M. IL-6 secretion in osteoarthritis patients is mediated by chondrocyte-synovial fibroblast cross-talk and is enhanced by obesity. Sci. Rep. 2017, 7, 3451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkelaar, M.H.M.; Korthagen, N.M.; Jansen, G.; van Spil, W.E. Synovial Macrophages: Potential Key Modulators of Cartilage Damage, Osteophyte Formation and Pain in Knee Osteoarthritis. J. Rheum. Dis. Treat. 2018, 4, 059. [Google Scholar]
- Moqbel, S.A.A.; He, Y.; Xu, L.; Ma, C.; Ran, J.; Xu, K.; Wu, L. Rat Chondrocyte Inflammation and Osteoarthritis Are Ameliorated by Madecassoside. Oxidative Med. Cell Longev. 2020, 2020, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutton, S.; Clutterbuck, A.; Harris, P.; Gent, T.C.; Freeman, S.L.; Foster, N.; Barrett-Jolley, R.; Mobasheri, A. The contribution of the synovium, synovial derived inflammatory cytokines and neuropeptides to the pathogenesis of osteoarthritis. Vet. J. 2009, 179, 10–24. [Google Scholar] [CrossRef]
- Zakrzewska, K.; Azzi, A.; de Biasi, E.; Radossi, P.; de Santis, R.; Davoli, P.; Tagariello, G. Persistence of parvovirus B19 DNA in synovium of patients with haemophilic arthritis. J. Med. Virol. 2001, 65, 402–407. [Google Scholar] [CrossRef]
- Mehraein, Y.; Lennerz, C.; Ehlhardt, S.; Venzke, T.; Ojak, A.; Remberger, K.; Zang, K.D. Detection of Parvovirus B19 Capsid Proteins in Lymphocytic Cells in Synovial Tissue of Autoimmune Chronic Arthritis. Mod. Pathol. 2003, 16, 811–817. [Google Scholar] [CrossRef] [Green Version]
- Mehraein, Y.; Lennerz, C.; Ehlhardt, S.; Zang, K.D.; Madry, H. Replicative multivirus infection with cytomegalovirus, herpes simplex virus 1, and parvovirus B19, and latent Epstein–Barr virus infection in the synovial tissue of a psoriatic arthritis patient. J. Clin. Virol. 2004, 31, 25–31. [Google Scholar] [CrossRef]
- Naciute, M.; Mieliauskaite, D.; Rugiene, R.; Nikitenkiene, R.; Jancoriene, L.; Mauricas, M.; Nora-Krūkle, Z.; Murovska, M.; Girkontaite, I. Frequency and significance of parvovirus B19 infection in patients with rheumatoid arthritis. J. Gen. Virol. 2016, 97, 3302–3312. [Google Scholar] [CrossRef]
- Marks, M.; Marks, J.L. Viral arthritis. Clin. Med. 2016, 16, 129. [Google Scholar] [CrossRef]
- Agut, H.; Bonnafous, P.; Gautheret-Dejean, A. Human Herpesviruses 6A, 6B, and 7. Microbiol. Spectr. 2016, 4, 157–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agut, H.; Bonnafous, P.; Gautheret-Dejean, A. Update on infections with human herpesviruses 6A, 6B, and 7. Médecine Mal. Infect. 2017, 47, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wołącewicz, M.; Becht, R.; Grywalska, E.; Niedźwiedzka-Rystwej, P. Herpesviruses in Head and Neck Cancers. Viruses 2020, 12, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denner, J.; Bigley, T.M.; Phan, T.L.; Zimmermann, C.; Zhou, X.; Kaufer, B.B. Comparative Analysis of Roseoloviruses in Humans, Pigs, Mice, and Other Species. Viruses 2019, 11, 1108. [Google Scholar] [CrossRef] [Green Version]
- Drago, F.; Malaguti, F.; Ranieri, E.; Losi, E.; Rebora, A. Human herpes virus-like particles in pityriasis rosea lesions: An electron microscopy study. J. Cutan. Pathol. 2002, 29, 359–361. [Google Scholar] [CrossRef]
- Watanabe, T.; Kawamura, T.; Aquilino, E.A.; Blauvelt, A.; Jacob, S.E.; Orenstein, J.M.; Black, J.B. Pityriasis Rosea is Associated with Systemic Active Infection with Both Human Herpesvirus-7 and Human Herpesvirus-6. J. Investig. Dermatol. 2002, 119, 793–797. [Google Scholar] [CrossRef] [Green Version]
- Broccolo, F.; Drago, F.; Careddu, A.M.; Foglieni, C.; Turbino, L.; Cocuzza, C.E.; Gelmetti, C.; Lusso, P.; Rebora, A.; Malnati, M.S. Additional Evidence that Pityriasis Rosea Is Associated with Reactivation of Human Herpesvirus-6 and -7. J. Investig. Dermatol. 2005, 124, 1234–1240. [Google Scholar] [CrossRef] [Green Version]
- Drago, F.; Ciccarese, G.; Parodi, A. HHV-6 reactivation as a cause of fever in autologous hematopoietic stem cell transplant recipients: A reply. J. Infect. 2018, 76, 101–102. [Google Scholar] [CrossRef]
- Nahidi, Y.; Meibodi, N.T.; Ghazvini, K.; Esmaily, H.; Esmaeelzadeh, M. Association of classic lichen planus with human herpesvirus-7 infection. Int. J. Dermatol. 2016, 56, 49–53. [Google Scholar] [CrossRef]
- Skuja, S.; Zieda, A.; Ravina, K.; Chapenko, S.; Roga, S.; Teteris, O.; Groma, V.; Murovska, M. Structural and Ultrastructural Alterations in Human Olfactory Pathways and Possible Associations with Herpesvirus 6 Infection. PLoS ONE 2017, 12, e0170071. [Google Scholar] [CrossRef] [Green Version]
- Kadiša, A.; Nora-Krūkle, Z.; Kozireva, S.; Svirskis, S.; Studers, P.; Groma, V.; Lejnieks, A.; Murovska, M. Effect of Human Herpesviruses 6 and 7 Infection on the Clinical Course of Rheumatoid Arthritis/Cilvēka Herpesvīrusa 6 un 7 Infekcijas Ietekme uz Reimatoīdā Artrīta Klīnisko Gaitu. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2016, 70, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Kondo, K.; Yamanishi, K. HHV-6A, 6B, and 7: Molecular basis of latency and reactivation. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Cambridge University Press (CUP): Cambridge, UK, 2010; pp. 843–849. [Google Scholar]
- Burgos, R.; Ordoñez, G.; Vázquez-Mellado, J.; Pineda, B.; Sotelo, J. Occasional presence of herpes viruses in synovial fluid and blood from patients with rheumatoid arthritis and axial spondyloarthritis. Clin. Rheumatol. 2015, 34, 1681–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, J.R.; Xie, F.; Yun, H.; Bernatsky, S.; Winthrop, K.L. Real-world comparative risks of herpes virus infections in tofacitinib and biologic-treated patients with rheumatoid arthritis. Ann. Rheum. Dis. 2016, 75, 1843–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siebuhr, A.S.; Bay-Jensen, A.; Jordan, J.; Kjelgaard-Petersen, C.; Christiansen, C.; Abramson, S.; Attur, M.; Berenbaum, F.; Kraus, V.; Karsdal, M.; et al. Inflammation (or synovitis)-driven osteoarthritis: An opportunity for personalizing prognosis and treatment? Scand. J. Rheumatol. 2015, 45, 1–12. [Google Scholar] [CrossRef]
- Hussein, M.R.; Fathi, N.A.; El-Din, A.M.E.; Hassan, H.I.; Abdullah, F.; Al-Hakeem, E.; Backer, E.A. Alterations of the CD4+, CD8+ T Cell Subsets, Interleukins-1β, IL-10, IL-17, Tumor Necrosis Factor-α and Soluble Intercellular Adhesion Molecule-1 in Rheumatoid Arthritis and Osteoarthritis: Preliminary Observations. Pathol. Oncol. Res. 2008, 14, 321–328. [Google Scholar] [CrossRef]
- Krenn, V.; Morawietz, L.; Haupl, T.; Neidel, J.; Petersen, I.; König, A. Grading of Chronic Synovitis—A Histopathological Grading System for Molecular and Diagnostic Pathology. Pathol. Res. Pract. 2002, 198, 317–325. [Google Scholar] [CrossRef]
- de Lange-Brokaar, B.J.E.; Ioan-Facsinay, A.; van Osch, G.; Zuurmond, A.-M.; Schoones, J.W.; Toes, R.E.; Huizinga, T.W.; Kloppenburg, M. Synovial inflammation, immune cells and their cytokines in osteoarthritis: A review. Osteoarthr. Cartil. 2012, 20, 1484–1499. [Google Scholar] [CrossRef] [Green Version]
- Riis, R.G.C.; Gudbergsen, H.; Simonsen, O.; Henriksen, M.; Al-Mashkur, N.M.; Eld, M.; Petersen, K.; Kubassova, O.; Bay-Jensen, A.-C.; Damm, J.; et al. The association between histological, macroscopic and magnetic resonance imaging assessed synovitis in end-stage knee osteoarthritis: A cross-sectional study. Osteoarthr. Cartil. 2017, 25, 272–280. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y. Recent topics related to human herpesvirus 6 cell tropism. Cell Microbiol. 2009, 11, 1001–1006. [Google Scholar] [CrossRef]
- Tang, H.; Serada, S.; Kawabata, A.; Ota, M.; Hayashi, E.; Naka, T.; Yamanishi, K.; Mori, Y. CD134 is a cellular receptor specific for human herpesvirus-6B entry. Proc. Natl. Acad. Sci. USA 2013, 110, 9096–9099. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.; Wang, J.; Mahmoud, N.F.; Mori, Y. Detailed Study of the Interaction between Human Herpesvirus 6B Glycoprotein Complex and Its Cellular Receptor, Human CD134. J. Virol. 2014, 88, 10875–10882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.; Mori, Y. Determinants of Human CD134 Essential for Entry of Human Herpesvirus 6B. J. Virol. 2015, 89, 10125–10129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frenkel, N.; Schirmer, E.C.; Wyatt, L.S.; Katsafanas, G.; Roffman, E.; Danovich, R.M.; June, C.H. Isolation of a new herpesvirus from human CD4+ T cells. Proc. Natl. Acad. Sci. USA 1990, 87, 748–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caselli, E.; di Luca, D. Molecular biology and clinical associations of Roseoloviruses human herpesvirus 6 and human herpesvirus 7. Microbiol. Q. J. Microbiol. Sci. 2007, 30, 173–187. [Google Scholar]
- Pourgheysari, B.; Khan, N.; Best, D.; Bruton, R.; Nayak, L.; Moss, P. The Cytomegalovirus-Specific CD4+ T-Cell Response Expands with Age and Markedly Alters the CD4+ T-Cell Repertoire. J. Virol. 2007, 81, 7759–7765. [Google Scholar] [CrossRef] [Green Version]
- Pawłowska, J.; Mikosik, A.; Soroczynska-Cybula, M.; Jozwik, A.; Łuczkiewicz, P.; Mazurkiewicz, S.; Lorczyński, A.; Witkowski, J.M.; Bryl, E. Different distribution of CD4 and CD8 T cells in synovial membrane and peripheral blood of rheumatoid arthritis and osteoarthritis patients. Folia Histochem. Cytobiol. 2010, 47, 627–632. [Google Scholar] [CrossRef]
- Yamada, H.; Nakashima, Y.; Okazaki, K.; Mawatari, T.; Fukushi, J.-I.; Oyamada, A.; Fujimura, K.; Iwamoto, Y.; Yoshikai, Y. Preferential Accumulation of Activated Th1 Cells Not Only in Rheumatoid Arthritis but Also in Osteoarthritis Joints. J. Rheumatol. 2011, 38, 1569–1575. [Google Scholar] [CrossRef]
- Moradi, B.; Schnatzer, P.; Hagmann, S.; Rosshirt, N.; Gotterbarm, T.; Kretzer, J.; Thomsen, M.N.; Lorenz, H.-M.; Zeifang, F.; Tretter, T. CD4+CD25+/highCD127low/-regulatory T cells are enriched in rheumatoid arthritis and osteoarthritis joints—Analysis of frequency and phenotype in synovial membrane, synovial fluid and peripheral blood. Arthritis Res. Ther. 2014, 16, R97. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Zhang, X.; Jiang, Y.; Liu, X.; Huang, L.; Wei, Q.; Huang, Y.; Wu, W.; Gu, J. Alterations in peripheral T cell and B cell subsets in patients with osteoarthritis. Clin. Rheumatol. 2019, 39, 523–532. [Google Scholar] [CrossRef]
- Shen, P.-C.; Wu, C.-L.; Jou, I.-M.; Lee, C.-H.; Juan, H.-Y.; Lee, P.-J.; Chen, S.-H.; Hsieh, J.-L. T helper cells promote disease progression of osteoarthritis by inducing macrophage inflammatory protein-1γ. Osteoarthr. Cartil. 2011, 19, 728–736. [Google Scholar] [CrossRef] [Green Version]
- Staheli, J.P.; Dyen, M.R.; Deutsch, G.; Basom, R.S.; FitzGibbon, M.P.; Lewis, P.; Barcy, S. Complete Unique Genome Sequence, Expression Profile, and Salivary Gland Tissue Tropism of the Herpesvirus 7 Homolog in Pigtailed Macaques. J. Virol. 2016, 90, 6657–6674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowd, J.B.; Bosch, J.A.; Steptoe, A.; Jayabalasingham, B.; Lin, J.; Yolken, R.; Aiello, A.E. Persistent Herpesvirus Infections and Telomere Attrition Over 3 Years in the Whitehall II Cohort. J. Infect. Dis. 2017, 216, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Weidner, M.; Kruminis-Kaszkiel, E.; Savanagouder, M. Herpesviral Latency—Common Themes. Pathogens 2020, 9, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Ponce, Y.; Varela-Fascinetto, G.; Romo-Vázquez, J.C.; Martínez, B.L.; Sánchez-Huerta, J.L.; Parra-Ortega, I.; Fuentes-Pananá, E.M.; Sánchez, A.M. Simultaneous Detection of Beta and Gamma Human Herpesviruses by Multiplex qPCR Reveals Simple Infection and Coinfection Episodes Increasing Risk for Graft Rejection in Solid Organ Transplantation. Viruses 2018, 10, 730. [Google Scholar] [CrossRef] [Green Version]
- Kakurina, N.; Kadisa, A.; Lejnieks, A.; Mikazane, H.; Kozireva, S.; Murovska, M. Use of exploratory factor analysis to ascertain the correlation between the activities of rheumatoid arthritis and infection by human parvovirus B19. Medicina 2015, 51, 18–24. [Google Scholar] [CrossRef]
- Kadiša, A.; Nora-Krūkle, Z.; Švirskis, S.; Studers, P.; Girkontaite, I.; Lejnieks, A.; Murovska, M. Cytokines and MMP-9 Levels in Rheumatoid Arthritis and Osteoarthritis Patients with Persistent Parvovirus B19, HHV-6 and HHV-7 Infection. Proc. Latv. Acad. Sci. 2019, 73, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Humby, M.S.; O’Connor, C.M. Human Cytomegalovirus US28 Is Important for Latent Infection of Hematopoietic Progenitor Cells. J. Virol. 2015, 90, 2959–2970. [Google Scholar] [CrossRef] [Green Version]
- Poole, E.; Sinclair, J. Sleepless latency of human cytomegalovirus. Med. Microbiol. Immunol. 2015, 204, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Wills, M.; Poole, E.; Lau, B.; Krishna, B.; Sinclair, J.H. The immunology of human cytomegalovirus latency: Could latent infection be cleared by novel immunotherapeutic strategies? Cell Mol. Immunol. 2014, 12, 128–138. [Google Scholar] [CrossRef]
- Prusty, B.K.; Gulve, N.; Rasa, S.; Murovska, M.; Hernandez, P.C.; Ablashi, D.V. Possible chromosomal and germline integration of human herpesvirus 7. J. Gen. Virol. 2017, 98, 266–274. [Google Scholar] [CrossRef]
- O’Donovan, A.; Pantell, M.S.; Puterman, E.; Dhabhar, F.S.; Blackburn, E.H.; Yaffe, K.; Cawthon, R.M.; Opresko, P.; Hsueh, W.-C.; Satterfield, S.; et al. Cumulative Inflammatory Load Is Associated with Short Leukocyte Telomere Length in the Health, Aging and Body Composition Study. PLoS ONE 2011, 6, e19687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.; de Vivo, I.; Lin, X.; Fang, S.C.; Christiani, D.C. The Relationship between Inflammatory Biomarkers and Telomere Length in an Occupational Prospective Cohort Study. PLoS ONE 2014, 9, e87348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Bai, L. Recent advances in the understanding of molecular mechanisms of cartilage degeneration, synovitis and subchondral bone changes in osteoarthritis. Connect. Tissue Res. 2016, 57, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Haynes, M.K.; Hume, E.L.; Smith, J.B. Phenotypic Characterization of Inflammatory Cells from Osteoarthritic Synovium and Synovial Fluids. Clin. Immunol. 2002, 105, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S.S.; Klotz, U.; Alscher, D.M.; Mais, A.; Lauer, G.; Schweer, H.; Seyberth, H.W.; Fritz, P.; Bierbach, U.; Alscher, M.D. Osteoarthritis of the knee–clinical assessments and inflammatory markers. Osteoarthr. Cartil. 2004, 12, 469–475. [Google Scholar] [CrossRef] [Green Version]
- Benito, M.J.; Veale, D.J.; Fitzgerald, O.; Berg, W.B.V.D.; Bresnihan, B. Synovial tissue inflammation in early and late osteoarthritis. Ann. Rheum. Dis. 2005, 64, 1263–1267. [Google Scholar] [CrossRef] [Green Version]
- May, N.A.; Glosson, N.L.; Hudson, A.W. Human Herpesvirus 7 U21 Downregulates Classical and Nonclassical Class I Major Histocompatibility Complex Molecules from the Cell Surface. J. Virol. 2010, 84, 3738–3751. [Google Scholar] [CrossRef] [Green Version]
- Altman, R.; Alarcon, G.; Appelrouth, D.; Bloch, D.; Borenstein, D.; Brandt, K.; Brown, C.; Cooke, T.D.; Daniel, W.; Feldman, D.; et al. The American College of Rheumatology criteria for the classification and reporting of osteoarthritis of the hip. Arthritis Rheum. 1991, 34, 505–514. [Google Scholar] [CrossRef]
- Altman, R.; Asch, E.; Bloch, D.; Bole, G.; Borenstein, D.; Brandt, K.; Christy, W.; Cooke, T.D.; Greenwald, R.; Hochberg, M.; et al. Development of criteria for the classification and reporting of osteoarthritis: Classification of osteoarthritis of the knee. Arthritis Rheum. 1986, 29, 1039–1049. [Google Scholar] [CrossRef]
- Berneman, Z.N.; Ablashi, D.V.; Li, G.; Eger-Fletcher, M.; Reitz, M.S.; Hung, C.L.; Brus, I.; Komaroff, A.L.; Gallo, R.C. Human herpesvirus 7 is a T-lymphotropic virus and is related to, but significantly different from, human herpesvirus 6 and human cytomegalovirus. Proc. Natl. Acad. Sci. USA 1992, 89, 10552–10556. [Google Scholar] [CrossRef] [Green Version]
- Kozireva, S.; Užameckis, D.; Bariševs, M.; Murovska, M. Sensitivity and Reproducibility of Polymerase Chain Reaction Assays for Detection of Human Herpesviruses 6 and 7. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2009, 63, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Kempf, W.; Adams, V.; Mirandola, P.; Menotti, L.; di Luca, D.; Wey, N.; Müller, B.; Campadelli-Fiume, G. Persistence of human herpesvirus 7 in normal tissues detected by expression of a structural antigen. J. Infect. Dis. 1998, 178, 841–845. [Google Scholar] [CrossRef] [PubMed]
- Latchney, L.R.; Fallon, M.A.; Culp, D.J.; Gelbard, H.A.; Dewhurst, S. Immunohistochemical Assessment of Fractalkine, Inflammatory Cells, and Human Herpesvirus 7 in Human Salivary Glands. J. Histochem. Cytochem. 2004, 52, 671–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Shi, F.; Li, Y.; Yu, X.; Peng, S.; Li, W.; Luo, X.; Cao, Y. Post-translational modifications as key regulators of TNF-induced necroptosis. Cell Death Dis. 2016, 7, e2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krenn, V.; Morawietz, L.; Burmester, G.-R.; Kinne, R.W.; Müller, B.; Häupl, T.; Mueller-Ladner, U. Synovitis score: Discrimination between chronic low-grade and high-grade synovitis. Histopathology 2006, 49, 358–364. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Groma, V.; Tarasovs, M.; Skuja, S.; Semenistaja, S.; Nora-Krukle, Z.; Svirskis, S.; Murovska, M. Inflammatory Cytokine-Producing Cells and Inflammation Markers in the Synovium of Osteoarthritis Patients Evidenced in Human Herpesvirus 7 Infection. Int. J. Mol. Sci. 2020, 21, 6004. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176004
Groma V, Tarasovs M, Skuja S, Semenistaja S, Nora-Krukle Z, Svirskis S, Murovska M. Inflammatory Cytokine-Producing Cells and Inflammation Markers in the Synovium of Osteoarthritis Patients Evidenced in Human Herpesvirus 7 Infection. International Journal of Molecular Sciences. 2020; 21(17):6004. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176004
Chicago/Turabian StyleGroma, Valerija, Mihails Tarasovs, Sandra Skuja, Sofija Semenistaja, Zaiga Nora-Krukle, Simons Svirskis, and Modra Murovska. 2020. "Inflammatory Cytokine-Producing Cells and Inflammation Markers in the Synovium of Osteoarthritis Patients Evidenced in Human Herpesvirus 7 Infection" International Journal of Molecular Sciences 21, no. 17: 6004. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176004